










 Serviços customizados
Serviços customizados Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Jatropha curcas L., conocida como piñón, es una planta oleaginosa originaria de América tropical (Tsuchimoto, 2017), que pertenece a la familia Euphorbiaceae, con aproximadamente 188 especies, de amplia distribución en Centroamérica, el Caribe, Sudamérica, Asia y África. Según Laviola et al. (2017), lo más probable es que haya sido distribuida desde el Caribe por los navegantes portugueses a países de África a través de Cabo Verde y Guinea Bissau, así como a naciones del sudeste de Asia, como Indonesia, Malasia y Filipinas.
En la actualidad, se cultiva J. curcas para la producción de biocombustible (Borah et al., 2018) en el centro y sudeste de la India. También en Asia, África y América se realizan investigaciones acerca de sus potencialidades para este fin (Savaliya et al., 2015).
J. curcas se considera una planta multipropósito (Zavala, 2016), debido a la variedad de usos que tiene. Su cobertura vegetal se aprovecha, en la mayor parte de la India, para proteger los cultivos del daño causado por los animales, ya que el vacuno y caprino no la consume. En este sentido, el extracto acuoso del aceite de sus semillas contiene toxinas biodegradables, que se pueden aplicar como insecticida natural en diversos cultivos (Laviola et al., 2017).
El uso comestible de J. curcas solo se ha documentado en México (Pecina-Quintero et al., 2014), en los estados de Quintana Roo y Veracruz. En este último, en particular en Totonacapan, se usa tradicionalmente como alimento (Valdés-Rodríguez et al., 2013). En esta región, sus semillas se emplean como botana, salsas, tamales y en platillos tradicionales (Martínez-Herrera et al., 2012). Estos materiales se consideran no tóxicos, por carecer de ésteres de forbol. Para el caso de los territorios del sur del estado, su uso es muy limitado, debido a que la mayoría de las plantas son tóxicas (Sukla et al., 2015).
En los últimos años, se ha prestado gran atención a las características fisiológicas, agronómicas, agroecológicas y productivas de J. curcas (Niestche et al., 2015). Esta caracterización considera descriptores cuantitativos y cualitativos (Martín y Montes, 2015). Algunos trabajos en varias poblaciones de esta especie han informado bajo nivel de variabilidad genética (Santos et al., 2016).
A partir de lo anterior, el objetivo de este estudio fue caracterizar la variabilidad morfoagronómica de accesiones no tóxicas de J. curcas, propagadas por semillas y por estacas o varetas; establecidas hace ocho años en el campo experimental del Colegio de Postgraduados-Campus Veracruz, en la región centro del estado de Veracruz, México.
Materiales y Métodos
Área de estudio. La investigación se llevó a cabo en el banco de germoplasma de J. curcas L. no tóxicas, ubicado en el campo experimental del Colegio de Postgraduados-Campus Veracruz, en la región centro del estado de Veracruz, México. Esta instalación se localiza en el km 88,5 de la carretera Federal Xalapa-Veracruz, Colonia Tepetates, municipio Manlio Fabio Altamirano, Veracruz, vía Paso de Ovejas. Su ubicación geográfica corresponde a 19° 11’ 8.62’’ N y 96° 20’ 31.26’’ O, a una altitud de 24 m. El clima es del tipo Aw (w) (i) g, que se corresponde con el clima subhúmedo de lluvias en verano. La precipitación y temperatura media anual es de 1 239, 5 mm y de 25 °C, respectivamente, con menos de 5 % de precipitación en invierno y fluctuación de temperatura de 5 a 7 °C (García, 1988).
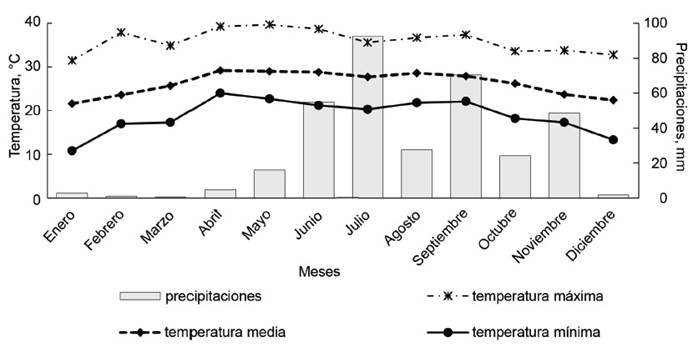
Características climáticas. Los datos de temperatura (máxima, media y mínima) y precipitaciones se obtuvieron de la Estación Meteorológica del Colegio de Postgraduados-Campus Veracruz, de enero a septiembre de 2019.
En la figura 1 se muestra el comportamiento de las temperaturas (máxima, media y mínima); además de las precipitaciones durante el período de evaluación (enero a septiembre). La temperatura máxima fue de 35,6 °C, y la mínima de 20,3 °C.
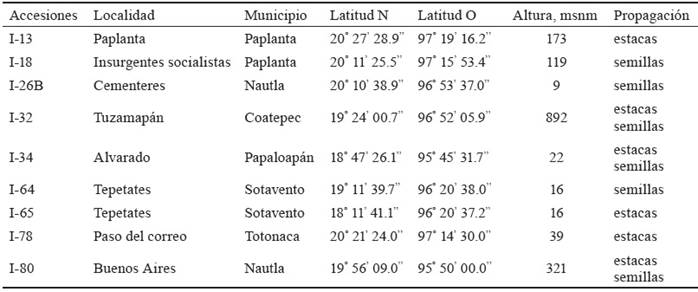
Material vegetativo. Se trabajó en una plantación de 49 accesiones (23 sembradas por semillas y 26 por estacas), con ocho años de edad, sembradas según diseño completamente al azar, recolectadas en el estado de Veracruz. De ellas, se seleccionaron seis propagadas por semillas y seis por estacas, con tres repeticiones. Para la selección se tuvieron en consideración los resultados de trabajos de investigación realizados por García (2015), en los que se evaluaron características morfológicas y productivas de estas accesiones, y los estudios de Zavala (2016), quien hizo una caracterización morfogenética en sus centros de origen. En la tabla 1, se muestran las accesiones evaluadas.
Diseño experimental. El experimento se estableció según diseño de bloques completamente al azar. Se consideró cada planta a muestrear como una réplica. Las plantas se sembraron en junio del 2011, a una distancia de 3 m entre surcos x 2 m entre plantas. La única práctica de manejo fue el control periódico de plantas arvenses (malezas).
Variables morfológicas evaluadas. Se tomaron datos de las plantas propagadas por semillas y por estacas (varetas), usando los descriptores indicados por Campuzano (2009), Laviola et al. (2009) y por la Red de Jatropha spp. (SAGARPA y SNICS, 2014). La variable altura de la planta (AP) se midió desde la base del tallo hasta el ápice, con una regla graduada en cm. El diámetro o grosor del tallo (DT) se determinó con un pie de rey (mm), a 10 cm por encima del nivel medio del suelo. El número total de ramas se obtuvo al contar las de cada una de las plantas muestreadas. Las mediciones antes mencionadas se realizaron en julio y septiembre, después de las lluvias.
También se contabilizó el número de ramillas y su longitud (mm), la cantidad de inflorescencias y su longitud (mm) y el número de racimos (NRac) y de frutos (NF) por planta. Estos últimos se muestrearon con frecuencia semanal. Se midió la longitud (LF), el ancho (AF) y el espesor (EF) según la metodología descrita por Laviola y Macedo (2009). Además, se contabilizó el número de semillas por fruto y el número de semillas por fruto. Las semillas se extrajeron de los frutos de forma manual, y una vez secas se tomó una muestra al azar (50). Con un pie de rey se midió la longitud, el ancho y el espesor (mm).
Plagas y enfermedades. Se estimó el porcentaje de enfermedades en cada uno de los árboles evaluados (infestación producida por hongos, virus, o por ambos, en la planta entera) y el porcentaje de daños producidos por insectos, potencialmente plaga. Se utilizó una escala de cuatro grados:
Grado
Rango de lesiones
0 a 1 % (inmune)
2 a 10 (resistente)
11 a 20 % (tolerante)
> de 20 % (susceptible)
Análisis estadístico. Los datos se procesaron mediante un análisis de componentes principales (ACP). Se tomó como criterio de análisis las componentes principales que presentaron valores propios superiores a 1, y factor de suma o de preponderancia mayor que 0,70 (Morrison, 1967). Se aplicó el análisis de conglomerados para la agrupación y selección de las accesiones, utilizando como índice de similitud la distancia euclidiana, a partir de lo obtenido en el ACP (Torres et al., 2006). Se determinaron los estadígrafos media y desviación estándar para las variables analizadas. Se utilizó, además, el análisis de correlación lineal para conocer la interrelación entre las variables. Todos los análisis se realizaron mediante el programa estadístico SPSS® versión 22,0 para Microsoft® Windows®.
Resultados y Discusión
En la tabla 2 se muestran los resultados del análisis de componentes principales. Se detectó una varianza acumulada de 91,65 % en las cinco primeras componentes. Las variables que mejor explicaron la varianza en la primera componente (34,20 %) fueron la cantidad, la longitud y el ancho de los frutos, además de la cantidad de semillas y sus dimensiones. La segunda componente extrajo una varianza de 19,96 %, explicada por la altura, el número de ramas y el diámetro (estos dos de forma negativa).
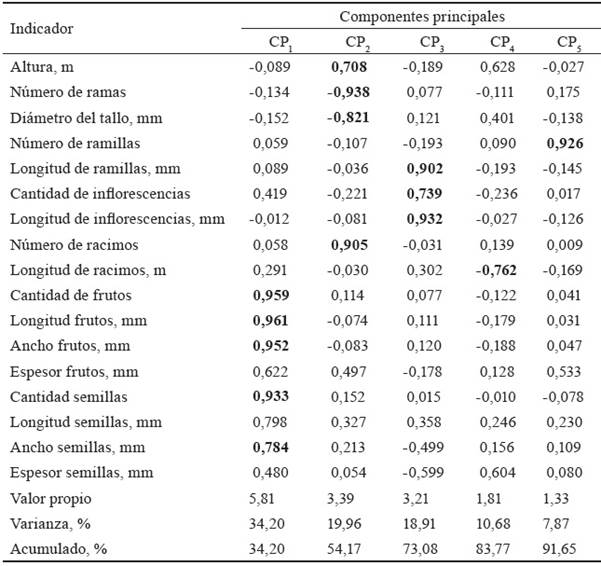
En la formación de la tercera componente influyeron la longitud de las ramillas, la cantidad y longitud de las inflorescencias, que explicaron 18,91 % de la varianza. Para la cuarta componente, se extrajo el 10,68 % de la varianza, fundamentada por la longitud de los racimos (de forma negativa); mientras que la quinta componente se explicó por el número de ramillas, y extrajo 7,87 % de la varianza.
El valor de la varianza de la primera componente (34,20) fue bajo, lo que evidencia la complejidad de las correlaciones entre las variables; es decir, existe alta variación entre los caracteres de las plantas, lo que también se relaciona con una gran diversidad morfológica, pues cada una de ellas puede que tenga un determinante propio, diferente en la productividad. No obstante, este análisis es válido, ya que cada una de las componentes seleccionadas tuvo un valor propio, superior a 1. Según los criterios de Kaiser (1960), ello supone considerar un factor que mejora la varianza proporcionada en un inicio para cada variable. Resultados similares, en cuanto al valor de la varianza, informaron Yengle-Ruíz (2012) en investigaciones realizadas para obtener índices sintéticos de calidad ambiental, así como Olivares (2014) en el diagnóstico socioambiental de un estudio de caso, y López-Roldán y Fachelli (2015), en trabajos de metodología de la investigación social cuantitativa.
Las variables espesor de los frutos y de las semillas no se incluyeron en ninguna de las cinco componentes, ya que el valor del factor de suma o de preponderancia (0,622 y 0,604; respectivamente) fue inferior a 0,70. Por tanto, se pudiera prescindir de ellos, cuando se realicen otras evaluaciones en circunstancias semejantes a las de esta investigación.
Para que la variabilidad correspondiente a cada indicador esté mejor relacionada con cada eje, en correspondencia con este tipo de análisis, el valor propio debe ser uno o mayor que uno (Philippeau, 1986). Esto se pudo comprobar en este trabajo, en el que la variabilidad estuvo bien distribuida, ya que dicho indicador en todos los casos fue superior.
Al respecto, se debe considerar lo planteado por Machado (2011), quien refiere que por lo regular las plantas frutales y otras leñosas suelen poseer alto grado de variabilidad. Esto se debe a que son netamente heterocigóticas y alógamas (de polinización cruzada), situación que conduce a la segregación genética en la descendencia, como sucede en J. curcas.
Lo anterior pudiera indicar que, independientemente de las condiciones edafoclimáticas, las poblaciones y, en particular, las accesiones de J. curcas (no tóxicas), pudieron expresar marcada variación entre individuos para algunos indicadores y agruparse en función de esas variables, lo que puede representar un elemento positivo en el trabajo de caracterización y evaluación. En ello pudo influir la variabilidad interespecífica e intraespecífica de la muestra estudiada, ya que estuvo conformada por varias accesiones recolectadas de disímiles ambientes y sembradas de forma diferente (por semilla y por estaca). También varían de forma marcada desde el punto de vista morfológico, y según las condiciones climáticas en las que transcurre la evaluación. Este es el caso de las diferencias que existen con respecto a las condiciones en que se desarrollaron los estudios de biología floral, cuantificación y medición de los componentes florales y su relación con el clima, realizados por García (2015). Este autor identificó los descriptores morfológicos relacionados con la producción en accesos de J. curcas, recolectados en diferentes regiones del estado de Veracruz, y propagados por semillas y estacas.
El valor alcanzado por la varianza acumulada y el valor propio de las componentes, permitió que casi todos los indicadores se incluyeran en el análisis de conglomerados, y poder determinar así la diferenciación o similitud entre las accesiones. Se exceptuaron las variables espesor de los frutos y de las semillas, ya que no contribuyeron a la expresión de la variabilidad de las componentes.
El análisis de conglomerados sobre la base de los resultados del ACP permitió la formación de tres grupos. Las accesiones pertenecientes a cada uno de ellos se muestran en la tabla 3, al igual que la media y la desviación estándar de cada uno de los grupos.
Tabla 3 Distribución de los individuos, media y desviación estándar, según el análisis de conglomerados.

E: sembradas por estacas
Los valores más altos, para la cantidad y la longitud de las ramillas y de las inflorescencias, la longitud de los racimos, el número y el ancho de los frutos, así como de la cantidad de semillas y sus dimensiones, se encontraron en el grupo III. Este último estuvo formado por las accesiones I-32 E, I-65 E (plantas sembradas por estacas). Le siguió el grupo II (I-32, I-34, I-18, I-80, I-13E, I-80E y I-78E), en cuanto al número de ramas, el diámetro del tallo, la cantidad de ramillas y la longitud de los frutos. En lo que respecta a la altura y cantidad de racimos, las cifras más altas correspondieron al I, representado por las accesiones I-26B, I-64, I-34E.
Como se puede observar, en los grupos I y II, se encuentran accesiones de plantas propagadas por semillas y por estacas. Esto pudiera indicar que en las poblaciones de esta especie se pueden encontrar genotipos, cuyo desarrollo se produce de forma rápida, mientras que otros son un poco más lentos, sin tener en cuenta que se sembraron o propagaron de manera directa por semilla, método que presupone un mejor establecimiento del vegetal. Por ello, independientemente de las diferencias que se observaron entre los indicadores, se debe señalar la evidente variabilidad que existe en la población de J. curcas.
Resultados similares registraron para esta especie Saadaoui et al. (2015), en Tunisia, al evaluar cinco caracteres cualitativos (área de la superficie foliar, longitud y ancho de la hoja, longitud del pecíolo y número de nodos), los que mostraron alta variabilidad morfológica (p < 0,05) en las ocho accesiones estudiadas. Saikia et al. (2015) encontraron variación moderada en la altura de la planta, el diámetro del tallo, el número de ramas por planta y el peso de la semilla.
Gwafila et al. (2019), en estudios de caracterización morfoagronómica y molecular de un germoplasma de J. curcas, en Botswana, también hallaron diferencias significativas entre las accesiones, para los indicadores cualitativos y cuantitativos. Chakrabarty et al. (2019), en investigaciones realizadas en 45 genotipos, en Bangladesh, encontraron significativa variación genética para los 17 indicadores morfoagronómicos evaluados.
En la tabla 4 se presentan los resultados del análisis de correlación entre los indicadores de las plantas evaluadas y los factores ambientales, por la importancia que se les atribuye a las interrelaciones entre estos últimos y las características morfológicas y productivas de las plantas.
Tabla 4 Matriz de las correlaciones fenotípicas entre los indicadores evaluados y los factores ambientales.
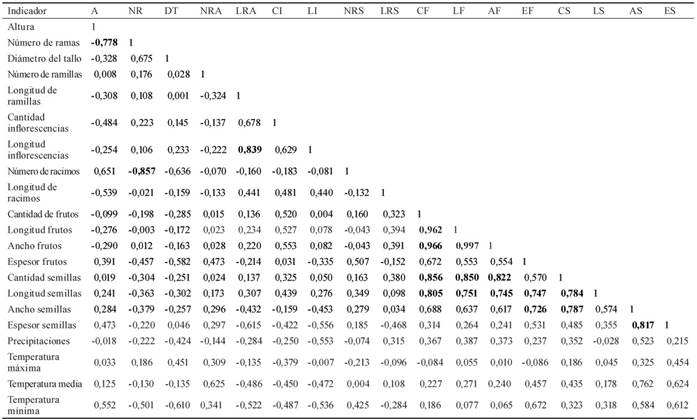
A: altura (mm), NR: número de ramas, DT: diámetro del tallo (mm), NRA: número de ramillas, LRA: longitud de ramillas (mm), CI: cantidad de inflorescencias, LI: longitud de inflorescencias (mm), NRS: número de racimos, LR: longitud de racimos (mm), CF: cantidad de frutos, LF: longitud frutos (mm), AF: ancho frutos (mm), EF: espesor frutos (mm), CS: cantidad semillas, LS: longitud semillas (mm), AS: ancho semillas (mm), ES: espesor semillas (mm) .
Existieron correlaciones fuertes entre la altura y el número de ramas (-0,778 de forma negativa). Y entre esta última y la cantidad de racimos (-0,857). Las hubo también entre la longitud de las ramillas y la de las inflorescencias (0,839), entre la cantidad de frutos con su longitud (0,962) y su ancho (0,966); así como entre la cantidad (0,856) y la longitud de las semillas (0,805).
De igual forma, se encontraron altas correlaciones entre la longitud del fruto y su ancho (0,996), la cantidad (0,850) y la longitud (0,751) de las semillas. Fueron elevadas las correlaciones entre el ancho del fruto con la cantidad (0,822) y la longitud (0,745) de las semillas, y entre el espesor del fruto y la longitud (0,749) y el ancho (0,726) de las semillas. Este comportamiento se repitió entre la cantidad de semillas con su longitud (0,784) y su ancho (0,787), y entre este último con su espesor (0,817).
Se observaron correlaciones moderadas entre la altura y la temperatura mínima (0,552); entre esta última y el número de ramas (-0,501), el diámetro del tallo (-0,610), la longitud de las ramillas (-0,522) y de las inflorescencias (-0,536). También entre el espesor del fruto (0,672) y el ancho (0,584) y espesor (0,612) de las semillas; entre la temperatura media y el número de ramillas (0,625), el ancho (0,762) y el espesor (0,624) de las semillas; y entre las precipitaciones y la longitud de las inflorescencias (-0,553) y el ancho (0,523) de las semillas.
Estos resultados son similares a los encontrados por Araiza-Lizarde et al. (2016), quienes plantearon que las temperaturas más altas no favorecieron el crecimiento de los individuos de J. curcas, a diferencia de la precipitación y la humedad relativa.
Características morfológicas de los frutos y las semillas. Se pudo corroborar lo planteado por Hidalgo y Grández (2013), en lo que respecta al fruto de esta especie. Se trata de una cápsula drupácea y ovoide. Después de la polinización se forma una fruta trilocular de forma elipsoidal. Al inicio es de color verde y carnoso, pero al madurar va cambiando su color a amarillo, hasta volverse de color café, oscuro o negro.
Solo se cosechó una gran cantidad de frutos de dos de las accesiones seleccionadas de J. curcas (no tóxica), que pertenecen a las plantas propagadas por estacas o varetas (I-32 e I-65). Estas también se destacaron, en cuanto a los indicadores cantidad de semillas por frutos, longitud, ancho y volumen, al igual que I-32, propagada por semillas.
Este comportamiento, en cuanto a la producción de frutos, se pudiera asociar a características de orden fisiológico, genético o de respuesta de las plantas a las condiciones ambientales, que no fueron las más idóneas (figura 1) para el buen desarrollo del cultivo; a pesar de que esta especie es capaz de desarrollarse en condiciones limitantes (Pérez-Vázquez et al., 2013). Otro factor que pudo haber influido fue la falta de manejo agronómico (en esta plantación de ocho años solo se realizó la chapea manual de las plantas arvenses), al que no todas las plantas responden de la misma forma.
Por otra parte, se observó la presencia de lepidópteros, trips, saltahojas (Empoasca sp.). Se encontró, además, infestación producida por hongos, presumiblemente de los géneros Colletotrichum y Cercospora. También se comprobó un síntoma de mosaico, con agarrotamiento en las hojas y en las nuevas inflorescencias, causado posiblemente por un agente viral.
El grado de afectación ocasionado por enfermedades fue de medio a bajo (entre 15 y 20 %), y su incidencia se manifestó en las accesiones propagadas por semillas y por estacas. No obstante, el engarrotamiento de las hojas fue más acentuado entre las propagadas por semillas.
Quiroga-Madrigal et al. (2014) refirieron síntomas de antracnosis, causados por Colletotrichum gloesporoides (Penz) y por Colletotrichum circinans (Berk) en el peciolo, las flores, las hojas y los frutos; así como de mancha foliar por Cercospora sp. Sacc, para las plantaciones de J. curcas en el sur de México.

