










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkPastos y Forrajes
versão On-line ISSN 2078-8452
Pastos y Forrajes vol.41 no.2 Matanzas apr.-jun. 2018
RESEARCH WORK
Diversity of the edaphic mesofauna in three soil uses in the Mayabeque province, Cuba
Ana América Socarrás-Rivero
Instituto de Ecología y Sistemática, Carretera de Varona km 3 ½, Capdevila, Boyeros, CP 11900, La Habana, Cuba
E-mail: anameri@ecologia.cu
ABSTRACT
The objective of the study was to characterize the mesofauna diversity in three soil uses in the Güines municipality Mayabeque province, Cuba. The studied systems were: silvopastoral system, secondary forest and cultivated pastureland. Five samples were taken in the first 10 cm of soil in each, during the rainy season. For the data analysis range/abundance curves were constructed, and the community coefficient (CC) and proportional similarity coefficient (PS) were calculated. A total of 399 edaphic microinvertebrates belonging to two classes, five orders and 19 families, were collected. The best represented order was Oribatida. The dominance-diversity curves showed, in the silvopastoral system, a more homogeneous distribution of its groups. In the secondary forest the highest quantity and diversity of detritivorous and exclusive families was obtained. In the secondary forest and the silvopastoral system, the CC showed the highest similarity in the composition of its taxa. In addition, the PS confirmed the similarity among the abundance values of both systems. In the use pastureland the minimum abundance and diversity values of these fauna groups were obtained, corroborated by CC and PS. The detritivores/predators and Oribatida/Astigmada ratios were favorable for detritivores in the three soil uses. It is concluded that the secondary forest and the silvopastoral system facilitate the re-colonization of edaphic communities and the conservation of their function; in addition, systems with trees contribute to the conservation of the soil biological quality.
Keywords: secondary forest, predators, Leucaena leucocephala.
INTRODUCTION
Cuban soils aimed at animal husbandry are subject to different disturbances, among which the following stand out: strong seasonality, low nutrient availability, variable grazing pressure, as well as fire incidence (Oumarou, 2015). A strategy for their amelioration is sowing trees, such as Leucaena leucocephala (Lam), in paddocks aimed at grazing, which guarantees shade and feed for the animals. The soils where silvopastoral systems are established improve their physical, chemical and biological properties; increase the content of organic matter and nitrogen; and broad cover is guaranteed. This allows higher biological diversity of the edaphic medium; nevertheless, their disturbance degree, which is caused by the intrinsic processes of animal husbandry management, should be taken into consideration.
In the soil different biological processes occur, among them the activity of the biota, which, along with climate factors, plays an important role in the transformations of organic matter. Mesofauna, as part of this biota, intervenes in the decomposition of organic matter, in the acceleration and recycling of nutrients and in the process of phosphorus and nitrogen mineralization, decisive factors for the maintenance of soil productivity (Delli and Flores-Torres, 2016). In addition, it participates directly within the trophic networks, because the mesofauna organisms are the main controllers of the fungi and bacteria populations (Tome et al., 2015). Many of these groups work as bioindicators of soil stability and fertility, being sensitive to climate change and anthropic disturbances of the edaphic medium, which causes variations in their density and diversity (Mahdi et al., 2016).
For such reason, the objective of this study was to characterize the mesofauna diversity in three soil uses in the Güines municipality Mayabeque province, Cuba.
MATERIALS AND METHODS
The study was conducted in areas of the Institute of Animal Science, Güines municipality Mayabeque province, Cuba, on two soil types: Brown with Carbonates and reddish Fersialitic Brown (Hernández-Jiménez et al., 2015), in the rainy season.
The evaluated systems were:
- Cultivated pastureland. Located on a Brown soil with Carbonates, had been exploited for more than 40 years with organic amendments. The paddock occupied an area of 0,75 ha, with 80 % of soil cover. As the main pasture species Cynodon nlemfuensis (star grass) was cultivated, and the area was managed with Voisin rational grazing.
- Silvopastoral system. It was located on a Brown soil with Carbonates, with 11 years of exploitation. It had two strata: a tree one, composed by L. leucocephala with more than 2 m of height; and an herbaceous one, with the grass Brachiaria decumbens. The grazing regime was similar to the one that was carried out in the cultivated pastureland system. No chemical fertilization was applied, only the cattle manure from the livestock that grazed in the system was incorporated.
- Secondary forest. Located on a reddish Fersialitic Brown soil, it occupied a surface of 2 ha to which the animals did not enter; for 15 years it had been in a natural regeneration process, with semideciduous vegetation. The prevailing species were L. leucocephala and Trichilia hirta.
Sampling of the edaphic mesofauna. In each area five soil samples were taken at only one depth (0-10 cm), with 5 cm of diameter cylinder and 10 cm of depth, following a completely randomized sampling design. For the extraction of the edaphic fauna Tullgren funnels were used, during seven days, without using any artificial source of light and heat. The individuals were counted and separated under the stereoscopic microscope; they were preserved in 70 % alcohol, and were identified to family level, following the classification proposed by Brusca and Brusca (2003) for insects and the one suggested by Krantz and Walter (2009) for mites.
Statistical procedure of the data. From this counting the density (ind/m2) of each taxon was obtained. In order to know whether there were differences in the density of the edaphic communities among the areas, as well as to determine their variations among uses, the Kruskal-Wallis test was applied; and in the cases in which the differences were significant, the Student-Newman-Keuls (SNK) test was used. The statistical processing was made through the package of the automated program TONYSTAT (Sigarroa, 1987).
To characterize the structure in each area dominance-diversity curves were constructed, from the decimal abundance logarithms (Magurran, 1989). The quantitative (QS) and qualitative similarity (CS) were calculated, according to Feisinger (2004). The biological indicators were selected taking into consideration the ecological characteristics of the present organisms and adopting criteria proposed by other researchers (Bedano, 2010; Socarrás, 2013).
RESULTS AND DISCUSSION
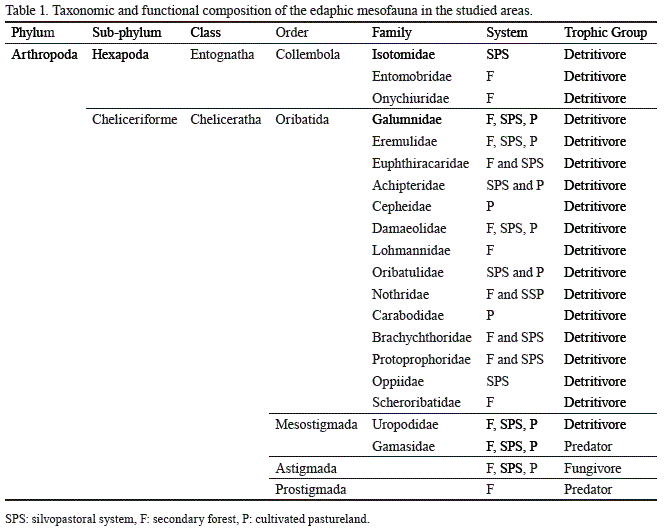
Taxonomic composition of the soil mesofauna in the areas. During the study, a total of 399 individuals belonging to the phylum Arthropoda, represented by two sub-phylum, two classes, five orders and 19 families, were collected. In particular, the class Cheliceratha was composed by four orders and 16 families. The analysis at order level showed the existence of 5 orders in the secondary forest, 4 in the silvopastoral system and 3 in the pastureland (table 1).
The taxonomic richness, evaluated at the level of identified families, showed the same number in the secondary forest and in the silvopastoral system (13); while in the cultivated pastureland 9 families were reported.
The best represented order in number of families in the three systems was Oribatida, with 9 in the secondary forest, 10 in the silvopastoral system and 7 in the cultivated pastureland. Likewise, 4 families exclusive of the secondary forest were found: 2 in the silvopastoral system and 2 in the cultivated pastureland (fig. 1).
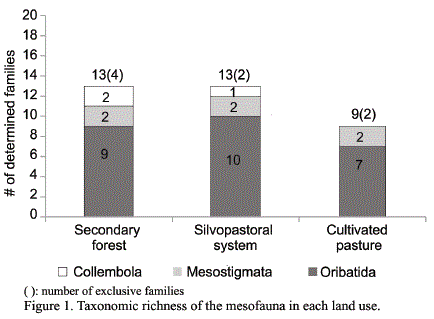
Analysis of dominance-diversity of the soil mesofauna. The range-abundance curves showed a higher number of groups for the forest and the agroforestry system, compared with the pastureland (fig. 2).
The curve related to the number of individuals in the forest showed that the prevailing family was Uropodidae, which is reported as a group of detritivore mites, with morphological and bioecological characteristics that make them highly demanding regarding the habitat quality (Bedano, 2010; Socarrás, 2013). Their prevalence in this area could have been favored by the presence of material to be decomposed and higher soil cover, which guarantees balanced conditions of humidity and temperature in the edaphic medium. In the forest several taxa with moderate abundance appeared: Astigmada, Gamasidae, Galumnidae, Scheloribatidae, Eremulidae and Euphthiracaridae; and as very rare groups, in which one or two individuals were captured: Prostigmada, Damaeolidae, Nothridae, Entomobridae, Onychiuridae, Brachychthoridae, Lohmannidae and Protoprophoridae. The presence in this area of indicative families of edaphic maturity or the so-called «exclusive» taxa» can be emphasized (Euphthiracaridae, Brachychthoridae and Protoprophoridae), which are more sensitive to the disturbances of the environment.
In the cultivated pastureland Eremulidae prevailed, along with Uropodidae, first; and the groups that had only one individual belonged to the families Achipteridae, Cepheidae, Damaeolidae and Carabodidae. In this area the abundance was concentrated in few taxa, pattern that proved the low equitability of the communities in this pastureland. Magurran (1989) stated that the groups with higher number of individuals occupy a large proportion of the niche, and make higher utilization of the available resources.
The expression of communities with marked dominance in the families showed that, in the cultivated pastureland and the secondary forest, disturbing factors were having incidence. In the former there was influence of grazing and the lack of tree cover; while in the latter the establishment time of the system and the competition between taxa over the resources could have influenced.
In the silvopastoral system Uropodidae stood out numerically. These mites are abundant in ecosystems with high OM values, in composting areas and in decomposing trunks (Rousseau et al., 2013). They are humicolous and respond positively to good soil aeration conditions. The study of their variations constitutes an accurate criterion of the health status of the edaphic medium.
This system has more heterogeneous resource sources, and also the tree cover provides a higher contribution of litter and shade to maintain stable the temperature and humidity values in the soil, all of which favors the development of more diverse, cosmopolitan and pioneer communities of disturbed media. Among the rare groups, with one or two individuals, Protoprophoridae, Oribatulidae and Isotomidae can be indicated. From the three studied areas, this one showed the curve with less abrupt slopes, which indicates higher equity among the taxa of the edaphic mesofauna, with predominance of groups with moderate abundance; and showed higher equitability, suggesting that the resources were distributed more homogeneously within the community (Pauli et al., 2012).
According to the expression of the abundance range curves, the secondary forest could be mentioned first; the silvopastoral system secondly, with higher diversity; and the cultivated pastureland as the use system with lower diversity of the soil mesofauna.
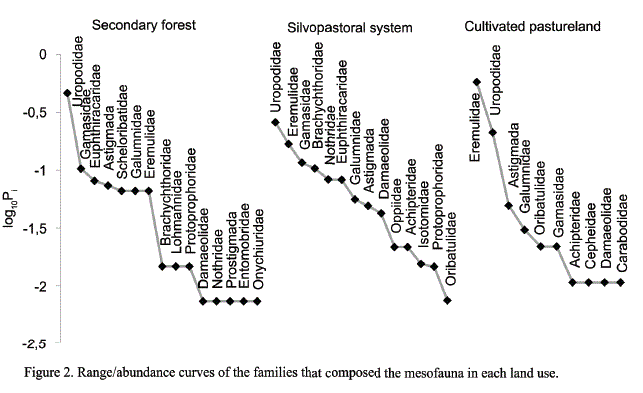
Density of the soil mesofauna in the areas. The highest average density values of the soil mesofauna (fig. 3) were found in the secondary forest (23 075 ind.m-2); the moderate ones, in the silvopastoral system (19 343 ind.m-2); and the lowest ones, in the pastureland (13 361 ind.m-2).
The Kruskal-Wallis test showed significant differences among the different uses in the case of density (H =1 3,59; p < 0,001; gl = 3). The variations in the mesofauna density between the secondary forest and the silvopastoral system could be ascribed to the vegetation structure, the natural input of organic matter to the medium and the quality of the litter contributed to the soil.
The strategy of planting trees for animal feeding, such as L. leucocephala, improves the structural stability of the edaphic medium, prevents erosion and water infiltration, increases edaphic biodiversity, and decreases the organic losses in the soil due to higher cover (Pérez, 2011).
A few authors state that the high density values of the soil fauna in the systems with L. leucocephala could be given by the presence of two strata: a tree one with the legume and an herbaceous one with native pasture, a higher quantity of better-quality litter and a more favorable microenvironment for the activity of the edaphic fauna (García, 2013; Socarrás and Izquierdo, 2014).
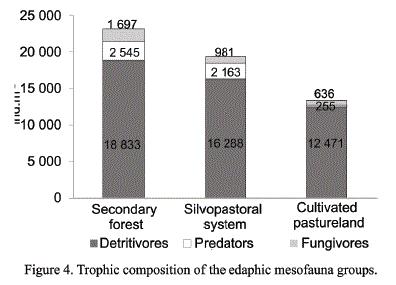
Trophic composition of the components of the edaphic mesofauna. The community of soil microarthropods was composed by three trophic categories in all the studied systems: detritivores, predators and fungivores. These categories play an important role in the ecological stability of the soil.
The increase in the number of detritivore individuals with regards to the other trophic groups in the three areas, with a peak in the secondary forest and in the silvopastoral system (fig. 4), should be highlighted. In the case of the forest, it has been favored by the scarce or null anthropic intervention, which has resulted in higher stability of the edaphic medium and its associated diversity. In the silvopastoral system, the establishment of this trophic group was favored by the incorporation of organic matter, the cover and improvement of the fertility conditions by the presence of L. leucocephala, which is a legume that fixes atmospheric nitrogen to the soil.
The second trophic group that stood out for its abundance in all the areas was that of predators, due to a direct relation with their prey. The groups that formed this category were predators in immature states of Oribatida and Collembola (detritivores), which were abundant, but maintained the prey-predator relation; and it represented a balance in the ecosystem (Schiavon et al., 2015).
Fungivores (Astigmada) are constituted by microarthropods, indicators of soil instability and infertility; the found values did not have marked differences, due to the season, edaphic conditions and system type. Socarrás and Robaina (2011) found response patterns for such trophic groups similar to the ones obtained in this study, with similar soil characteristics, season and land use.
Analysis of the similarity indexes of the soil mesofauna. The community coefficient (CC) showed that the secondary forest and the silvopastoral system had a higher similarity in their taxonomic composition. On the other hand, it showed that the uses secondary forests and cultivated pastureland shared less taxa (table 2).
With the analysis of proportional similarity (PS), the likeness in the abundance values between the secondary forest and the silvopastoral system was corroborated, with marked differences with regards to the values obtained for the cultivated pastureland.
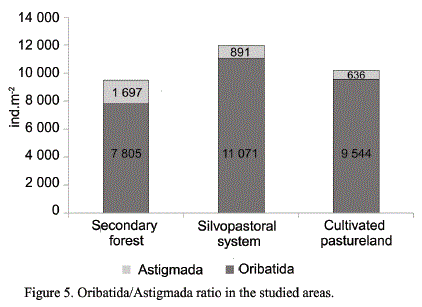
Biological indicators. Oribatida and Astigmada are two orders that compose the soil mesofauna and that perform antagonist functions; while one increases, the other decreases; hence the importance of this balance to measure the unbalance degree among the edaphic biocenosis (Socarrás and Izquierdo, 2014).
In the studied systems this relation was favorable to Oribatida, detritivore group of mites, indicator of soil stability and fertility, for which it can be stated that the edaphic medium was benefitted with these uses (fig. 5). The secondary forest as well as the silvopastoral system guaranteed the biotic and abiotic conditions for the development of mesofauna; regarding the cultivated pastureland, this use also ensured good soil cover and a contribution of organic material that resulted in the good establishment of the edaphic mesofauna.
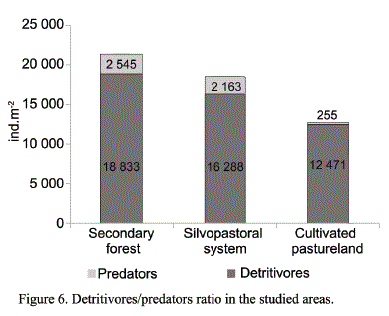
The detritivores/predators ratio has been validated by Hernández et al. (2012), and expresses the stability of edaphic communities. In this study the ratio was higher than one, that is, detritivores prevailed in all the areas, which confirms the existing balance between the prey-predator relation and the stability of the ecosystem (fig. 6).
CONCLUSIONS
- When making an analysis of the variation of the mesofauna components in the three systems, an affectation was observed in the sense secondary forest '! silvopastoral system '! pastureland, due to the decrease of density; as well as because of the changes in the functional structure of the populations, in the richness and the loss of exclusive families.
- The secondary forest and the silvopastoral system facilitated the recolonization of the edaphic communities and the conservation of their function.
- Systems with trees contribute to the conservation of the soil biological quality.
Received: August 5, 2017
Accepted: February 4, 2018
1. Bedano, J. C. La importancia de la mesofauna y macrofauna edáfica y su uso en la evaluación de la calidad del suelo. Memorias del XXII Congreso Argentino de la Ciencia del Suelo, II Reunión de Suelos de la región Andina. San Luis, Argentina: INTA Balcarse. p. 5, 2010.
2. Brusca, R. C. & Brusca, G. J. Invertebrates. Sunderland, USA: Sinauer Associates, 2003.
3. Delli, A. & Flores-Torres, María E. Estudio comparativo de indicadores físicos y químicos de la calidad del suelo y de la biodiversidad de la mesofauna edáfica en dos usos de suelo de la microcuenca del río Pomacocho, parroquia Achupallas, cantón Alausí, provincia de Chimborazo. Riobamba, Bolivia: Universidad Nacional de Chimborazo, 2016.
4. Feisinger, P. El diseño de estudios de campo para la conservación de la biodiversidad. Santa Cruz de la Sierra, Bolivia: Editorial FAN, 2004.
5. García, Y. La macrofauna edáfica como indicador de la calidad del suelo Ferralítico Rojo en cuatro usos de la tierra en la provincia de Matanzas. Tesis en opción al grado científico de Máster en Ciencias. San José de las Lajas, Cuba: Universidad Agraria de La Habana, 2013.
6. Hernández, Guillermina; Socarrás, Ana A.; Cabrera, Grisel; Alguacil, María M.; Torrecillas, Emma & Roldán, A. Impacto de la siembra de plantas con fines bioenergéticos sobre la biodiversidad edáfica. En: A. Valdés y M. A. Vales, eds. Análisis del impacto de la producción de biocombustibles en el medio natural. La Habana: Editorial Academia. p. 51-77, 2012.
7. Hernández-Jiménez, A.; Pérez-Jiménez, J. M.; Bosch-Infante, D. & Castro-Speck, N. Clasificación de los suelos de Cuba. Mayabeque, Cuba: Instituto Nacional de Ciencias Agrícolas, Instituto de Suelos, Ediciones INCA, 2015.
8. Krantz, G. W. & Walter, D. E., Eds. A manual of acarology. 2nd ed. USA: Texas Tech University Press, 2009.
9. Magurran, Anne E. Diversidad ecológica y su medición. Barcelona: Ediciones Vedra, 1989.
10. Mahdi, J. E.; Abbott, Lynnette K.; Pauli, Natasha & Solaiman, Z. M. Biological indicators for soil health: potential for development and use of on-farm tests. In: A. Varma and A. K. Sharma, eds. Modern tools and techniques to understand microbes. Switzerland: Springer. p. 123-134, 2016.
11. Oumarou, I. Efecto del uso del suelo sobre indicadores del estado de la calidad del suelo en áreas cañeras de la CPA «Batalla de Santa Clara». Tesis de pregrado. Santa Clara, Cuba: Facultad de Agronomía, Universidad Central de Las Villas, 2015.
12. Pauli, N.; Barrios, E.; Conacher, A. J. & Oberthür, T. Farmer knowledge of the relationships among soil macrofauna, soil quality and tree species in a smallholder agroforestry system of western Honduras. Geoderma. 189-190:186-198, 2012.
13. Peréz, D. Cambios en el estado de un suelo de composición Ferralítica por el manejo diferenciado de la cobertura. Tesis en opción al grado científico de Máster en Ciencias: Universidad Agrarias de La Habana, 2011.
14. Rousseau, L.; Fonte, S. J.; Téllez, O.; van der Hoek, R. & Lavelle, P. Soil macrofauna as indicators of soil quality and land use impacts in smallholder agroecosystems of western Nicaragua. Ecol. Indic. 27:71-82, 2013.
15. Schiavon, Greice de A.; Lima, Ana C. R. de; Schiedeck, G.; Schwengber, J. E.; Schubert, R. N. & Pereira, Caroline V. O conhecimento local sobre a fauna edáfica e suas relações com o solo em agroecosistema familiar de base ecológica: um estudo de caso. Ciência Rural. 45 (4):658-660, 2015.
16. Sigarroa, A. Paquete estadístico para BASIC. Cuba: Facultad de Biología, Universidad de La Habana, 1987.
17. Socarrás, Ana A. Mesofauna edáfica: indicador biológico de la calidad del suelo. Pastos y Forrajes. 36 (1):5-13, 2013.
18. Socarrás, Ana A. & Izquierdo, I. Evaluación de sistemas agroecológicos mediante indicadores biológicos de la calidad del suelo: mesofauna edáfica. Pastos y Forrajes. 37 (1):47-54, 2014.
19. Socarrás, Ana A. & Robaina, Nayla. Mesofauna edáfica en diferentes usos de la tierra en la Llanura Roja de Mayabeque y Artemisa, Cuba. Pastos y Forrajes. 34 (3):347-358, 2011.
20. Tome, S. A.; Cutz, L. Q. & Ortíz, H. J. Variación espacio-temporal de ácaros (Cryptostigmata) en puntos estratégicos de la bahía de Chetumal Quintana Roo, México. Ingeniantes. 1 (2):58-64, 2015.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}