










 Serviços customizados
Serviços customizados Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
Globally, diabetes mellitus (diabetes) and its complications are among the leading causes of morbidity and mortality, making it a growing problem for public health and national economies.1 In Cuba, diabetes has increased in incidence and prevalence and is among the top 10 causes of death at all ages.2
Pregnant women with pre-gestational diabetes and inadequate glycemic control have a high incidence of abnormalities of embryonic, fetal, and placental development; congenital malformations are common and increase perinatal morbidity and mortality.3,4,5,6
The mechanisms by which complications occur in the gestation of diabetic women are not fully clarified, but they are related to the pro-oxidant and pro-inflammatory intrauterine environment caused by metabolic disorders. Maternal hyperglycemia is associated with oxidative stress, a hypoxic state of the embryo, and changes in the regulation of apoptotic pathways, all of which alter crucial signaling events during prenatal development, the expression of genes related to morphogenesis, and the integrity of genetic material.4,5,6
Erythropoietin (EPO) is a glycoprotein whose primary function is to stimulate erythropoiesis. However, EPO has also been shown to have cytoprotective effects in non-hematopoietic tissues due to angiogenic, anti-apoptotic, anti-inflammatory, neurotrophic, and antioxidant actions.7
On the other hand, there is evidence that EPO is expressed early in several of the structures of the embryo8 and that it stimulates proliferation and inhibits apoptosis in cells of the decidua and trophoblast,9 mechanisms necessary for adequate morphogenesis. In addition, it has been observed that recombinant human EPO (rhuEPO) reduces glycemia in conditions of hyperglycemia, so research has increased on its potential cytoprotective role in diabetes.10,11,12
Activation of EPO receptors in non-hematopoietic tissues requires higher EPO concentrations than those necessary to stimulate erythropoiesis. High doses of rhuEPO produce adverse effects associated with the stimulation of hematopoietic and pro-coagulant pathways,7 so derivatives have been developed to enhance cytoprotective effects, without hematopoietic actions.13
At the Center for Molecular Immunology of Cuba, a hyposialic variant known as NeuroEPO is obtained during the production of rhuEPO. The use of a nasal formulation of this molecule has shown neuroprotection in cerebral ischemia14,15 and neurodegenerative diseases,16,17 without adverse effects at the hematological level. In addition, our group of research has found that NeuroEPO reduces hyperglycemia in diabetic rats.18
Considering the evidence of the cytoprotective effect14,15,16,17) and hypoglycemic effect18) of NeuroEPO, the involvement of EPO in embryonic development,8,9 and that there are reports that rhuEPO can cross the placenta,19 the objective is to evaluate the protective effect of NeuroEPO on the reproduction of diabetic rats.
Material and Methods
An experimental study was carried out in Wistar rats obtained from the National Center for the Production of Laboratory Animals (CENPALAB) of Cuba, following the ethical precepts established.20,21 The research was developed in the Department of Biochemistry of the Institute of Basic and Preclinical Sciences "Victoria de Girón", in the period from 2019 to 2021. In each group, a number of animals similar to the smallest number used in previous experimental studies of diabetes and pregnancy in rats was used.22,23
Animals and environmental conditions
Female, fertile and virgin rats, with an initial weight of 200 g ± 20 g, kept in independent boxes, with constant cycles of light and dark, at a temperature of 21 ºC - 23 ºC, and with free access to standard food (CENPALAB) and filtered water were used for the study. The animals were kept for a week in adaptation to the environment before starting the procedures.
Induction of diabetes
To provoke diabetes in rats, intraperitoneal injection of streptozotocin (SIGMA), 65 mg/kg in 200 μL of sodium citrate buffer 0,1 M pH 4,5 was administered; one week later, glycemia was determined and rats with glycemia greater than 11 mM24 were considered diabetic. Glycemia was determined with a glucometer (SUMA) in blood obtained by a cut at the tip of the animal's tail.
Induction of pregnancy
Diabetic rats and a group of non-diabetic rats were mated at night, in the proestrus and estrus phases of the estrous cycle.25 Day 0 of gestation was considered when sperm were found in the vaginal wash performed in the morning after mating.
Study groups
On day 0 of gestation, the rats were randomly assigned to the study groups. Each group consisted of 8 animals.
D 0,5: diabetic rats receiving NeuroEPO 0,5 mg/kg.
D 0,75: diabetic rats receiving NeuroEPO 0,75 mg/kg.
D 1: diabetic rats receiving NeuroEPO 1 mg/kg.
DV: diabetic rats that received the vehicle.
C: non-diabetic rat control.
The NeuroEPO and the vehicle were supplied by the Center for Molecular Immunology of Cuba. Both preparations were administered subcutaneously in the dorsal region of the rats, once a day, on alternate days, starting on day 0 of gestation, for a total of 6 applications.
Euthanasia and sample collection
On day 12 of gestation, the rats were euthanized by bleeding (intracardiac puncture) preformed under anesthesia (thiopental sodium 50 mg/kg intraperitoneal). The uterine horns and ovaries were dissected and placed in a petri dish (100 mm) at room temperature. With the help of scissors, tweezers, and stereoscope (Motic), the periuterine tissue was removed, the wall of the uterus was opened along its entire length, and all membranes were carefully removed.26
Variables
Glycemia of diabetic rats, on days 0, 6, and 12 of gestation, was determined by glucometer (SUMA) in blood obtained by a cut at the tip of the tail. The results were expressed as the percentage of the initial glycemia (%), considered as 100 %,24 which corresponded to the glycemia of day 0 of gestation.
The counting of corpus luteums, implantations, resorptions, and embryos was carried out using a stereoscope and an optical microscope (Motic)(22.26) In the embryos, the presence or absence of malformations (neural tube closure defects; alterations in the cerebral, optical, and otic vesicles; the sketches of the extremities; the cardiac area; and the embryonic folding) were analyzed26 (Figure 1). The total losses (number of corpus luteums - number of embryos) were calculated, as well as the percentage of pre-implantation losses [(number of corpus luteums - number of implantations) X100/number of corpus luteums] and post-implantation [(number of implantations - number of embryos) X 100/number of implantations)].22
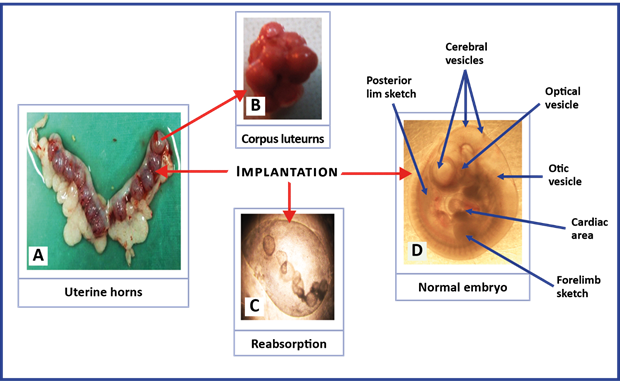
Fig. 1 Representative images of the structures for the analysis of reproductive variables and the study of malformations in rat embryos with 12 days of gestation. The images were taken with a digital camera coupled to Stereoscope in B (40X), and an optical microscope in C and D (100 X).
Statistical processing
GraphPad Prism software (version 5.01) was used. One-way and two-way analysis of variance (ANOVA) was used for comparisons of glycemic levels, and one-way ANOVA was used for the area under the curve; Bonferroni-type multiple comparisons were used in each case. One-way ANOVA tests and Newman-Keuls multiple comparisons were used for the analysis of the reproductive variables, and the Fisher's exact test was used for the comparison of proportions. There were significant differences with p values <0,05.
The datasets generated and/or analyzed during the current study are available in the Mendeley Data repository.27
The project was approved by the Scientific Council and the Research Ethics Committee of the institution.
Results
Diabetic rats in all groups began gestation with glycemic levels above 300 mg/dL (16,7 mM). During gestation, glycemia remained at similar levels in group DV, while in D 0,5 it was significantly reduced with respect to the initial glycemia and that of group DV, with a smaller area under the curve. The decrease in glycemia in D 0,5 was evident from day 6 of gestation and reached 27 % on day 12 of gestation. (Figure 2).
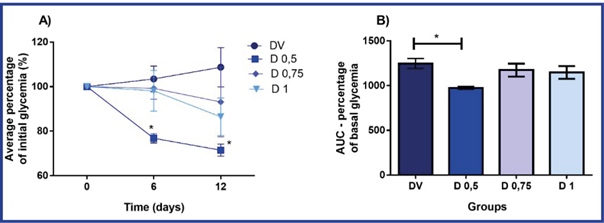
A) In each time, the percentage with respect to the initial glycemia (time 0) is shown. B) Area under the curve (AUC). The values of mean and standard deviation (n = 8 in each group) are presented.
DV: diabetic rats + vehicle; D 0,5, D 0,75 and D1: diabetic rats + 0,5 mg/kg, 0,75 mg/kg and 1 mg/kg of NeuroEPO, respectively. Subcutaneous route of administration, in alternative days, from day 0 to 10 of gestation (6 applications).
*p <0,05- Significant difference with respect to basal glycemia (A) and DV group (A and B). (Analysis of ANOVA variance of one and two tracks (A) and one-way (B), and Bonferroni test).
The study of reproductive variables in rats showed a similar number of corpus luteums in all groups. In group DV, the number of implantations and embryos were lower, and the pregnancy losses were greater when compared with group C. In group D 0,5 the results of all variables were similar to group C, with an increase in the number of embryos and a reduction in losses with respect to group DV. (Table 1).
In groups, D 0,75 and D1, the number of implantations, embryos, and losses was similar to those found in DV, with significant differences compared with both C and D 0,5. In addition, in D 0,75 and D1 the percentage of embryos per implantation was lower and the percentage of losses per corpus luteum was higher than that observed in the rest of the groups. (Table 1).
Table 1 Result of reproductive variables of diabetic rats that received different doses of NeuroEPO during pregnancy and its controls

N: number; SD: standard deviation. C: non-diabetic rat controls; DV: diabetic rats + vehicle; D 0,5, D 0,75 and D1: diabetic rats + 0,5 mg/kg, 0,75 mg/kg and 1 mg/kg of NeuroEPO, respectively. Subcutaneous route of administration, on alternate days, from day 0 to day 10 of gestation (6 applications).
The values of the mean ± SD (Analysis of variance ANOVA of a Newman-Keuls test pathway) and the percentage (%) (Fisher's exact test) are presented.
* p <0,05- Difference with C. # p <0,05- Difference with D 0,5. † p <0,05- Difference with C, DV and D 0,5.
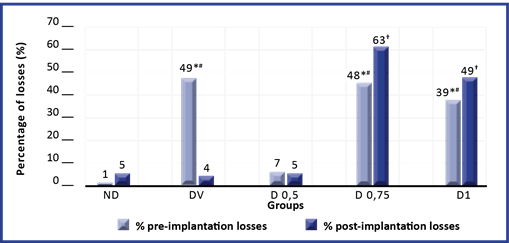
A more detailed study of the losses revealed that in groups DV, D 0.75, and D 1, in which an increase in the result of this variable was found (Table 1), the percentage of pre-implantation losses was higher than that observed in C and D 0,5. Also, when comparing the percentage of post-implantation losses, the DV group did not present differences with C and D 0,5, but groups D 0,75 and D1 showed an increase with respect to the rest of the groups. (Figure 3).
In the study of the embryos, no congenital malformations were identified in either group.
Discussion
In order to evaluate whether NeuroEPO offers protection in the reproduction of diabetic rats, its effect was investigated in a model of streptozotocin-induced diabetes. Rats injected with streptozotocin developed severe hyperglycemia, which was significantly reduced with repeated administration of 0,5 mg/kg of NeuroEPO. The result coincides with the reduction in hyperglycemia observed in diabetic rats after a single application of the same dose of NeuroEPO,18 which confirms the hypoglycemic effect of this Cuban biotechnology product.
In the previous report on the hypoglycemic effect of NeuroEPO, the results suggest an insulinotropic action of this molecule,18 which is consistent with research conducted with exogenous EPO in diabetic rats.10,11
Other studies have shown that EPO administered to diabetic rats reduces the intensity of hepatic gluconeogenesis10,28 and exerts a direct cytoprotective effect on pancreatic islets.11,29,30 Similar mechanisms may be involved in lowering glycemia in group D 0,5, which should be tested in future research.
On the other hand, it is known that hyperglycemia has harmful effects on the ovary and that in the model of streptozotocin-induced diabetes, the fertility of animals decreases progressively and irreversibly.31 However, no differences were observed between the groups in terms of the number of corpus luteums, which is an indirect indicator of the number of fertilized oocytes.22 The result may be because all diabetic rats became pregnant within two weeks of being diagnosed with diabetes, which may be a short time for ovaries damage to manifest.
The DV group, in addition to developing severe hyperglycemia maintained during the experiment, presented fewer implantations and embryos, as well as an increase in preimplantation losses, which are characteristic of the model used, widely used in diabetes and pregnancy studies.22,23,26,32 In this model it is shown that the alterations of reproduction depend on maternal hyperglycemia and not on streptozotocin; this substance is eliminated from circulation a few hours after its administration,31 so it does not affect reproduction when administered before gestation. However, embryonic structures exposed to an excess of glucose have a higher consumption of oxygen, which consolidates the hypoxic state that characterizes the early stages of embryonic development; the establishment of a hypoxic state produces oxidative stress, which is involved in the alterations of the reproduction of the diabetic mother.4,5,6
Alterations in the offspring of the diabetic mother are related to dysregulation of the processes of proliferation, differentiation, and apoptosis.4,5,6 Blastocyst cell apoptosis may lead to implantation failures,4 which result in an increase in preimplantation losses and a decrease in the number of embryos, as observed in the DV group.
Another feature of this model is the increase in post-implantation losses, which is equivalent to abortions and fetal deaths in diabetic pregnant women.3 However, in the present study, no increase in the percentage of post-implantation losses was observed in the DV group; this result could be because the analysis of this variable was carried out taking into account only the resorptions present in the gestational sacs. The absence of gestational sacs does not mean that there was no implantation, since losses may occur shortly after implantation,22 and no gestational sacs are detected at 12 days of gestation. Therefore, it is likely that part of the pre-implantation losses identified in the study corresponds to post-implantation losses.
Other results that suggest the possibility of an undetected increase in post-implantation losses in the DV group are the lower number of embryos and the absence of congenital malformations. It is known that embryos with severe malformations do not survive,4,5,6) so embryo losses may have occurred shortly after implantation.
In group D 0,5 there was an increase in the number of implantations and embryos, and a decrease in losses, to levels like those observed in non-diabetic rats. However, the higher doses of NeuroEPO evaluated (groups D 0,75 and D 1) did not prevent the negative effects of diabetes on reproductive variables and increased post-implantation losses (resorptions), with a lower percentage of embryos, which shows a harmful effect of high doses.
The beneficial effects observed in group D 0,5 may be due to the improved glycemic control of mother rats. However, it has been proven that, during embryonic development, EPO is not only indispensable for the maturation of the hematopoietic system but also for the proliferation and survival of other cell lines.8,9 In addition, there are reports that maternal EPO can cross the placenta.19 On the other hand, it has been shown that NeuroEPO produces neuroprotection in models of cerebral ischemia and neurodegenerative diseases by antioxidant, anti-inflammatory, and anti-apoptotic mechanisms.14,15,16,17 Therefore, it is possible that the NeuroEPO administered to diabetic rats during pregnancy reached the embryonic structures and exerted a direct cytoprotective effect in the control of processes involved in morphogenesis, with beneficial effects on reproduction.
In vitro and in vivo studies with NeuroEPO and other EPOs have shown bell-shaped concentration-response and dose-response curves.33,34,35 In addition, Rodríguez Cruz et al.17 reported that, in a model of Alzheimer's disease, the benefits of NeuroEPO were greater at the lowest dose. At the highest dose, some results showed more alterations than in the vehicle group, like what was observed in the present study.
As previously discussed, increased apoptosis is a mechanism underlying the reproduction alterations associated with diabetes,4,5,6 so the antiapoptotic action of NeuroEPO would be beneficial in these conditions.14,15,16,17 However, apoptosis is a required mechanism to balance cell proliferation during morphogenesis and development; the removal of redundant cells from the internal cell mass of the blastocyst, the development of neural crest cells, as well as the closure of the neural tube and palate, are examples of processes in which apoptosis is involved.36 Therefore, the antiapoptotic action of NeuroEPO could mediate the beneficial effects observed in diabetic rats with the dose of 0,5 mg/kg but could cause embryonic alterations incompatible with life at higher doses.

