










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCentro Agrícola
versão On-line ISSN 0253-5785
Ctro. Agr. vol.43 no.2 Santa Clara jun. 2016
ARTÍCULO DE INVESTIGACIÓN
Patogenicidad de especies fúngicas presentes en los rizomas de malanga (Xanthosoma y Colocasia)
Pathogenicity of fungal species in aroid (Colocasia and Xanthosoma) rhizomes
Amaurys Dávila Martínez1, Lidcay Herrera Isla2, Maryluz Folgueras Montiel1 y Ernesto Espinosa Cuellar1
1- Instituto de Investigaciones de Viandas Tropicales (INIVIT), Apartado 6, Santo Domingo, Villa Clara, Cuba. CP 53 000.
2- Universidad Central “Marta Abreu” de Las Villas (UCLV), Facultad de Ciencias Agropecuarias. Santa Clara, Villa Clara, Cuba. CP 54 830.
E-mail: micologia@inivit.cu
RESUMEN
Dentro de las enfermedades que afectan la malanga se encuentra la pudrición de los rizomas ocasionada por varios hongos fitopatógenos. Estas pudriciones aparecen por lo general en suelos mal drenados, pesados y con alto contenido de materia orgánica. Se manifiestan más en épocas lluviosas, pues se trata de un complejo de hongos habitantes del suelo que se favorecen con la alta humedad. Con el objetivo de evaluar la patogenicidad, así como el sinergismo o antagonismo entre las especies involucradas en este síndrome se realizó la presente investigación. Se confirmó la patogenicidad de las principales especies fúngicas identificadas, destacándose en Xanthosoma a S. rolfsii, F. sulphureum y F. chlamydosporum y en Colocasia a Phoma sp., Diplodia sp. y S. rolfsii. En las inoculaciones combinadas, Rhizoctonia solani manifestó sinergismo con el hongo Phoma sp. en Xanthosoma y F. chlamydosporum en Colocasia; con el resto de las especies el efecto es antagónico. S. rolfsii manifestó sinergismo con todos los hongos en Colocasia excepto con Diplodia sp. y Phoma sp., mientras que en Xanthosoma mostró antagonismo con todas las especies.
Palabras clave: Infecciones, pudriciones secas, rizomas, síntomas.
ABSTRACT
Among the diseases affecting aroids is rhizome rot caused by various pathogen fungi. These rots usually appear in poorly drained heavy soils with high organic matter content. These diseases appear more during the rainy season because it is a fungus complex living in the soil and is favored by high humidity. In order to know the virulence of different pathogens involved in this syndrome, cross- species inoculations were performed. Species of Rhizoctonia solani Kühn and Sclerotiun rolfsii Sacc were used in croos inoculations as they showed a higher percentage of appearance in the analyzed samples. The pathogenicity of the major fungal species was confirmed in Xanthosoma: S. rolfsii, F. sulphureum and F. chlamydosporum and in Colocasia: Phoma sp, Diplodia sp.and S. rolfsii. In the combined inoculations, Rhizoctonia solani showed synergism in the fungus Phoma sp in Xanthosoma and F. chlamydosporum in Colocasia and an antagonistic effect with the rest of the species. S. rolfsii showed synergism with all fungi in Colocasia except with Diplodia sp. and Phoma sp. while in Xanthosoma it showed antagonism with all species.
Key words: Infection, dry rot, rhisomes, symptoms.
INTRODUCCIÓN
Dos Aráceas alcanzan importancia mundial como alimentos energéticos: el taro (Colocasia esculenta L. Schott), originaria de Oceanía y sureste de Asia, y la yautía, malanga o quequisque (Xanthosoma sagittifolium L. Schott), de los trópicos americanos. Las enfermedades más importantes de la malanga Colocasia sp. y Xanthosoma sp. son Dasheen mosaic virus (DsMV) (González et al., 2002) y las pudriciones de los rizomas o mal seco (Reyes et al., 2005). Esta última enfermedad, se considera el factor que más ha propiciado la baja producción de cormelos frescos en Puerto Rico. En el presente, las importaciones para cubrir el consumo local de yautía sobrepasan el 97 % (Ortiz, 1997).
El síndrome se caracteriza por pudrición de cormos y cormelos, hojas viejas marchitas y cloróticas. En Puerto Rico se ha asociado a los hongos Pythium sp, Rhizoctonia solani Kühn, Fusarium solani (Mart) Sacc y últimamente a Sclerotium rolfsii Sacc (Morales, 2006).
En Costa Rica ha ocasionado gran pérdida a los productores, obligando a muchos de ellos a abandonar campos de siembra dedicados a este cultivo. La etiología de la enfermedad es incierta y no existe control químico efectivo ni variedades resistentes. Entre los patógenos asociados se encuentran Fusarium sp., Pythium sp., Rhizoctonia sp. y bacterias (Hidalgo et al., 2007).
En la República Dominica esta enfermedad se localiza en casi todas las áreas cultivadas de yautía citándose a Fusarium sp. como el principal organismo causal. Las investigaciones realizadas en las principales áreas de producción muestran que Pythium sp. es también asociado con el mal seco (Perneel, 2006).
Durante los últimos años la aparición de pudriciones secas del cormo y cormelos en la malanga, no han sido estudiadas a profundidad en Cuba (Rodríguez, 2002). Dávila (2011) determinó que los agentes causales de las pudriciones de los rizomas de la malanga son: F. sulfureum, F. solani, F. chlamydosporum, F. oxysporum, S. rolfsii, R. solani, Phoma sp., Diplodia sp. , R. nigricans, informando además otras especies de hongos asociados a esta sintomatología.
Con el objetivo de evaluar la patogenicidad, así como el sinergismo o antagonismo entre las especies involucradas en este síndrome se realizó la presente investigación.
MATERIALES Y MÉTODOS
La investigación se realizó en el Laboratorio de Fitopatología del Instituto de Investigaciones de Viandas Tropicales (INIVIT). La identificación de las especies patógenas se llevó a cabo en el Laboratorio de Microbiología Agrícola de la Facultad de Ciencias Agropecuarias de la Universidad Central “Marta Abreu” de Las Villas.
Para el aislamiento de los hongos se tomaron muestras de plantas que presentaban síntomas de escaso desarrollo vegetativo, clorosis foliar y pudriciones en las raíces. Los rizomas principales y secundarios afectados se lavaron con agua y detergente durante 5 minutos; seguidamente se sumergieron en una solución de Hipoclorito de Sodio (1 %), enjuagándolos con agua corriente durante 5-10 minutos. Los rizomas fueron cortados con la ayuda de un bisturí en fracciones de 5 mm y colocados en placas de Petri de 9 cm de diámetro que contenían Agar Papa Dextrosa (PDA) suplementado con 0,50 mg de cloranfenicol. Se incubaron a 25 oC durante cinco días en oscuridad constante.
Se prepararon suspensiones de conidios de los organismos aislados y se extendieron 0,1 ml de cada una de ellas sobre la superficie de una capa de agar de agua al 2 % y se incubaron a 28 oC durante 24 horas. Posteriormente, se marcaron los conidios germinados con el auxilio del microscopio clínico (Olympus Vanox), con 200 X de aumento. Los conidios germinados se colocaron en medio de cultivo PDA y de cada una de las colonias fungosas obtenidas, se transfirieron porciones de micelio a tubos de ensayo (15x150 mm) que contenían igual medio.
Se seleccionaron los caracteres morfológicos que definen a los géneros y especies de manera objetiva y resultan de fácil observación, tales como: tamaño del conidio (largo y ancho), forma y número de septos de los conidios, tipo de micelio, pigmentación, presencia de clamidosporas, macro y microconidas. También se emplearon claves taxonómicas y manuales especializados para estos fines (Herrera, 1994 y Castañeda, 2001).
En las pruebas de patogenicidad, se utilizaron rizomas principales y secundarios de los clones ‘INIVIT MX 95-1’(género Xanthosoma) y ‘Camerun-14’(género Colocasia) obtenidos por cultivo de tejidos que se encontraban plantados en suelo estéril.
Para la producción del inóculo, las cepas de los hongos aislados se trasfirieron a placas de Petri de 9 cm de diámetro que contenían medio de cultivo PDA para su multiplicación y se incubaron a 28 oC ± 2 oC por un periodo de siete días.
Se realizaron dos inoculaciones por especies de hongos con 15 días de diferencias entre sí. Las especies fúngicas empleadas fueron: Fusarium sulphureum Schlecht, Fusarium chlamydosporum Wollew and Reinking, Fusarium oxysporum Link, Sclerotium rolfsii Sacc, Rhizoctonia solani Kühn, Phoma sp., Rhizopus nigicans Ehr, Fusarium solani (Mart) Sacc, Diplodia sp, Aspergillus niger Tiegh y Penicillum chrysogenum Thom.
En ambas inoculaciones, los rizomas utilizados fueron lavados. Posteriormente se sumergieron en alcohol al 40 % y luego se enjuagaron con agua corriente durante 10 minutos. Con un perforador de tapones de 5 mm de diámetro se obtuvieron discos de micelio de los diferentes hongos objeto de estudio. Se hizo una perforación en el extremo apical de los rizomas principales y secundarios y se introdujo cada disco a 1,0 cm de profundidad cubriendo el mismo con el fragmento del tejido extraído. Se emplearon como controles, discos de medio sin la presencia del hongo. Se colocaron los materiales inoculados en bandejas metálicas, a temperatura de 27 oC ± 2 oC y oscuridad, durante 15 días. Los daños ocasionados por cada uno de los patógenos se evaluaron realizando un corte longitudinal de los rizomas donde se midió el largo y el ancho (en centímetros) de las lesiones a partir del punto de inoculación. Se evaluó el posible antagonismo o sinergismo entre las especies.
Para evaluar el sinergismo o antagonismo entre las especies se realizaron inoculaciones cruzadas, donde se emplearon dos combinaciones. En la primera se combinó a R. solani con todas las especies patógenas y en la segunda se combinó a S. rolfsii con las mismas especies, siguiendo el procedimiento descrito anteriormente (Tabla). Se emplearon estas dos especies como patrones por presentar mayor porcentaje de aparición en las muestras analizadas. Se consideró antagonismo cuando el tamaño de la lesión, en el rizoma, al combinar las dos especies patógenas, era menor que cuando se inoculó cada una de ellas por separado, mientras que el caso del sinergismo ocurre lo contrario.
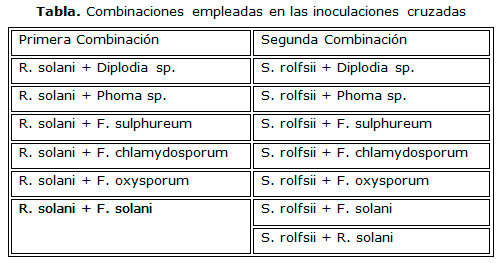
Los datos, correspondientes a tres réplicas por tratamiento, se procesaron con el empleo del paquete estadístico SPSS versión 13.0, mediante análisis de varianza de clasificación simple y la comparación múltiple de medias según la dócima de Dunnett´C, posteriores a la determinación de los supuestos de homogeneidad de varianza y normalidad (Portal Estadística, 2011).
RESULTADOS Y DISCUSIÓN
La especie que ocasionó los mayores daños en los rizomas inoculados fue S. rolfsii, con diferencias estadísticas significativas respecto a las demás especies en estudio (Figura 1). Las especies R. nigricans, A. niger y P. chrysogenum no mostraron diferencias estadísticas con el control. En Nigeria, Igbokwe et al. (1984) informan una alta incidencia de S.rolfsii en malanga, aunque este hongo no se asoció a pudriciones severas del cultivo en el campo y sólo se consideró importante a nivel de la postcosecha. Estos resultados coinciden con los expresados por Mayea (1978) quien determinó como agentes causantes de la pudrición seca en papa las especies F. solani, F. sulphureum y F. oxysporum, mostrando mayor patogenicidad estas dos últimas.
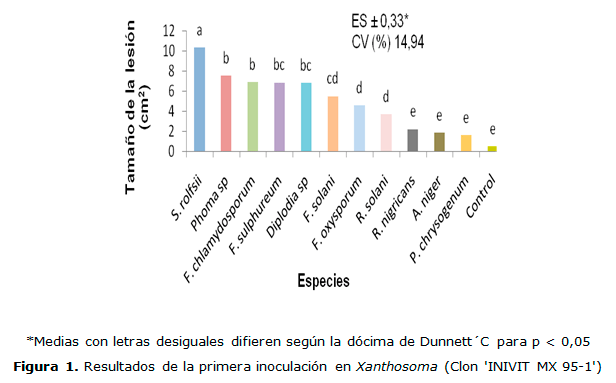
En la segunda inoculación los mayores niveles de infección fueron producidos por S. rolfsii, sin diferencias estadísticas con F. sulphureum, estos resultados están en correspondencia con los informados por Eze y Ameh (2011) al evaluar la patogenicidad de hongos causantes de pudriciones durante el almacenamiento de la malanga Colocasia esculenta. Las especies de A. niger y P. chrysogenum, prácticamente no ocasionaron daños (Figura 2). Bejerano (1998) demostró que en plantas de malanga inoculadas con estos patógenos, se producen síntomas similares y se afectó además, el número de brotes, el diámetro de las raíces, la altura de la planta y el peso seco de los rizomas.

En Colocasia, los niveles de infección superiores estuvieron asociados a Phoma sp., quien presentó valores con diferencias estadísticas significativas respecto al resto de los hongos en estudio (Figura 3).

Las especies S. rolfssi, Diplodia sp. y F. Sulphureum, causaron lesiones semejantes sin diferir estadísticamente, siguiéndolo en orden los hongos F. solani, F. chlamydosporum y F. oxysporum, los cuales no presentaron diferencias estadísticas entre ellos. El resto de las especies inoculadas no manifestaron diferencias con el control. En este estudio se confirmó lo planteado por Smith et al. (1997) y CABI (2000) quienes informan a Phoma exigua var. Foveata, como uno de los principales patógenos en papa (Solanum tuberosum L.), aunque se le ha encontrado en malezas que crecen en campos de papa y cítricos.
En la segunda inoculación al género Colocasia (Figura 4), las especies de Diplodia sp. y F. solani causaron las mayores lesiones con diferencias estadísticas entre ellas y con el resto de las especies inoculadas. Las especies Phoma sp., S. rolfsii y F. sulfureum produjeron lesiones similares sin mostrar diferencias estadísticas significativas entre ellas, seguidas de R. solani que presento valores que difieren estadísticamente de las anteriores. Los resultados expresados por el resto de las especies inoculadas no presentaron diferencias estadísticas respecto al control. Resultados similares de la patogenicidad de Diplodia sp. fueron informados en el cultivo de la yuca por varios investigadores (Lozano y Nolt, 1994; Folgueras, 2010).
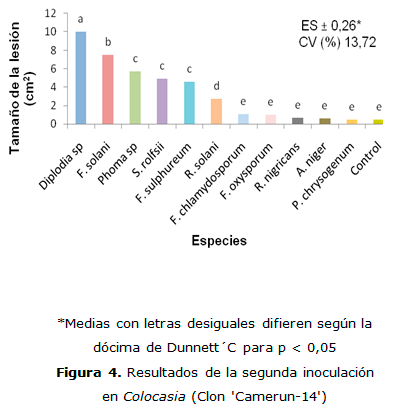
Estos resultados corroboran lo expresado por Morales (2006) quien informa a F. solani entre los principales agentes causales de las pudriciones de los rizomas en el cultivo de la malanga, en Puerto Rico.
La respuesta de los clones ‘INIVIT MX 95-1’ y ‘Camerun-14’ a la primera combinación de inoculaciones cruzadas con las especies fúngicas más frecuentes aparece en la (Figura 5), donde se aprecia que todos los hongos inoculados ocasionaron mayores lesiones que R. solani, siendo mayores los tamaños de las lesiones en el género Xanthosoma, excepto al inocular Phoma sp. en Colocasia (Figura 5B).
Las lesiones provocadas por las combinaciones de R. solani con cada una de las especies fitopatógenas evaluadas fueron mayores siempre en el clon ‘INIVIT MX 95-1’. Al combinar R. solani + Phoma sp. (Figura 5B) se produjeron mayores lesiones en Xanthosoma. Las combinaciones R. solani + F. sulphureum (Figura 5C), R. solani + F. chlamydosporum (Figura 5D) y R. solani + F. solani (Figura 5F), originaron lesiones mayores que cuando R. solani se inoculó sólo.
En el clon ‘Camerun-14’ resultaron mayores las lesiones con la combinación R. solani + F. chlamydosporum (Figura 5D). El resto de las combinaciones ocasionaron lesiones de tamaños mayores a la provocada por R. solani, excepto en las combinaciones R. solani + Diplodia sp. (Figura 5A) y R. solani + F. oxysporum (Figura 5E).
Los resultados obtenidos en cuanto al antagonismo expresado por R. solani con todas las especies de hongos inoculadas excepto con Phoma sp en Xanthosoma y F. chlamydosporum en Colocasia son similares a los informados por Zink y Secor (1982), al realizar inoculaciones combinadas de la bacteria Erwinia carotovora subsp. atroseptica con F. sulphureum, F. solani y F. oxysporum. Estos autores determinaron que el incremento en el porcentaje de los tallos con “pata prieta” no fue significativamente diferente con respecto a las especies de hongos inoculadas, lo que indica que tuvo lugar un efecto antagonista en vez de una relación sinergística específica entre los hongos y la bacteria.
La (Figura 6) se muestra la respuesta de los clones ‘INIVIT MX 95-1’ y ‘Camerun-14’ a la segunda combinación de inoculaciones cruzadas. Siempre los hongos fitopatógenos estudiados provocaron mayores lesiones en el género Xanthosoma, excepto Phoma sp. que produjo lesiones de 9,82 cm2 en Colocasia (Figura 6G).
Todas las combinaciones efectuadas en el clon ‘INIVIT MX 95-1’ causaron lesiones de menor tamaño que las provocadas por S. rolfsii. En el clon ‘Camerun-14’, al combinar las dos especies, se originaron lesiones mayores que cuando se inocularon cada una de ellas individualmente, a excepción de Diplodia sp. (Figura 6B) y Phoma sp (Figura 6G). Estos resultados obtenidos en Colocasia corroboran los estudios realizados por Espinosa (2003) quien determinó que la combinación de S. rofsii, R. solani y F. oxysporum provocaba mayores afectaciones en los rizomas que cuando cada especie se inoculaba de forma aislada. Nishimura y Kudo (1994) en inoculaciones realizadas sobre 13 variedades de Taro (Colocasia) en Japón confirmaron la patogenicidad de F. oxysporum y F. solani como causantes de la podredumbre seca en el cultivo.
Este ensayo constituye una prueba preliminar para evaluar la respuesta de los diferentes patógenos inoculados por lo que se requiere de un estudio con un diseño experimental y un tratamiento estadístico para confirmar estos resultados.
CONCLUSIONES
1. Se confirmó la patogenicidad de las principales especies fúngicas identificadas, destacándose en Xanthosoma S. rolfsii, F. sulphureum y F. chlamydosporum y en Colocasia Phoma sp, Diplodia sp. y S. rolfsii.
2. En las inoculaciones combinadas, Rhizoctonia solani manifestó sinergismo con el hongo Phoma sp en Xanthosoma y F. chlamydosporum en Colocasia y con el resto de las especies un efecto antagónico. S. rolfsii manifestó sinergismo con todos los hongos en Colocasia excepto con Diplodia sp. y Phoma sp. mientras que en Xanthosoma mostró antagonismo con todas las especies.
3. Los patógenos inoculados provocaron mayores afectaciones en el género Colocasia que en Xanthosoma.
BIBLIOGRAFÍA
1. Bejerano, C. A.; A.M. Zapata; E. Bosques; I.L. Rivera: Sclerotium rolfsii como componente del complejo patológico causante del mal seco de la yautía (Xanthosoma sagittifolium): Seminario Taller sobre Producción de aráceas, ñame, otoe y yuca. Puerto Rico. 1998, 53 p.
2. CABI: Data sheet for Phoma exigua var, foveata. Crop Protection Compendium. Global Module. 2nd. Edition. CAB International, Carnegie, UK, Pp. 57-68, 2000.
3. Castañeda, R. F.: Identificación de hifomicetes causantes de enfermedades en hortalizas comunes en Cuba. Tesis presentada en opción del grado científico de Doctor en Ciencias Agrícolas. INIFAT, Ciudad Habana, Cuba. 2001, 100 p.
4. Dávila, A.: Las pudriciones secas de la malanga (Xanthosoma y Colocasia). Etiología y sintomatología. Tesis de Maestría. Facultad de Ciencias Agropecuarias, Universidad Central “Marta Abreu” de Las Villas, Villa Clara, Cuba. 2011, 54 p.
5. Espinosa, E.: Estudio de las pudriciones secas en el cultivo de la malanga (Xanthosoma spp.) y Colocasia esculenta Schott. Tesis de Maestría. Facultad de Ciencias Agropecuarias, Universidad Central “Marta Abreu” de Las Villas, Villa Clara, Cuba. 2003, 45 p.
6. Eze, C.S. and G.I. Ameh: Comparative Assessment of Pathogenicity of Storage Rot Causing Fungi of Cocoyams Colocasia esculenta (L.) Schott and their Host. Pathogen Interactions, 22: 23-27, 2011.
7. Folgueras, M.: Las pudriciones radiculares de la yuca (Manihot esculenta Crantz) en Cuba: Tesis presentada en opción del grado científico de Doctor en Ciencias Agrícolas. Universidad Central “Marta Abreu” de Las Villas, Santa Clara, Cuba. 2010, 98 p.
8. González, M. Y.; R. Hernández; J.E. González; Y. Beovides; D. Horta: Método eficiente para el diagnóstico del DMV en malanga. Trabajo presentado en evento “35 Aniversario de INIVIT”. Santa Clara. Cuba, octubre, 2002, 17 p.
9. Herrera, L. y S. Mayea: Fitopatología General. Editorial Félix Varela. Ciudad de la Habana, Cuba. 1994, 316 p.
10. Hidalgo, N.; F. Saborio; l. Gómez; S. Torres: Estudio preliminar de patógenos relacionados con el mal seco en quequisque blanco (Xanthosoma sagittifolium). Boletín No. 4 del Centro de Investigaciones en Protección de Cultivos, Universidad de Costa Rica, San Pedro de Montes de Oca, Costa Rica. Pp. 12-14, 2007.
11. Igbokwe, M.C.; D.B. Arene; T.C. Naubuizu; E.E. Umanda: Intercropping cocoyams with plantain: Efects on the yield and disease cocoyams. In Tropical Root Crops: Production and uses In Africa. pp 182-184.1984.
12. Lozano, J. C. y B. Nolt: Diseases of Cassava (Manihot esculenta Crantz) Phytopathology. The American Phytopathological Society. St. Paul, Minnesota, EE.UU. 1994, 344 p.
13. Mayea, S.; D. Seidel; J. Gotz: Especies de Fusarium en los tubérculos de papa en Cuba, en Tag. Ver. Akacl. Lanwirstsch-Wiss. RDA. 157: 67-70, 1978.
14. Morales, A.: La enfermedad del mal seco en tiquizque (Xanthosoma sagitifolium). Ministerio de Agricultura y Ganadería. Tesis de Grado. Dirección regional Brunca, Costa Rica. 2006, 45 p.
15. Nishimura, N. and K. Kudo: Fusarium oxysporum f. sp. Colocasiae n. f. sp. Causing Dry Rot of Taro (Colocasia esculenta). Ann. Phytopathology Soc. Japan 60 (4): 448-453, 1994.
16. Ortiz, L. J.: Situación y perspectiva económica de la Empresa de Raíces y Tubérculos. Informe económico, USDA- ARSTARS, Mayagüez, Puerto Rico. 1997, 35 p.
17. Perneel, M.: The root rot pathogen Pythium myriotylum on cocoyam (Xanthosoma sagittifolium (L.) Schott): intraspecific variability and biological control. PhD thesis. Ghent University, Belgium. 2006, 178 p.
18. Reyes, G.; M. Nyman; A. C. Ronnberg – Wastljung: Agronomic performance of three cocoyam (Xanthosoma violaceum Schott) genotypes grown in Nicaragua. Euphytica 142: 265 – 272, 2005.
19. Rodríguez, S.: Informe sobre la producción de las raíces y tubérculos. Reunión del Grupo Nacional de Viandas. Santo Domingo, Villa Clara, Cuba. 2002, 23 p.
20. Smith, I. M.; D. G. Mcnamara; P. R. Scott; M. Holderness: Phoma exigua var, foveata. Data Sheets on Quarantine Pest. Pp. 865-871. In: Quarantine Pest for Europe. Second Edition. CABI International & EPPO, Carnegie, UK. 1997, 1425 p.
21. Zink, R.T. and G. A. Sencor: Interaction of fungal wilt pathogens and potato blackleg. Plant Disease 66: 1053 – 1056, 1982.
Recibido: 13/01/2015
Aceptado: 03/02/2016


{kind=link}
{kind=link}
{kind=link}