










 Serviços customizados
Serviços customizados Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El cacao tradicional, es un recurso genético local con características morfológicas y genéticas poco conocidas. Su presencia en zonas aisladas con pocas atenciones culturales, indican que puede ser resistente a factores bióticos y abióticos del ambiente y las semillas blancas de algunas plantas son muy apreciadas en el mercado internacional. Estas son potencialidades para el mejoramiento genético de la calidad y la adaptación a las condiciones de Cuba. Las potencialidades genéticas del cultivo pueden constituir una vía para combatir la podredumbre oscura de la mazorca, pero se requiere conocer las especies que causan esta enfermedad.
Basados en lo anterior nos planteamos los objetivos: Caracterizar la variabilidad morfológica y genética del cacao tradicional cubano para clasificarlo en los grupos tradicionales y genéticos del cultivo, Caracterizar morfológica y molecularmente las especies de Phytophthora, causantes de la podredumbre oscura de la mazorca en Cuba.
MÉTODOS
Análisis morfológico del cacao tradicional cubano
En 537 plantas de cacao tradicional cubano y 105 plantas de referencia se calculó la frecuencia de 33 descriptores morfológicos cualitativos 1-3 y se clasificaron en los grupos tradicionales mediante un análisis de conglomerados con la varianza mínima de Ward.
Análisis genético mediante microsatélites
El ADN genómico de las plantas de cacao se extrajo y amplificó con los 15 pares de cebadores microsatélites 4,5 según se describió previamente. 6 Se calculó la frecuencia y el número de alelos, alelos raros, alelos privados, 7 intervalo de tamaño de los alelos, heterocigosidad observada y esperada, contenido de información polimórfica, 8 índice de Garza-Williamson9 y la desviación del equilibrio de Hardy-Weinberg.10
Con el programa structure versión 2.3.4 11 se determinó la estructura poblacional del cacao tradicional cubano y se clasificó en los grupos genéticos al analizarlo con plantas de referencia, 12 según los parámetros previamente descritos. 6
Selección de las plantas que integrarán la colección núcleo
Se seleccionó la colección núcleo de cacao tradicional cubano con el algoritmo de máxima diversidad genética 13,14 implementado en el lenguaje de programación estadística R versión 3.01. (15
Caracterización morfológica de las cepas de Phytophthora
Se tomaron 90 aislados de Phytophthora de frutos enfermos y se mantuvieron en medio agar V8 a 24-29 °C. Las características morfológicas se observaron al microscopio óptico Olympus BX50 y se fotografiaron con el programa Ulead Photo explorer SE versión 7.0.
Análisis filogenéticos de las cepas de Phytophthora
El ADN se extrajo de micelio fresco crecido en extracto de malta líquido a 25 °C en la oscuridad y se amplificó la β-tubulina, TEF1-α e ITS con el par de cebadores TUBUR1-TUBUF2, 16 ELONGF1-ELONGR1 16,17 y NS7-ITS4, 18 respectivamente, según se describió previamente. 19
El análisis filogenético se realizó por el método de máxima parsimonia de PAUP* versión 4.0b10. 20 Se identificó el árbol de mayor parsimonia con búsquedas heurísticas con adición al azar de secuencia (1000), un conjunto máximo de árboles de 100, evaluado posteriormente mediante bootstrap, se retuvieron los clados compatibles con la regla de mayoría del 50 % en el árbol bootstrap consenso.
Los ITS de las secuencias de β-tubulin y TEF1-α se compararon con la base de datos de Genbank.
RESULTADOS Y DISCUSIÓN
Análisis morfológico del cacao tradicional cubano
Se observaron 98 de las 104 clases fenotípicas de los 33 descriptores morfológicos evaluados, lo que indica una alta variabilidad morfológica del cacao tradicional cubano. Las clases fenotípicas más representativas, con frecuencia superior a 0,5, fueron:
Del brote de la hoja: ausencia de verde.
En la flor: flor cerrada blanca, pedúnculo violeta claro, sépalos separados, con orientación horizontal y blanco, pétalos con limbo pendiente, amarillo intenso y con ausencia de rojo, filamento mayor que la concha y el limbo, presencia de líneas de guía, estaminoides divergentes y violeta intenso y filamentos de los estambres blancos.
Del fruto: LA relación largo por ancho mayor que dos, forma general angoleta, superficie rugosa en grado intermedio, profundidad del surco mediana, presencia de cinco grupos de dos lomos cada uno, fruto maduro amarillo, ausencia de antocianinas en el lomo y el surco del fruto inmaduro y en el lomo y el surco del fruto maduro y mesocarpio duro.
En la semilla: pulpa blanca, sabor levemente ácido, sección transversal aplastada y violeta intenso.
La variabilidad morfológica de las plantas de cacao tradicional cubano fue similar o superior a los clones del proyecto CFC/ICCO/IPGRI, 21 accesiones del banco de germoplasma internacional de cacao, Trinidad, 22 fincas y el banco de germoplasma de Camerún. 23
Las 68 plantas con semillas de cotiledón blanco son de interés para elaborar un chocolate fino y de excelente calidad, 24,25 con bajas concentraciones de antocianinas y flavonoides 26 y altas concentraciones de teobromina. 27
La caracterización morfológica por primera vez del cacao tradicional cubano es de gran importancia, más cuando se estudia fundamentalmente la variabilidad genética, y hay pocos estudios de variabilidad morfológica. 28,29
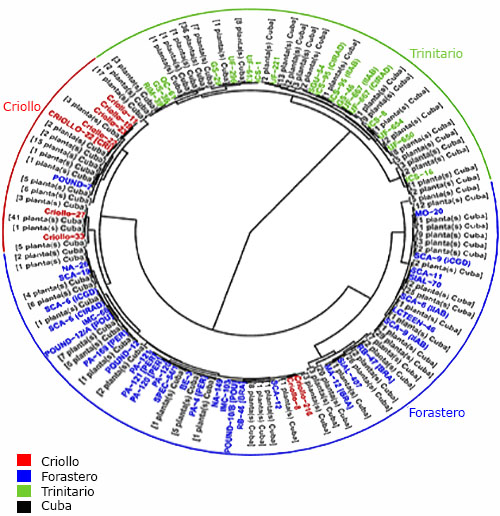
En el dendrograma se clasificaron 111 plantas de cacao tradicional cubano como trinitarias, 212 como forasteras y 102 como criollas, resultado relacionado con la variabilidad morfológica de los grupos tradicionales Trinitario y Forastero 30 (Fig. 1).
Las semillas blancas son típicas del grupo genético Criollo, pero el cacao tradicional cubano con esta característica no se clasificó como Criollo. Sin embargo, los cultivares tradicionales finos de aroma, con antecedentes Criollo y Forastero producen un chocolate fino 31 para mercados especializados, 31,32 con altos precios. 33,34
Análisis genético mediante microsatélites
La suma de las frecuencias de dos alelos en todos los microsatélites fue superior a 0,8. Esta característica es típica de plantas trinitarias, 35,36 por las hibridaciones e introgresiones en Trinidad de Criollo antiguo y Amelonado homocigóticas, 37,38 lo que indica que el cacao tradicional cubano es Trinitario.
Se observaron 139 alelos por microsatélite, con media de 9,267 alelos por locus. Los alelos raros fueron 104, con una media por locus de 6,933 (Tabla 1).
Tabla 1 Variabilidad genética de los 15 microsatélites evaluados en el cacao tradicional cubano. Elaboración propia
| Microsatélite | Alelos | Alelos raros | Alelos privados | Intervalo de tamaño (pb) | Ho | He | Equilibrio de |
PIC | Índice de Garza- |
| mTcCIR1 | 6 | 4 | 2 | 129-141 | 0,404 | 0,418 | 0,031 | 0,337 | 0,500 |
| mTcCIR6 | 9 | 7 | 2 | 227-251 | 0,335 | 0,363 | 0,369 | 0,341 | 0,375 |
| mTcCIR7 | 6 | 3 | 1 | 149-161 | 0,238 | 0,300 | 0,076 | 0,279 | 0,500 |
| mTcCIR8 | 11 | 9 | 4 | 284-306 | 0,270 | 0,292 | 0,096 | 0,263 | 0,500 |
| mTcCIR11 | 8 | 6 | 0 | 290-316 | 0,364 | 0,399 | 0,000 | 0,361 | 0,346 |
| mTcCIR12 | 10 | 8 | 2 | 177-252 | 0,440 | 0,425 | 1,000 | 0,375 | 0,133 |
| mTcCIR15 | 10 | 8 | 1 | 234-258 | 0,449 | 0,526 | 0,003 | 0,463 | 0,417 |
| mTcCIR18 | 7 | 5 | 0 | 332-356 | 0,387 | 0,441 | 0,015 | 0,392 | 0,292 |
| mTcCIR22 | 9 | 7 | 4 | 275-296 | 0,318 | 0,376 | 0,017 | 0,330 | 0,429 |
| mTcCIR24 | 6 | 4 | 0 | 186-202 | 0,181 | 0,182 | 1,000 | 0,169 | 0,375 |
| mTcCIR26 | 7 | 5 | 2 | 285-305 | 0,248 | 0,346 | 0,003 | 0,319 | 0,400 |
| mTcCIR33 | 17 | 14 | 5 | 273-345 | 0,380 | 0,486 | 0,001 | 0,467 | 0,250 |
| mTcCIR37 | 12 | 9 | 0 | 135-175 | 0,394 | 0,495 | 0,045 | 0,470 | 0,325 |
| mTcCIR40 | 11 | 8 | 1 | 262-286 | 0,481 | 0,644 | 0,283 | 0,578 | 0,500 |
| mTcCIR60 | 10 | 7 | 2 | 188-214 | 0,660 | 0,597 | 0,194 | 0,535 | 0,423 |
| Total | 139 | 104 | 26 | - | - | - | - | - | - |
| Media | 9,267 | 6,933 | 1,733 | - | 0,370 | 0,419 | - | 0,379 | 0,384 |
| Desv. est. | 2,816 | 2,620 | 1,526 | - | 0,113 | 0,116 | - | 0,105 | 0,101 |
pb: pares de bases, Ho: Heterocigosidad observada, He: Heterocigosidad esperada, PIC: Contenido de Información Polimórfica, Desv. est. Desviación estándar
El número de alelos del cacao tradicional cubano fue superior al observado en Bolivia, 39 Ecuador, 40 Nicaragua 33 y República Dominicana 41 e inferior a bancos de germoplasma como la colección alto amazónica, 42 África Occidental 43 y CATIE 44. Además, los alelos privados y raros indican información genética típica o exclusiva, potencialmente útil para el mejoramiento genético. 45
La heterocigosidad observada varió entre 0,181 y 0,660 y la esperada entre 0,182 y 0,644. Estos resultados fueron superiores a plantaciones de Criollo en México (0,28), 46 pero inferiores a plantaciones de la costa ecuatoriana, República Dominicana y bancos de germoplasma en Costa Rica, Perú, Puerto Rico y Ghana (0,460-0,717). 47-50
La media de la heterocigosidad esperada en el cacao tradicional cubano fue inferior a la observada en plantaciones de la costa ecuatoriana, Amazonas, Perú, República Dominicana, Nicaragua y Criollo en México (0,476 y 0,74). 33,41,46,51-53
Ocho de los microsatélites no estaban en equilibrio de Hardy-Weinberg, con déficit de heterocigotos. Resultados similares se observaron en cacao Criollo en México, 46 Refractario en Ecuador, 40 en el Amazonas brasileño, 52 Ecuador 40 y el cacao Nacional boliviano. 53
El bajo valor de la heterocigosidad observada, esperada y el déficit de heterocigóticos indican baja variabilidad genética, debida a la alta frecuencia de dos alelos por locus y la endogamia, favorecida por sus poblaciones reducidas y aisladas. 54,55
El Índice de Garza-Williamson fue inferior a 0,68; límite por debajo del cual se considera una reducción reciente en el tamaño poblacional. 9 Este resultado confirma la historia del cultivo en Cuba, introducido por Cabaiguán (1540) y expandido por la zona central que concentró el 90 % del cacao durante el siglo XVIII. Posteriormente (1791-1803) se introdujo por Ti Arriba desde Haití, se expandió por la región oriental y disminuyó en el centro. 56
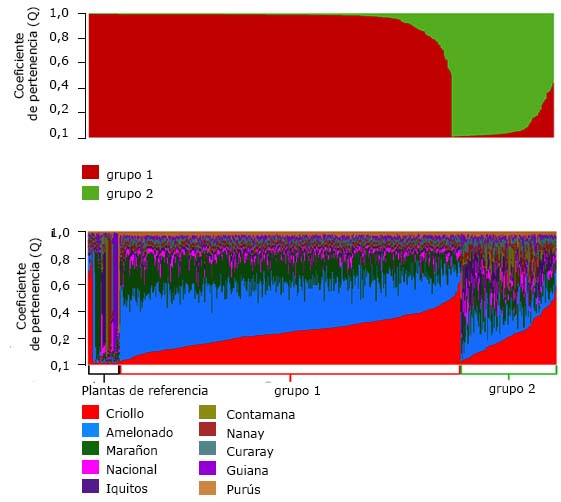
Se observaron dos grupos en el cacao tradicional cubano. La media del coeficiente de pertenencia de cada planta a su grupo fue de 0,952. En el grupo 1 se observaron 419 plantas y 118 en el grupo 2. (Fig. 2a).
Los grupos genéticos más frecuentes del cacao tradicional cubano fueron Amelonado (61,64 %) y Criollo (27,37 %). Las plantas restantes se clasificaron en los grupos Marañón (5,40 %), Iquitos (2,23 %), Contamana (1,49 %), Nanay (1,12 %) y Nacional (0,75 %) (Fig. 2b).
En el grupo 1 (asociado con la región oriental), el 96,90 % de las plantas se clasificaron en los grupos genéticos Amelonado y Criollo, lo que indique que son Trinitarias. Esta puede ser una evidencia de la introducción a Cuba en 1791-1803, cuando se cultivaba el Forastero y Trinitario. El 3,10 % restante pertenece al grupo Marañón.
En el grupo 2 (asociado con la región central) las plantas se clasificaron en siete grupos genéticos: Criollo (35,59 %), Amelonado (25,42 %), Marañón (13,56 %), Iquitos (10,17 %), Contamana (6,78 %), Nanay (5,09 %) y Nacional (3,39 %), lo que indica una mayor variabilidad genética. El 35,59 % de las plantas del grupo 2 se clasificaron en el grupo Criollo, evidencia de la introducción en 1540, cuando se cultivaba cacao Criollo. 38,56
Los grupos genéticos del cacao tradicional cubano coincidieron con los de República Dominicana: Amelonado (61,64 % y 72,10 %), Criollo (27,37 % y 9,50 %), y los grupos Marañón, Iquitos, Contamana, Nanay y Nacional con menos del 20 %. 41 Como el origen del cacao en República Dominicana y Haití es el mismo, es evidencia de la introducción a Cuba desde Haití. 38
Selección de las plantas que integrarán la colección núcleo
Se seleccionaron 185 plantas para la colección núcleo, que se está constituyendo en Baracoa. Esta colección núcleo se utilizará en el mejoramiento genético del cacao, en especial para incrementar la calidad. Las colecciones núcleo disminuyen los costos de conservación y facilitan el mejoramiento genético. 57,58
Caracterización morfológica de las cepas de Phytophthora
En el medio V8, 88 cepas cubanas presentaron un patrón de crecimiento petaloide. La producción de esporangios fue abundante, con formas ovoides a elipsoidal, pedicelio ≤ 4 µm de longitud, 38 µm-60 µm de longitud y 27 µm-40 µm de ancho. El radio L/A (largo/ancho) varió de 1,25 a 1,92. Se observaron las clamidosporas, con diámetro de 32,5 µm -43,5 µm. Estas características morfológicas corresponden a las observadas en Phytophthora palmivora.59,60
En dos aislados cubanos (CU58 y PHYTO40) y en los aislados de la Guyana Francesa, los esporangios midieron 37 µm-50 µm de longitud y 22 µm -32 µm de ancho, con un radio L/A de 1,8 a 2,4. Se observó una larga hifa de 50 µm. Según estas características las cepas pertenecen al complejo Phytophthora capsici-Phytophthora tropicalis61.
Análisis filogenéticos de las cepas de Phytophthora
Se identificaron dos especies de Phytophthora en Cuba: P. palmivora y P. tropicalis, dos en la Guyana Francesa: P. tropicalis / capsici complex y una potencial nueva Phytophthora sp. En Cuba P. palmivora fue la especie dominante y en la Guyana francesa fue P. tropicalis / capsici complex.
Las cepas de P. palmivora tuvieron un alto grado de similaridad (Figura 3).
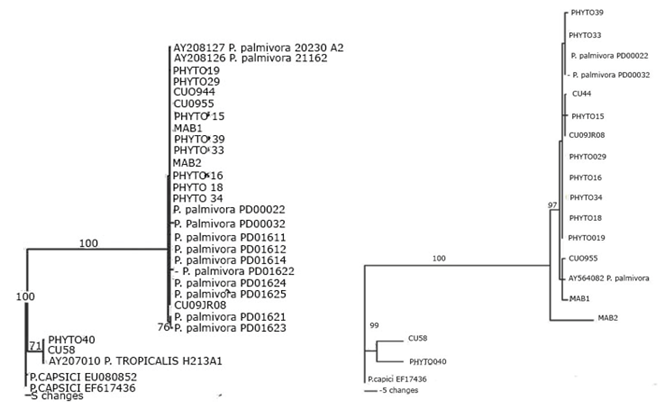 Elaboración propia.
Elaboración propia.Fig. 3 Árbol filogenético de las cepas cubanas de Phytophthora inferido por análisis de máxima parsimonia. a. basado en las secuencias de la región ITS, b. basado en las secuencias del factor de elongación 1-α.
En los árboles filogenéticos de la región ITS y el gen TEF1- α se observó un máximo de 5 bp entre los 700 pares de bases de longitud de la región ITS, lo que corresponde a una proporción de sitios variables de alrededor de 0,01. La región TEF1-α de 1000 pares de bases de longitud fue más variable, con un máximo de 20 bp diferentes y una proporción de sitios variables de 0,02 bp.
Para el complejo Phytophthora tropicalis / capsici se contó con 865 bp, de los cuales 67 fueron informativos para la parsimonia. El análisis resultó en 981 árboles más parsimoniosos (MPT) (140 steps in length, CI = 0,771; RI = 0,904), de las 1000 búsquedas heurísticas. Se seleccionaron dos topologías concurrentes para las cepas de la Guyana Francesa (Fig. 4) que forman un clado bien soportado.
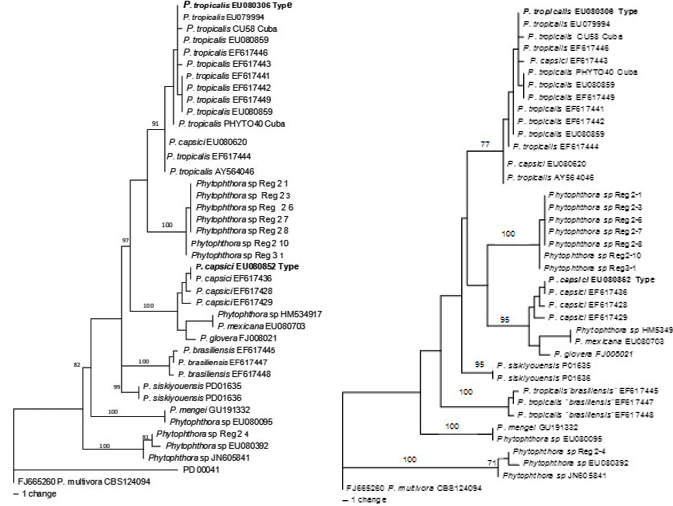 Elaboración propia.
Elaboración propia. Fig. 4 Árbol filogenético que muestra las cepas cubanas de Phytophthora basado en las secuencias del gen β-tubulina, inferido por análisis de máxima parsimonia. a. Topología más frecuente. b. Topología menos frecuente.
En los árboles filogenéticos y en todas las secuencias utilizadas (ITS, β-tubulina), los aislados de Cuba (CU58, PHYTO40) tentativamente identificados como complejo P. tropicalis / capsici se agruparon en el clado P. tropicalis s.s. Este clado contiene a la cepa tipo de P. tropicalis (CBS434.91), cepas de varios orígenes y varios hospederos. Las cepas cubanas difieren de P. tropicalis (type) en 4 bp a 5 bp, 6 bp a 7 bp, 3 bp a 5 bp, en ITS y β-tubulina, respectivamente.
Los aislados de la Guyana Francesa tentativamente identificados como P. capsici62 se ubicaron en dos diferentes linajes según sus diferencias morfológicas.
El primer clado comprende 7 cepas mientras el segundo clado contiene una cepa (Reg 2-4). El linaje Reg 2-4 fue en todas las filogenias, distantemente relacionado con P. tropicalis y P. capsici. Las otras 7 cepas forman un clado bien soportado en los tres análisis filogenéticos (linaje Guyana).
En las inferencias filogenéticas basadas en β-tubulina, el linaje de Guyana formó un clado bien soportado (Bootstrap 100) que se ubicó en una posición aislada, pero sus afinidades no están resueltas. Se observaron topologías concurrentes del análisis individual de cada gen. En su topología dominante (Figura 4a), el clado de Guyana se asoció al clado de P. tropicalis (type). En la topología menos frecuente (Figura 4b), este clado se asoció a P. capsici.
Conclusiones
La alta variabilidad morfológica y la baja variabilidad genética del cacao tradicional cubano es el resultado de su evolución a partir de plantas introducidas en Cuba con antecedentes de siete grupos genéticos y sus híbridos, con una estrecha base genética como resultado de un cuello de botella.
La clasificación del cacao tradicional cubano en el grupo Trinitario basado en su variabilidad morfológica y genética, le atribuye potencialidades para su empleo en el mejoramiento genético del cacao comercial presente en Cuba.
El algoritmo de máxima diversidad genética permitió seleccionar las plantas más representativas de la variabilidad morfológica y genética del cacao tradicional cubano, las que se proponen como colección núcleo para su conservación ex situ.
Dos especies de Phytophthora afectan los frutos de cacao en Cuba: Phytophthora palmivora, predominante en las muestras, y Phytophthora tropicalis, la cual se observó en una frecuencia muy baja.

