










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Cubana de Ciencias Forestales
versão On-line ISSN 2310-3469
Rev CFORES vol.11 no.2 Pinar del Río maio.-ago. 2023 Epub 04-Ago-2023
Original article
Edaphic macroinvertebrates in productive systems of Coffea arabica in Garzón, Huila, Colombia
1Universidad de la Amazonia, Facultad de Ingeniería. Florencia, Colombia.
Agroforestry systems are an increasingly implemented alternative for coffee production in Colombia due to the multiple socioeconomic and environmental benefits. They generate an ideal habitat for soil macrofauna. The objective of this research was to evaluate the incidence of two coffee production systems on the population behavior of edaphic macroinvertebrates. In order to do this, the composition, richness, abundance and vertical distribution of soil macroinvertebrate families in Secondary Forest (Bs), Coffee Agroforestry Systems (SAFc) and Coffee Monoculture (Mc) systems were evaluated in four strata (leaf litter, 0-10 cm, 10-20 cm and 20-30 cm). 22 orders and 53 families of edaphic macroinvertebrates were identified, presenting the highest abundance of Lumbricidae, Formicidae, Porcellionidae and the lowest of Forficulidae, Geophilidae and Polydesmidae, with Bs > SAFc > Mc, except Forficulidae, Polydesmidae and Porcellionidae, which were found in greater abundance in Mc than in SAFc; in all systems the abundance was higher in the first 10 cm of the soil (p<0.05). Wealth showed a similar behavior, although without statistically significant effects. Although there are soil macroinvertebrates that are tolerant to the disturbances caused by forest cover changes to establish coffee production systems, some families of macroinvertebrates are susceptible to these cover changes, which makes them potential indicators of soil disturbances.
Key words: abundance; coffee; edaphic fauna; richness; agroforestry system.
INTRODUCTION
Coffee (Coffea arabica L.) is an example of the agricultural crops that have been developed in Colombia since the 18th century to the present. Currently, the departments of Huila, Nariño and Cauca make up the new 'Coffee Axis'. The cultivated surfaces have grown 63.6 % in the last 13 years, which leads to a greater expansion of the local agricultural territory, compromising the soil resource. Thanks to its agricultural vocation and coffee tradition, the department of Huila has established itself in recent years as the main coffee producer in Colombia (Cerquera et al., 2020)
The transformation of native landscapes due to the intensification of coffee monocultures has been one of the main causes of the loss of biodiversity in high Andean ecosystems. Edaphic macroinvertebrates, which are soil invertebrates larger than 2 mm, can play an important role as bioindicators. Its abundance, taxonomic richness and vertical distribution vary depending on the vegetation cover, the different uses and management of the soil, as well as seasonal variations (Doblas et al.,2009; Cabrera et al., 2017; Velásquez and Lavelle, 2019)
Agroforestry systems (crop and tree association) represent an opportunity for coffee production with sustainability criteria. These systems are positively correlated with biodiversity Campera et al. (2022) and the abundance of soil macrofauna (Rojas et al., 2021). In contrast, in coffee monocultures bioindicator organisms predominate, that indicates imbalance and disturbance in the soil (Rodríguez et al., 2019).
The objective of this research is to evaluate the incidence of two coffee production systems on the composition, richness, abundance and vertical distribution of edaphic macroinvertebrates in the municipality of Garzón, Huila, Colombia.
MATERIALS AND METHODS
Study zone
It is located 15 km east of the municipality of Garzón - Huila (Colombia). Nine farms with an average area of 1.0 ha were identified. These were located between the coordinates 02º20'52" north latitude and 75º54'17" west longitude of Greenwich, at an altitude of 1,460 to 1,668 masl. All the farms are within the Premontane Humid Forest life zone. (Bh-P) according to the Holdridge classification. Average temperatures between 18 °C and 24 °C predominate and annual rainfall ranges between 2,000 and 3,500 mm. The soils of the area are superficial to deep, well drained, slightly acid and of moderate fertility (IGAC, 1994, IGAC 2014).
Systems evaluated
Coffee monocultures (MC) were evaluated with a planting distance of 1.2 mx 1.8 m and a density of 4,630 trees per hectare, agroforestry systems of coffee and plantain (Musa paradisiaca L.) (SAFC), established at a distance of 1.2 mx 1.8 m and 5.6 mx 3 m, respectively, and areas of secondary forest (BS) moderately intervened as positive control. The coffee plantations corresponded to plantations of the Catimor variety, established 11 years ago, two years after the last stumping (total pruning of the stems 30 centimeters from the ground to stimulate the regrowth of new stems).
The agronomic management of MC and SAFC was carried out by the producers following the recommendations of the technical assistants in the area, thus: applications of 100 cm3 of glyphosate in 20 L of water to control weeds, twice a year and pruning with a scythe every four months. Pest control was carried out through applications of pesticides based on Phenylpyrazole, Chlorpyrifos and Thiamethoxam + Ciproconazole, in doses of 25 ml per 20 L of water, with annual applications in SAFC and biannual in MC. Fertilization was performed with urea, diammonium phosphate (DAP) and calcium oxide (CaO). In each case, a dose of 50 grams per plant was used, every four months.
The secondary forest (Bs) is approximately 40 years old; Ararat and Prager (2002) found in these forests in the area, species such as Goupia glabra A. (white cedar), Cedrela odorata L. (pink cedar), Erythrina poeppigiana W. (cachingo), Persea caerulea R&P (aguacatillo), Chrysophyllum cainito L. (caimo), Inga marginata W. (guamo) and Jacaranda copaia A. (gualandai).
Establishment of the design and sampling in the field
A Randomized Complete Blocks (BCA) design was established in a factorial arrangement with four (4) replicates. Factor A corresponded to the evaluated systems (Mc, SAFc and Bs) and factor B to the four invertebrate sampling depths (leaf litter, 0-10 cm, 10-20 cm and 20-30 cm).
The samplings were carried out in the month of October 2018 (there was a collection permit, according to Resolution 01140 of 2016), in the rainy season, before starting the second significant coffee harvest of the year to avoid anthropic disturbances.
The edaphic macroinvertebrates were collected using the method suggested by Velasquez and Lavelle, (2019). In each system, three monoliths (25 x 25 cm) or subsamples per replicate were collected; the monoliths were divided into four strata: litter, 0-10, 10-20 and 20-30 cm. For the collection of edaphic macroinvertebrates, they used entomological tweezers and brushes. These were preserved in 70 % alcohol, with the exception of the worms, which were preserved in vials with 5 % formalin. Subsequently, they were transferred to the Entomology Laboratory of the University of the Amazon (LEUA) for their characterization at the order and family level. The taxonomic keys and descriptions of Ruiz et al. (2008) and Borror et al. (1992) were used.
Response variables and analysis of biological data
For the analysis of macroinvertebrates, the values of richness (number of families), specific abundance (individuals m -2), density (number of individuals m -2) and vertical distribution of the macroinvertebrates were determined as a function of abundance in each profile or soil layer. The overall results in each of the systems were calculated and presented.
In addition, the assumptions of normality and homogeneity of the variances of the data were validated by means of an exploratory residual analysis (QQ plot, Shapiro-Wilk test, and plot of predicted values against residuals). The data that presented a normal distribution and homogeneity of variance (richness) were analyzed using Fisher's LSD test (p<0.05) and the data that did not present a normal distribution were analyzed using the non-parametric Kruskall-Waill test. (p<0.05).
A multivariate analysis was used in order to reduce the information and facilitate its interpretation. In the case of diversity, a correspondence analysis (CA) was carried out based on the presence - absence of soil macrofauna families. This allowed us to identify those families that were exclusive or shared between systems. All analyzes were performed with the InfoStat software, version 2018.
RESULTS AND DISCUSSION
Composition of the richness and abundance of edaphic macroinvertebrates
The taxonomic richness was represented by two phylums, seven classes, 22 orders and 53 families of edaphic macroinvertebrates. The greatest richness was found in Bs, followed by SAFc and Mc. Despite this, the evaluated systems did not present statistically significant differences (p>0.072) in the richness of taxonomic groups (Table 1). This type of behavior may correspond to a sampling effort or insufficient statistical power; however, these results present a trend in which richness decreases as the degree of disturbance and differences in vegetation increase, as occurs with high Bs heterogeneity and Mc homogeneity The foregoing coincides with reports by Delgado et al. (2011) and Cabrera et al. (2011) who indicate that the richness of species is higher in the forest and in agroforestry systems than in production systems without the presence of trees, thanks to the diversity of the vegetation, the availability of food for the species, the complex structure of the vegetation that provides habitat sites and offers better protection than in simpler agricultural systems. Rojas et al. (2011) also affirm that coffee agroecosystems with complex vegetation help maintain a high richness of macroinvertebrates, which allows a greater similarity with the forest, as is the case of SAFc (Table 1).
Table 1. - Abundance (individuals m-2 ) and richness of soil macrofauna in coffee production systems (Mc and SAFc) and in secondary forest (Bs) in the department of Huila
| Family | Mc | SAFc | B.s | ||
|---|---|---|---|---|---|
| Amaurobiidae | 1.33 ± 1.33 | 2.67 ±1.8 | - | ns | |
| Anisolabididae | 1.33 ± 1.33 | 5.33 ±5.33 | - | ns | |
| Araneidae | 2.67 ±2.67 | - | - | ||
| Bdellidae | - | - | 5.33 ±1.54 | ||
| Cantharidae | - | 1.33 ± 1.33 | - | ||
| Carabidae | 4 ± 2.28 | 1.33 ±2.28 | 10.67 ± 4.55 | ns | ns |
| Cheliferidae | - | - | 5.33 ± 1.54 | ||
| Chthoniidae | - | 1.33 ± 1.33 | - | ||
| Cixiidae | 1.33 ± 1.33 | - | - | ||
| Clubionidae | - | 2.67 ± 2.67 | - | ||
| Coccinellidae | - | 1.33 ± 1.19 | 5.33 ± 2.37 | ns | |
| Chrysomelidae | 8 ± 6.95 | 10.67 ± 6.95 | 5.33 ± 13.9 | ns | ns |
| Cryptopidae | 8 ± 3.96 | 10.67 ± 3.96 | 16 ± 7.93 | ns | ns |
| Curculionidae | - | 8 ± 3.68 | - | ||
| Cybaeidae | 1.33 ± 1.33 | 2.67 ± 1.8 | - | ns | |
| Cydnidae | - | 5.33 ± 4.1 | 10.67 ± 10.67 | ns | |
| Dictynidae | 12 ± 9.26 | 4 ± 2.09 | - | ns | |
| Ectobiidae | 8 ± 6.3 | 9.33 ± 6.3 | 42.67 ± 12.59 | ns | ns |
| Elateridae | 1.33 ± 1.33 | 4 ± 2.87 | - | ns | |
| Forficulidae | 1.33B |
- | 10.67 |
** | |
| Formicidae | 205.33b |
325.33 |
138.67 ± 151.99 | * | ns |
| Geometridae | 2.67 ± 1.8 | - | - | ||
| Geophilidae | 1.33B |
1.33B |
21.33 |
ns | * |
| Gryllidae | - | - | 10.67 ± 1.54 | ||
| Hubbardiidae | 2.67 ± 2.67 | - | - | ||
| Isotomidae | 21.33 ± 8.22 | 5.33 ± 8.22 | 5.33 ± 16.44 | ns | ns |
| Julidae | - | - | 10.67 ± 1.54 | ||
| Labiduridae | 6.67 ± 5.38 | 16 ± 16 | - | ns | |
| Largidae | - | 8 ± 2.83 | - | ||
| Lepidopsocidae | 1.33 ± 1.33 | - | - | ||
| Linyphiidae | 17.33 ± 8.77 | 25.33 ± 8.77 | 10.67 ± 17.54 | ns | ns |
| Lumbricidae | 165.33 |
352 |
666.67 |
** | * |
| Lycosidae | 1.33 ± 1.33 | - | - | ||
| Mycetophagidae | 2.67 ± 1.8 | 5.33 ± 3.01 | - | ns | |
| Nabidae | 1.33 ± 1.33 | 1.33 ± 1.33 | - | ns | |
| Noctuidae | 5.33 ± 2.27 | 17.33 ± 6.67 | - | ns | |
| Nymphalidae | 13.33 ± 13.33 | 5.33 ± 5.33 | - | ns | |
| Parajulidae | - | 17.33 ± 17.33 | - | ||
| Pentatomidae | 1.33 ± 1.33 | - | - | ||
| Pisauriidae | - | 13.33 ± 4.33 | - | ||
| Polydesmidae | 8B |
2.67 |
21.33 |
ns | * |
| Porcellionidae | 13.33B |
10.67B |
80A |
ns | * |
| Ptiliidae | - | 5.33 ± 4.10 | - | ||
| Salticidae | - | - | 5.33 ± 5.33 | ||
| Scarabaeidae | 8 ± 8.36 | 13.33 ± 8.36 | 48 ± 16.72 | ns | ns |
| Siphonotidae | - | 8 ± 8 | - | ||
| Spirobolidae | - | 1.33 ± 1.19 | 5.33 ± 2.37 | ns | |
| Staphylinidae | 45.33 ± 14 | 52 ± 14 | 53.33 ± 28.01 | ns | ns |
| Superstitioniidae | - | - | 5.33 ± 1.54 | ||
| Tetragnathidae | 2.67 ± 3.16 | 6.67 ± 3.16 | 5.33 ± 6.32 | ns | ns |
| Theridiidae | 4 ± 4.35 | 8 ± 4.35 | 16 ± 8.69 | ns | ns |
| Trogositidae | - | 1.33 ± 1.33 | - | ||
| Trombidiidae | 1.33 ± 1.33 | - | - | ||
| Total abundance | 586.67 |
969.33 |
1216 |
* | * |
| Wealth | 8.58 ± 0.84 | 10.17 ± 0.84 | 13 ± 1.69 | ns | ns |
* = P < 0.05; ** = P < 0.01 ; ns = not significant. † corresponds to comparisons between production systems; †† corresponds to the comparisons between Bs, Mc and SAFc
Means ± standard error. Means followed by the same lowercase letter do not differ significantly between Mc and SAFc; and means followed by the same capital letter do not differ significantly between Bs, Mc and SAFc according to Fisher's LSD test p<0.05.
Correspondence analysis (Figure 1) showed that eight families (Araneidae, Lycosidae, Hubbardiidae, Trombidiidae, Cixiidae, Pentatomidae, Geometridae and Lepidopsocidae) were associated with a single system, the Mc; ten families (Clubionidae, Pisauriidae, Chthoniidae, Parajulidae, Siphonotidae, Cantharidae, Curculionidae, Ptiliidae, Trogositidae and Largidae) were only recorded in SAFc, while Salticidae, Superstitioniidae, Cheliferidae, Bdellidae, Julidae and Gryllidae were only recorded in Bs (Figure 1). The low representativeness of the reported families indicates that they are more susceptible to environmental disturbances. Families associated with Mc and SAFc were also recorded (Table 1 and Figure 1), this may be due to the fact that C. arabica is found covering most of the soil, providing a similar habitat in the two ideal systems for these organisms (Campera et al., 2022)

Abundance of individuals per system
22.320 individuals m-2 were recorded (Figure 2), with a general average of 826.7 individuals m-2 per monolith. The highest abundance on average occurred in B.s, followed by S.A.F.c, these two systems being statistically similar, but presenting significant differences with Mc (p<0.014) (Table 1).

Fig. 2. - Total abundance of soil macrofauna (at family level) in coffee production systems and secondary forest
There were differences (p<0.049) between the coffee production systems (Table 1, Figure 2), showing that the macroinvertebrate communities present in the soil use systems are determined by the availability of resources for their survival and the levels of soil intervention in the development of agricultural activities. The results show that there is a direct positive relationship between less disturbed systems with the abundance of edaphic macroinvertebrates, as argued by other authors Delgado et al. (2011).
In this sense, it is suggested that the main factor influencing the pattern of abundance was plant heterogeneity; the result is due to the fact that the diversification of productive systems with other plant species improves the abundance of macroinvertebrates Delgado et al. (2011), although these effects are also conditioned by the application of insecticides and fertilizers.
The taxonomic families that presented the highest abundance in the three systems were: Lumbricidae, Formicidae, Staphylinidae, Porcellionidae, Scarabaeidae and Ectobiidae. In the same way, Rojas et al. (2011). found that in most of the macrofauna found in Colombian coffee systems, earthworms and ants dominate in abundance. These six (6) families represented 80.7 % of the edaphic macroinvertebrates identified throughout the study, while the "Other" groups corresponding to 47 families represented 19.3 % of the taxonomic groups as they presented low density in the systems evaluated (> 2%).
However, only three families (Lumbricidae, Formicidae and Porcellionidae) of the most abundant and three of the "other" category (Forficulidae, Geophilidae and Polydesmidae) presented significant differences (p< 0.01), evidencing a potential use as bioindicators of soil quality and anthropic disturbances in coffee systems. The abundance of Lumbricidae presented significant statistical differences between the evaluated systems, reaching the highest value in Bs, followed by SAFc and lower abundance in Mc (p< 0.01) (Table 1). This taxonomic order is characterized by participating in the regulation of the dynamics of organic matter and the biogeochemical cycle of nutrients, in the neutralization of toxins generated by agrochemical products and in the modification of the soil structure.
On the other hand, the families of Geophilidae, Polydesmidae and Porcellionidae were statistically similar within the same families in both SAFc and Mc, however, significant differences were found compared to Bs (Table 1), with p<0.031, p <0.045 and p<0.036 respectively. This is due to the fact that these families require humid environments because of their loss of water, an attribute that Bs provides.
Regarding the Formicidae family, it presented greater abundance in the SAFc compared to the Mc with significant differences being found between them (p<0.046) (Table 1). The Forficulidae family presented greater abundance in Bs compared to Mc, finding significant differences between them (p<0.001) (Table 1); mainly because they are organisms that are sensitive to land use because they often live in deep tunnels (Cabrera, 2011).
The Others group corresponds to the remaining 47 family groups that individually contribute less than 2 % each. The abundance of individuals in S.A.F.c did not present statistical differences with B.s, a similar result was presented by Delgado et al. (2011) in a coffee-banana agroforestry system, showing that the macrofauna present in the different land use systems is determined by the availability of resources for their survival and by the intensity of agricultural activities, which suggests that agroforestry systems with coffee can be a viable option to conserve the abundance of edaphic macroinvertebrates.
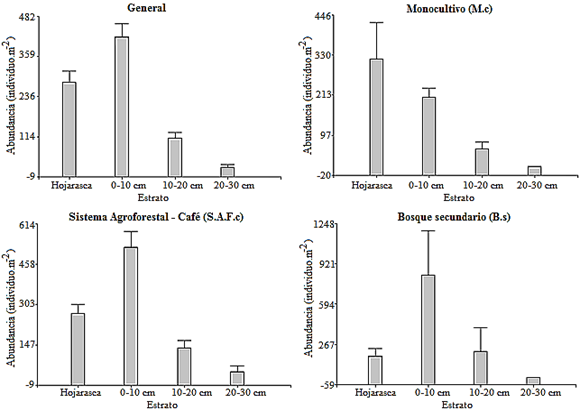
General
The greatest abundance was found in the 0-10 cm stratum, with significant differences with respect to the litter stratum and other depths (Figure 3); this agrees with the findings of Olayemi et al., (2022) who establish that the macrofauna is confined to the upper part, in the first centimeters of the soil. Similarly, there were significant differences in the litter in relation to the depths of 10-20 cm and 20-30 cm. These last two strata were statistically similar to each other.
M.c
Greater abundance was found in the litter layer, probably due to the greater contribution of litter in the superficial part, thus influencing the formation of a more favorable habitat for the development of these organisms. However, it is statistically similar to the depth of 0-10 cm, nevertheless, there was a statistically significant difference in litter with respect to the strata of 10-20 cm and 20-30 cm; also, between the 0-10 cm stratum compared to the 20-30 cm stratum (Figure 3). This decrease in macrofauna in the deep strata may be due to the fact that as the soil profile deepens, the porosity, oxygen and organic matter content that provides habitat and food decrease (Vanolli et al., 2023).
The foregoing supports what was mentioned by Armbrecht and Perfecto (2001), that the coffee monoculture is an exemplary case of how the intensification of agriculture negatively affects tropical biodiversity, because by reducing the presence of shade trees, fauna associated with crops also diminishes. In part, this is explained by the decrease in the structural complexity of the crop and, therefore, of the ecological niches of the fauna resident in it (Rodríguez et al., 2019). In this context of simplification, it is possible to study how the interspecific relationships between the macroinvertebrates that inhabit agroecosystems are affected and how these relationships can favor or harm the interests of the farmer.
S.A.F.c.
Greater abundance was found in the 0-10 cm stratum, presenting statistically significant differences with respect to the litter stratum and other depths. Similarly, there were significant differences in the litter in relation to the depths of 10-20 cm and 20-30 cm; these last two depths, despite having different abundances, were statistically similar (Figure 3). (Doblas et al.,2009 point out that one of the causes that leads to a greater presence of organisms in the first strata (superficial and 0-10 cm) is due to the amount of organic matter due to the plant cover which provides biomass and generates microhabitats favorable for the life of the macrofauna. Duran et al. (2023), suggest that the greater structural complexity and botanical composition of agroforestry systems promote the richness and diversity of the edaphic macrofauna by improving the physical conditions, the organic matter content of the soil, enhancing this by improving its processes of aggregation and porosity.
B.s
The stratum that presented the greatest abundance was 0-10 cm deep (Figure 3). This behavior may be related to the soil that is formed by the addition of leaf litter, in addition, the presence of the tree in the systems makes temperatures and the impact of rain decrease compared to open land, constituting alternatives to achieve the colonization (Lavelle et al., 2022).
CONCLUSIONS
The abundance and distribution of the edaphic macrofauna are highly sensitive attributes when the forest is replaced by coffee production systems. However, the implementation of coffee cultivation in agroforestry systems emerges as an important strategy for the conservation of these organisms.
The most abundant families (Lumbricidae, Formicidae and Porcellionidae) and less abundant (Forficulidae, Geophilidae and Polydesmidae) present a high susceptibility to changes in coverage in the systems.
In the coffee agroforestry system, the greatest diversity of macroinvertebrates is observed in the first 10 cm of soil, while in the monoculture the greatest diversity is found in the litter.
REFERENCIAS BIBLIOGRÁFICAS
ARARAT, M.C., y PRAGER, M., 2002. Efecto de cinco manejos agroecológicos de un Andisol (Typic Dystrandept) sobre la macrofauna en el municipio Piendamó, departamento del Cauca, Colombia. Acta Económica. vol 51, no. (3/4) 121-129. Disponible en: https://revistas.unal.edu.co/index.php/acta_agronomica/article/view/47796/48927 [ Links ]
ARMBRECHT, I., y PERFECTO, I., 2001. Diversidad de artrópodos en los agroecosistemas cafeteros. Revista Protección Vegetal, vol 12 no. 2: 11-16. Disponible en: http://hormigas.univalle.edu.co/pdf/2001ArtropodosCafetales.pdf [ Links ]
BORROR, D.J., TRIPLEHORN, C.A., y JOHNSON, N.F., 1992. An introduction to the study of insects [en línea]. 6ta. Estados Unidos: Sunders College Publishing. [consulta: 09/06/2023]. ISBN 9780030253973. Disponible en: Disponible en: https://latam.casadellibro.com/libro-an-introduction-to-the-study-of-insects-6th-ed/9780030253973/831454 . [ Links ]
CABRERA DÁVILA, G., SOCARRÁS RIVERO, A., HERNÁNDEZ VIGOA, G., PONCE DE LEÓN, D., MENÉNDEZ RIVERO, Y., y SÁNCHEZ RENDÓN, J., 2017. Evaluación de la macrofauna como indicador del estado de salud en siete sistemas de uso de la tierra, en Cuba/Evaluation of the macrofauna as indicator of the health status in seven land use systems, in Cuba. Pastos y Forrajes [en línea], vol. 40, no. 2, Disponible en: https://www.researchgate.net/publication/320191245_Evaluacion_de_la_macrofauna_como_indicador_del_estado_de_salud_en_siete_sistemas_de_uso_de_la_tierra_en_CubaEvaluation_of_the_macrofauna_as_indicator_of_the_health_status_in_seven_land_use_systems_in_. [ Links ]
CABRERA, G., ROBAINA, N., y PONCE DE LEÓN, D., 2011. Composición funcional de la macrofauna edáfica en cuatro usos de la tierra en las provincias de Artemisa y Mayabeque, Cuba. Pastos y Forrajes [en línea], vol. 34, no. 3, Disponible en: https://www.redalyc.org/pdf/2691/269121083008.pdf. [ Links ]
CAMPERA, M., BUDIADI, B., BUŠINA, T., FATHONI, B., DERMODY, J., NIJMAN, V., IMRON, M., y NEKARIS, K.A., 2022. Abundance and richness of invertebrates in shade-grown versus sun-exposed coffee home gardens in Indonesia. Agroforestry Systems [en línea], vol. 96, no. 5-6, DOI 10.1007/s10457-022-00744-9. Disponible en: https://www.researchgate.net/publication/360606929_Abundance_and_richness_of_invertebrates_in_shade-grown_versus_sun-exposed_coffee_home_gardens_in_Indonesia. [ Links ]
CERQUERA LOSADA, Ó.H., PÉREZ GÓMEZ, V.H., y SIERRA CHAVARRO, J., 2020. Análisis de la competitividad de las exportaciones del café del Huila. Tendencias [en línea], vol. 21, no. 2, [consulta: 12/06/2023]. ISSN 0124-8693, 2539-0554. Disponible en: Disponible en: https://dialnet.unirioja.es/servlet/articulo?codigo=7646596 . [ Links ]
DELGADO MIRANDA, G., BURBANO, A., y SILVA PARRA, A., 2011. Evaluación de la macrofauna del suelo asociada a diferentes sistemas con café Coffea arabiga L. Revista de Ciencias Agrícolas [en línea], vol. 28, no. 1, [consulta: 12/06/2023]. ISSN 2256-2273, 0120-0135. Disponible en: Disponible en: https://dialnet.unirioja.es/servlet/articulo?codigo=5104119 . [ Links ]
DOBLAS MIRANDA, E., SÁNCHEZ PIÑERO, F., y GONZÁLEZ MEGÍAS, A., 2009. Vertical distribution of soil macrofauna in an arid ecosystem: Are litter and belowground compartmentalized habitats? Pedobiologia [en línea], vol. 52, no. 6, [consulta: 12/06/2023]. ISSN 0031-4056. DOI 10.1016/j.pedobi.2008.11.006. Disponible en: Disponible en: https://www.sciencedirect.com/science/article/pii/S0031405609000043 . [ Links ]
DURÁN, B. E.H., ANGEL, S. Y.K., BERMÚDEZ, M.F., y SUÁREZ, J.C., 2023. Agroforestry systems generate changes in soil macrofauna and soil physical quality relationship in the northwestern Colombian Amazon. Agroforest Syst 97, 927-938. https://doi.org/10.1007/s10457-023-00838-y [ Links ]
INSTITUTO GEOGRÁFICO AGUSTÍN CODAZZI. 1994. Estudio General de suelos del Departamento del Huila, Santafé de Bogotá. IGAC [ Links ]
INSTITUTO GEOGRÁFICO AGUSTÍN CODAZZI., 2014. Estudio general de suelos y zonificación de tierras departamento de Caquetá, escala 1.100.000. Bogotá. 410p IGAC. [ Links ]
LAVELLE, P., MATHIEU, J., SPAIN, A., BROWN, G., FRAGOSO, C., LAPIED, E., DE AQUINO, A., BAROIS, I., BARRIOS, E., BARROS, M. E., BEDANO, J. C., BLANCHART, E., CAULFIELD, M., CHAGUEZA, Y., DAI, J., DECAËNS, T., DOMINGUEZ, A., DOMINGUEZ, Y., FEIJOO, A., SUPP, S., 2022. Soil macroinvertebrate communities: A world-wide assessment. Global Ecology and Biogeography, 31, 1261-1276. https://doi.org/10.1111/geb.13492 [ Links ]
OLAYEMI, P.O., SCHNEEKLOTH, J. P., WALLENSTEIN, M.D., TRIVEDI, P., CALDERÓN, F.J., CORWIN, J., y FONTE, S. J., 2022. Soil macrofauna and microbial communities respond in similar ways to management drivers in an irrigated maize system of Colorado (USA), Applied Soil Ecology, vol. 178. https://doi.org/10.1016/j.apsoil.2022.104562. [ Links ]
ROBAINA RODRÍGUEZ, N., MÁRQUEZ, S.M., y RESTREPO, L.F., 2019. The edaphic macrofauna in three components of the coffee plant arrangement associated with different management typologies, Antioquia, Colombia. Revista de la Facultad de Ciencias Agrarias UNCuyo [en línea], vol. 51, no. 2, Disponible en: https://bibliotecas.uncuyo.edu.ar/explorador3/Record/I11-R107article-2597/Description#tabnav. [ Links ]
ROJAS MÚNERA, D.M., FEIJOO MARTÍNEZ, A., MOLINA RICO, L.J., ZÚÑIGA, M.C., y QUINTERO, H., 2021. Differential impact of altitude and a plantain cultivation system on soil macroinvertebrates in the Colombian Coffee Region. Applied Soil Ecology [en línea], vol. 164, [consulta: 13/06/2023]. ISSN 0929-1393. DOI 10.1016/j.apsoil.2021.103931. Disponible en: Disponible en: https://www.sciencedirect.com/science/article/pii/S0929139321000524 . [ Links ]
RUIZ, N., y LAVELLE, P., 2008. Soil macrofauna field manual. Technical level [en línea]. Roma: FAO. [consulta: 13/06/2023]. Disponible en: Disponible en: https://docplayer.net/159975720-Soil-macrofauna-field-manual-technical-level.html . [ Links ]
VANOLLI, B. S., PEREIRA, A. P.A., FRANCO, A. L.C., y CHERUBIN, M. R., 2023. Edaphic and epigeic macrofauna responses to land use change in Brazil, European Journal of Soil Biology, vol. 117. https://doi.org/10.1016/j.ejsobi.2023.103514. [ Links ]
VELÁSQUEZ, E., y LAVELLE, P., 2019. Soil macrofauna as an indicator for evaluating soil based ecosystem services in agricultural landscapes. Acta Oecologica [en línea], vol. 100, [consulta: 13/06/2023]. ISSN 1146-609X. DOI 10.1016/j.actao.2019.103446. Disponible en: Disponible en: https://www.sciencedirect.com/science/article/pii/S1146609X18302613 . [ Links ]
Received: June 01, 2023; Accepted: August 03, 2023

