










 Serviços customizados
Serviços customizados texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
La madera es un material orgánico producto de procesos fisiológicos de crecimiento de los árboles, que posibilita la fijación de carbono en compuestos orgánicos como celulosa, hemicelulosa, lignina, entre otros (Nabeela et al., 2015; Hallet al., 2017); es un material que se ha implementado a lo largo de la historia de la humanidad para el desarrollo de estructuras habitacionales, herramientas y fuente de energía (Kiat et al., 2019; Prãger et al., 2019). En la actualidad, la demanda mundial de madera continúa en crecimiento debido a su adaptabilidad a condiciones ambientales (Cesprini et al., 2020) , bajo requerimiento energético para la transformación del material (Yang et al., 2019), atractivo estético que varía según la especie (Nabeela et al., 2015), y biosostenibilidad que simplifica el manejo de los residuos en comparación a otras materias primas (Hall et al., 2017; Careviäc et al., 2019; Kiat et al., 2019).
La región tropical presenta una amplia variedad de especies maderables, lo cual infiere en que la oferta de maderas sea amplia (Simo-Tagne et al., 2016). Costa Rica cuenta con más de 2 000 especies arbóreas identificadas (Powers et al., 2009), pero con caracterizaciones de la madera limitadas a un reducido número de especies en lo que respecta al conocimiento de las propiedades físicas, mecánicas y anatómicas (Raãssa et al., 2014).
La diversidad de especies maderables en las regiones tropicales implica que el mercado de venta sea amplio (Holm et al., 2018), incidiendo que en muchos casos se comercialice una especie mal identificada, lo cual genera en ventas erróneas de especies, se mezclen las especies por características visuales similares o los precios de venta se sobre o subestimen (Schmidt et al., 2019). Existe la necesidad de disponer de sistemas de identificación de especies que permitan la comercialización justa de madera (Knauf, 2016); tradicionalmente, se utilizan características físicas para la identificación (Machado et al., 2019), aspectos como el color, olor, sabor, textura, tipo de nudo, reventaduras o dureza son implementados para diferenciar las maderas (Kobayashi et al., 2019). Sin embargo, hay especies que por su alta similitud o procedencia muestran aspectos muy similares, siendo necesario el análisis macro o microscópico para la diferenciación, a partir de elementos anatómicos como el tipo de vaso, densidad de vasos, tipo y características del parénquima, características de las fibras; este procedimiento tradicional es un sistema lento, que requiere de especialistas y que en algunos casos precisa de consulta de xilotecas especializadas (Fernández et al., 2019).
Ante tal situación, en la última década se ha iniciado el desarrollo de metodologías alternativas de tipo no destructivas, que sean de rápida evaluación y alto nivel de precisión (Ruffinatto et al., 2014; Whitau et al., 2016). Se han implementado metodologías y tecnologías variadas como sensores de olor (Schreineret al., 2018), sistemas de identificación a partir de DeepLearning en aplicaciones portables (Affonso et al., 2017; Cao et al., 2019) e identificación de patrones hiperespectrales (Bonifazi et al., 2015). Esta última técnica, la hiperespectroscopía, consiste en la recopilación y procesamiento de información a lo largo de todo el espectro electromagnético (Li et al., 2019). Su implementación ha sido ampliada a varias temáticas científicas (Hänel et al., 2019), por ejemplo, en el uso en sensores remotos para la identificación de coberturas (Yuan et al., 2019), calidad de alimentos (Baiano, 2017), microbiología (Herrero-Langreo, 2020) y ciencias farmacológicas (Wilczyñski et al., 2016). Además de la identificación de variedades y grados de nutrición en especies agrícolas con fines productivos (Yang et al., 2010).
En el área de las ciencias naturales, la hiperespectometría, específicamente la reflectancia, se ha implementado en los últimos años en varias temáticas entre las que destacan: indicador de grado nutrición (Chi et al., 2016), diferenciador de efectos de condiciones adversas del suelo (Tan et al., 2020), diferenciador de plantas con presencia de patógenos (Ulrici et al., 2013), diferenciador de especies (Liu et al., 2019). En todos estos ámbitos se ha trabajado a nivel de hoja, debido a la simplicidad y comprensión en la estimación de dichas variables a partir de la reflectancia, absorbancia, transmitancia y fluorescencia de la hoja, aspectos que afectan directamente la capacidad fotosintética de una planta (Chi et al., 2016) y que han permitido relacionarlos con variables como color de lámina foliar, contenido de humedad foliar, densidad de nervaduras y taza de crecimiento (Li et al., 2019). Se ha encontrado que las especies tropicales tienden a formar puntos de reflectancia específicos dentro del espectro, que son únicos y que permiten la utilización de firmas hiperespectrales; en las últimas décadas se ha iniciado la creación de colecciones de firmas para especies arbustivas y arbóreas, encontrando que esta característica puede ser única en cada especie a nivel foliar (Chi et al., 2016; Machado et al., 2019).
Sin embargo, con muestras de maderas, los estudios que se han desarrollado con hiperespectroscopía han sido limitados. Estudios desarrollados por Tan et al., (2020) han demostrado relaciones entre la reflectancia entre los 300 y 450 nm con la densidad de la madera en cinco especies arbóreas de clima templado, encontrando diferenciaciones significativas entre especies, pero con homogeneidad de comportamiento dentro de cada especie. Por su parte Kobayashi et al., (2019) encontró para diez especies del género Eucaliptus spp. que no hay diferencias colorimétricas entre ellas, pero sí las respectivas firmas hiperespectrales entre los 550 y 710 nm, hallando puntos de reflectancia únicos para cada especie. Por su parte, Colares et al., (2016) con la especie Swetenia macrophylla, determinó que se pueden dar diferenciaciones en lignina, celulosa y hemicelulosa a nivel macroscópico a partir de los datos del espectro visible y el infrarrojo cercano, permitiendo conocer de forma no destructiva propiedades químicas de la madera Mvondo (2017). Finalmente, Schmidt et al., (2019) determinó para cinco especies del género Pinus spp. correlaciones entre la reflectancia hiperespectral entre 800 y 1 000 nm con la densidad y aspectos anatómicos de la madera, indicando que es una variable no destructiva para proyectar características físicas del material. Considerando esta base científica, el presente estudio tuvo como objetivo identificar patrones hiperspectrales y colorimétricos en función de aspectos anatómicos y físicos para la caracterización de la madera seca de Peltogyne purpurea Pittier procedente de planes de manejo forestal sostenible en el sur de Costa Rica.
Materiales y métodos
Especie y sitio de estudio
Se colectaron y prepararon 24 segmentos de fuste (secciones transversales, la cantidad de muestras seleccionadas se basó en disponer una potencia estadística del 95 %) de seis árboles de Peltogyne purpurea Pittier (cuatro muestras por árbol distanciadas a 4,5 m aproximadamente). Los segmentos de madera provenían de árboles con un diámetro entre 45 a 65 cm y cada muestra con un grosor de 50 mm, con un contenido de humedad inferior al 20 % y una clara presencia visual de albura y duramen. El material se obtuvo a partir de un plan de manejo forestal sostenible de bosque natural en una finca privada ubicada en la Península de Osa, Puntarenas, Costa Rica (8°53'38'' N y 83°30'51'' O). El sitio se caracterizó por tener una altitud de 350 m, con una temperatura media anual de 32 °C y precipitación anual de 4 500 mm; la topografía del terreno fue moderadamente ondulada, con pendientes hasta de 35° y suelos arcillosos. La densidad de árboles con más de 10 cm de diámetro en el bosque fue de 245 árboles. Los árboles seleccionados se distinguieron por tener un diámetro a 1,3 m sobre nivel de suelo, superior a 45 cm y estar en un mismo rango de edad, condición fistosanitaria y condiciones ambientales.
Procedimiento de medición de variables
En cada segmento de fuste se midió su contenido de humedad con el fin de que fuera uniforme y evitara ser una fuente de variación (el contenido de humedad final se trabajó entre el 12 al 15 %). A cada muestra se le seleccionó la cara más uniforme, que presentara la menor cantidad de defectos naturales (nudos, rajaduras, curvaturas, picaduras de insectos, entre otros) y mínimo deterioro por el proceso de aprovechamiento (marcas de corte, golpes, entre otros). En la cara seleccionada se determinaron tres radios desde el eje central de la muestra (Figura 1), cada radio se distanció a 120° y de midió desde el inicio de la corteza al centro de la pieza. En cada sesión transversal se midió la longitud de la albura (que para esta especie muestra una coloración amarillo claro), duramen (de una coloración morado oscuro) y la médula (una coloración generalmente morado oscuro).
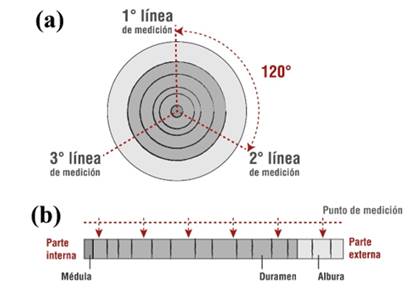
Fig. 1 - Detalle del proceso de ubicación de puntos de muestreo de los segmentos de fuste de Peltogyne purpurea
En cada línea de medición se colocaron seis puntos de control, en los que se realizaron todas las pruebas del estudio. En cada punto de medición se limpió un área de 1 cm2, en la cual se eliminó todo defecto de la madera con un bisturí. Los puntos de medición se distanciaron proporcionalmente y se evitaron colocar en el punto de transición albura-duramen. Tanto el primer como el último punto de medición se distanciaron a 30 mm del borde y de la médula de la muestra respectivamente.
Evaluación hiperespectral
La evaluación hiperespectral, específicamente de la reflectancia, se desarrolló con un espectro radiómetro portable de PP System, tipo UniSpec SC. Se trabajó con un rango de 310 a 1 100 nm (espectro visible e infrarrojo cercano), con un intervalo de medición de 10 nm y tiempo de medición programado de 4 ms. El sensor implementado contó con una certificación Estándar "SMA 905 connectors", acorde para este tipo de instrumentación, con un área efectiva de 3 mm2. El sensor se colocó perpendicular a la superficie mediante un compartimento cerrado con el fin de evitar sesgos en la medición por efecto de la luz externa. Todas las mediciones se desarrollaron en un sitio con una luminosidad homogénea de 600 lux.
Evaluación colorimétrica
El color se evaluó con un espectrofotómetro NIX Pro de cromatografía CIE estandarizado. Analiza la colorimetría entre el rango de los 400 a 700 nm con un punto de medición de 10 mm de diámetro. Para la observación de la reflexión del componente especular (SCI modo) se ajustó a un ángulo de 10, que es normal para la superficie de materiales orgánicos (D65 / 10) y con un D65 (correspondiente a la luz del día en 6 500 K). El color se evaluó en formato CIELAB, el cual generó tres parámetros para explicar el color, que consistieron en: L* (luminosidad, con una escala de 0 a 100, donde 0 es negro y 100 es blanco), a* (tendencia de color de rojo a verde, con escala infinita, conforme el valor sea más positivo tiende a rojo y más negativo a verde) y b* (tendencia de color de amarillo a azul, con una escala infinita, conforme el valor tienda a positivo será amarillo y negativo a azul).
A partir de los valores obtenidos, tanto de albura como de duramen, se compararon entre sí mediante el diferencial de color (ΔE*) que se definió como la variación neta del color entre dos escenarios, según lo establecido por la norma ASTM D 2244 (2013) cuya fórmula es detallada en la ecuación 1. El ΔE* se implementó para encontrar diferencias de color entre las fotografías y muestras físicas y con ello identificar el grado de representatividad del color, utilizando las categorías de Cuiet al. (2004), que establece cinco niveles de cambio de color percibido por el ojo humano: si 0 <ΔE*> 1,5, el cambio del color no es percibido por el ojo humano; si 1,5 <ΔE*> 3,0, el cambio de color apenas se percibe; si 3,0 <ΔE*> 6,0, se percibe el cambio; si 6,0 <ΔE*> 12,0, el cambio de color es bastante notable por el ojo humano y finalmente si ΔE* es mayor que 12, hay un cambio total de color (Ecuación 1).
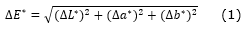
Caracterización anatómica y densidad
Anatómicamente se determinó la densidad y diámetro de vasos presentes en cada punto de medición con un área de análisis de 1 cm2. La imagen para el análisis se obtuvo mediante un microscopio de conexión electrónica marca Celestron, en modo color y con una resolución de 5 MP y las fotografías se tomaron con un aumento de 40 X; posteriormente se procesaron con el programa de Image J, según el procedimiento de Valverde et al., (2018).
Con respecto a la densidad de la madera, de varias secciones transversales se sacaron muestras cúbicas de 1 cm de cara, a las cuales se les aplicó el método de Arquímedes para determinación de la densidad, dividiendo la masa entre el volumen (se usó agua como fluido asumiendo una densidad de 1 g cm-3). Para su estimación, las muestras previamente se secaron a un contenido de humedad homogéneo del 12 %.
Análisis estadístico
Primeramente, se realizó un análisis descriptivo de las características anatómicas, colorimétricas y densidad; para determinar si se dieron diferencias significativas de las propiedades entre albura y duramen, se realizó un análisis de varianza de una vía (ANDEVA) (previamente se realizó el análisis de supuestos de normalidad de los residuos y homocedasticidad de las varianzas). En caso de presentar diferencias significativas en los resultados, se aplicó la prueba de Tukey para la comparación de las medias.
En el caso de la reflectancia, se generó la curva de reflectancia promedio y se desarrolló un ANDEVA comparativo entre los diferentes puntos, con el fin encontrar valores de diferenciación significativa. Posteriormente, se le aplicó la segunda derivada a la curva según la metodología de Romero-Ramírez et al., (2015), con el fin eliminar el ruido de aspectos químicos y físicos no considerados en el estudio y que pudieran generar error en la medición; seguidamente, a los puntos de la longitud identificados se les aplicó nuevamente un análisis de ANDEVA para validar la reflectancia. Los puntos mostraron diferenciación en ambos análisis y se les aplicó un análisis de componentes principales (ACP) con el fin de cuantificar el grado de variación existente entre albura y duramen.
Finalmente, los puntos de reflectancia con significancia se correlacionaron con las variables físicas y anatómicas mediante un análisis de correlación de Pearson. Los análisis estadísticos se realizaron con los programas STATISTICA versión 3,1 (Statsoft, 2015), con un nivel de significación de 0,05.
Resultados
Caracterización física y anatómica de la especie
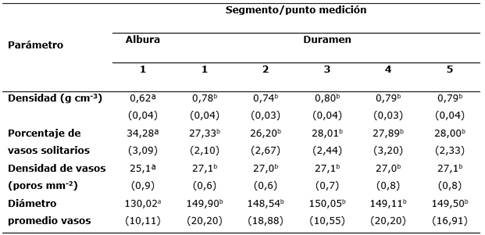
Se determinó que la madera de P. purpurea presenta variaciones significativas de la densidad y características anatómicas diferenciadas entre la albura y el duramen; encontrando que la densidad promedio en la albura es de 0,62 g cm-3, siendo significativamente menor a la obtenida en el duramen de 0,78 g cm-3 (que no mostró variaciones significativas a lo largo del duramen). De igual manera, se encontraron variaciones significativas a nivel anatómico, con una presencia mayor de vasos solitarios en la albura (34,28 %), con respecto al duramen (27,32 %), y con una densidad de 27,07 poros mm-2 y diámetro promedio de 148,53 ìm (Tabla 1).
Variación colorimétrica
A nivel colorimétrico (Tabla 2) se encontraron variaciones significativas entre albura y duramen en el parámetro L* (luminosidad) y b* (tendencia amarillo-azul), encontrando que la luminosidad de la albura (56,80) es significativamente mayor al duramen (promedio 42,07), caso similar se presentó con a*, en que el duramen presentó una coloración rojiza elevada (29,40), en comparación a duramen que presentó una tendencia neutra (1,20). En cambio, con el parámetro b* se encontraron diferencias considerables, la albura mostró una tendencia amarillenta (32,81), en comparación a duramen que en promedio presentó un valor de 1,08 considerado como neutro.
Con respecto al ΔE*, la diferencia entre las mediciones de duramen fue de 0,09, considerado como un cambio no percibido por el ojo humano, mientras la diferenciación entre albura y duramen fue superior a 14,55 y se clasificó como un cambio total de color; en este caso totalmente perceptible por el ojo humano, siendo una variable de simple diferenciación a la vista.
Tabla 2 - Caracterización colorimétrica y diferencial del color de la albura y duramen de madera seca de P. purpurea procedente de bosque natural
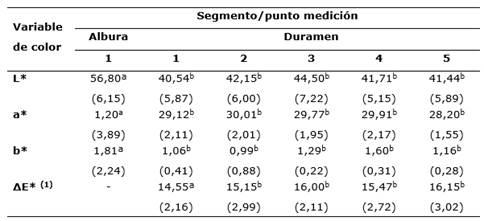
Nota: letras diferentes muestran diferencias significativas (P < 0,05); valores entre paréntesis corresponden a desviación estándar. (1) Datos del ΔE* se estimaron entre la albura con respecto a cada valor de duramen de las muestras
Comportamiento de reflectancia de la madera
Con la reflectancia (Figura 2) a se determinó que entre los valores de 310 y 450 nm no se obtuvieron diferencias en la reflectancia de albura y duramen, encontrado entre los 310 y 400 nm una oscilación de reflectancia de 0,80 a 0,92. Posterior a los 400 nm, la reflectancia mostró patrones uniformes, con un decrecimiento hasta los 450 nm de 0,212. Es a partir de los 460 nm en los que ambos segmentos de madera mostraron diferenciaciones, siendo significativamente mayor en albura que en duramen; comportamiento que se mantuvo constante hasta los 580 nm, en los que se determinó una reflectancia mayor en albura (0,234) con respecto a duramen (0,178). Seguidamente, la reflectancia fue en aumento hasta los 900 nm, presentando en el proceso un aumento mayor en el duramen que la albura, pero sin presentar diferencia significativa estadística. Es posterior a los 900 nm en que la reflectancia se empieza a estabilizar en valores cercanos de 0,844; pero con una diferenciación estadística en los 1 020 nm, en la que el duramen fue mayor a la albura.
Para eliminar los efectos de la interferencia en la reflectancia generada por compuestos químicos y físicos que podrían estar presentes en la madera dada por los efectos del secado, se determinó la segunda derivada a partir de la curva de los datos (Figura 2)b, encontrando con ello las siguientes características: la variación de la reflectancia de la segunda derivada es de ±0,0025, lo que implica que es una variación homogénea; se evidencia que en el espectro visible a los 460, 580 nm y en el infrarrojo cercano a los 1080 nm hay diferencias significativas entre albura y duramen, confirmando los patrones dados en la reflectancia; entre los 590 y 1 000 nm hay oscilaciones comunes entre albura y duramen que no muestran significancia, siendo un comportamiento similar entre ambas secciones; de los 310 a 400 nm y de los 1 050 a 1 100 nm las variaciones obtenidas no son representativas debido a la gran variación de reflectancia de dichos segmentos que la segunda derivada no es representativa.
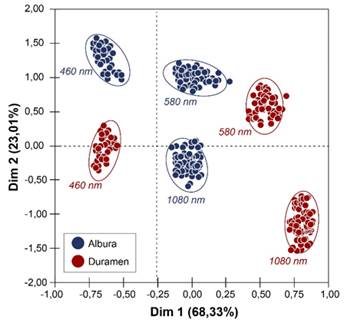
Al determinar el grado de variación con significancia de la reflectancia espectral (Figura 2) a se confirmó estadísticamente que a los 460, 580 y 1 020 nm las diferencias son significativas, con valores de diferenciación del 65 a 82 %, considerados como altos y permisibles a ser considerados como criterios de diferenciación entre ambos segmentos, siendo en el espectro visible los 460 nm como el punto de mayor variación, mientras en el infrarrojo cercano la diferenciación a los 1 020 nm es del 88 % entre ambas secciones. Con respecto al análisis de componentes principales (Figura 3) se encontraron agrupaciones de los datos de albura y duramen de las tres longitudes de reflectancia analizados que tendieron a mostrar agrupaciones diferenciadas.
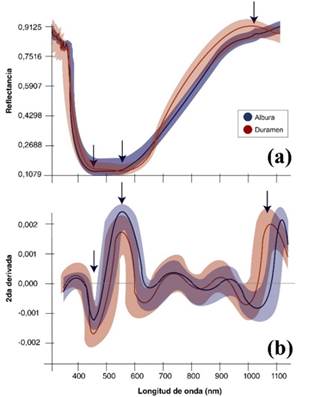
Fig. 2 - Reflectancia (a) y segunda derivada de la reflectancia (b) de las secciones de albura y duramen en madera seca de P. purpurea procedente de bosque natural
Relación de la reflectancia y variables físicas y anatómicas
Los análisis de correlación de Pearson que relacionan las características físicas y anatómicas con respecto a los tres puntos de longitud de onda en las que la reflectancia mostró variación (Tabla 3), permiten en primera instancia observar los mismos comportamientos con la albura y el duramen. No presentaron correlaciones significativas con la densidad de vasos, diámetro de vasos y porcentaje de vasos solitarios, encontrando coeficientes de correlación inferiores a 0,22. En cambio, se encontraron correlaciones significativas de las relaciones con la densidad de madera, que variaron de 0,55 a 0,60 siendo la longitud del infrarrojo cercano de 1 020 nm la que presentó la mayor correlación. Con respecto a la correlación entre las tres longitudes de onda, no fueron significativas y variaron entre 0,15 a 0,42. Por su parte, a nivel anatómico se encontraron correlaciones superiores a 0,85 entre el porcentaje de vasos solos y su diámetro, además de la densidad de la madera con la densidad de vasos. Finalmente, no se encontró correlación ente la densidad y el porcentaje de vasos solitarios.
Tabla 3 - Análisis de correlación de Pearson de los principales tres puntos de longitud de onda, en los que la reflectancia mostró significancia con datos físicos y anatómicos de las secciones de albura y duramen en madera seca de P. purpurea
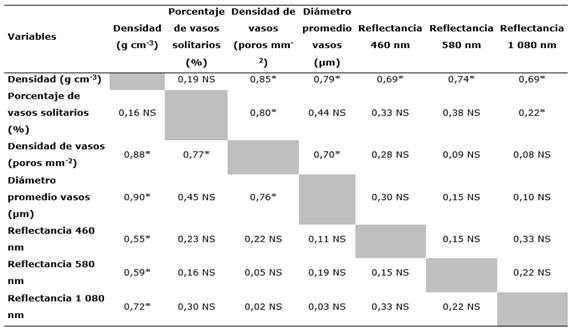
Nota: NS: diferencias no significativas (P < 0,05); parte superior de la diagonal corresponde a las correlaciones de albura, parte inferior al duramen
En la correlación entre las tres longitudes relevantes de reflectancia espectral con el color en formato CIELab (Tabla 4), nuevamente se evidenció el mismo comportamiento de correlación que el mostrado en albura y duramen; primeramente, entre las tres variables de color se obtuvieron coeficientes de correlación superiores a 0,70. En cambio, con la luminosidad (L*) no se obtuvieron correlaciones significativas con valores inferiores a 0,25; con el valor a* únicamente se mostró una correlación inversamente significativa con la longitud de 460nm (de -0,62 en albura y -0,60 en duramen). En las longitudes de 580 nm y 1020 nm no se encontraron correlaciones. Finalmente, con el parámetro b*, la única correlación significativa se registró con la longitud de 580 nm y fue mayor a 0,75.
Tabla 4 - Análisis de correlación de Pearson de los principales puntos de reflectancia y datos de color Lab en las secciones de albura y duramen en madera seca de P. purpurea
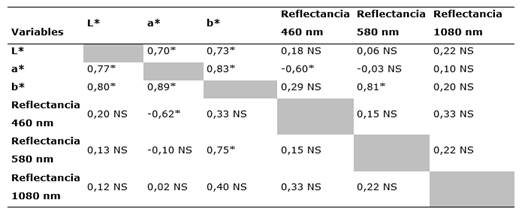
Nota: NS: diferencias no significativas (P < 0,05); parte superior de la diagonal corresponde a las correlaciones de albura, parte inferior al duramen
Discusión
Variaciones entre albura y duramen
Las diferenciaciones de las propiedades anatómicas y físicas identificadas entre la albura y el duramen del P. purpurea se explican con una mayor densidad de vasos y diámetro mayor en la albura, región que se caracteriza por ser la parte activa del xilema (Cao et al., 2019). Estudios desarrollados por Colares et al., (2016) encontraron para doce especies tropicales que el tejido xilemático de la albura es activo y moviliza entre un 10 y 20 % más de agua y nutrientes dentro del árbol; en cambio, el duramen es una región de células muertas que produce una acumulación de extractivos por subprocesos fisiológicos, oxidación de extraíbles que inciden en que la coloración de la madera sea mayor (mayor oscuridad del color). Colares et al., (2016) mencionan que las diminuciones del diámetro de vasos en el duramen se deben a la acumulación de sustancias orgánicas e inorgánicas en las paredes de los vasos, acumulación de taninos, aceites, gomas, resinas, colorantes, compuestos aromáticos, carbonato de calcio, sílice, entre otros; engrosan la pared incidiendo en que su diámetro sea menor y que en algunos casos los vasos se llenen, provocando que en el momento de cuantificación de la densidad de vasos se disminuyan. Por su parte, Machado et al., (2019) consideran que especies de latifoliadas que presenten vasos solitarios tienden a cerrar cerca del 15 % de sus vasos en el xilema por la acumulación de taninos y ceras que inciden en que su densidad se disminuya.
Es importante destacar que la inferencia de acumulación de sustancias orgánicas e inorgánicas en las paredes y disminución de la densidad incide en que la densidad de la madera aumente y que se den cambios en la coloración de la madera; Fernández et al., (2019) determinaron para tres especies latifoliadas que el aumento de la densidad de la madera del duramen se debe a las características de acumulación y movilización de nutrientes de latifoliadas, que tiende a almacenar nutrientes y en especial soluciones orgánicas en el duramen para ser empleadas en el momento en que la especie se someta al estrés hídrico, pues al presentar vasos más cerrados, la pérdida de nutrientes y agua por presiones negativas del suelo es menor, lo cual le permite una mayor probabilidad de sobrevivencia de la especie. Mientras, la diferenciación colorimétrica entre albura y duramen, según Yang et al., (2019), se debe a la acumulación de taninos y ceras en las partes inactivas de la xilema, que puede ser de 50 a 70 % mayor que la parte activa del xilema, debido a la actividad celular, movilidad hidráulica de la especie, que al momento de ser cortada y disminuir su contenido de humedad, comienza un proceso de oxidación y reducción en el duramen que incide que obtenga una coloración más oscura que la albura. Chi et al., (2016) mencionan que, según sea menor la concentración de almidones y mayor la de fenoles y taninos, la coloración se hará más oscura y rojiza.
Implicaciones de las propiedades físicas y anatómicas con la reflectancia hiperespectral
Los resultados no presentaron una correlación directa con aspectos anatómicos, esto se debió según Chi et al., (2016), a que los aspectos que mayor inferencia tienen en la reflectancia en maderas se deben a la composición química. La lignina, celulosa y hemicelulosa afectan directamente la reflectancia, además de que la presencia de extraíbles incide en que aumente la absorción de luz. Corales et al., (2016), con Swetenia macrophylla, encontraron que si bien las discrepancias anatómicas entre la albura y duramen son significativas, la diferenciación de la reflectancia se debió a la concentración de lignina, sustancias extraíbles y hemicelulosa, que fueron mayores en el duramen que en la albura, lo cual permitió encontrar diferencias a los 490, 700 y 1 050 nm; en caso de la densidad de la madera, los mismos autores mencionan que las correlaciones que se pueden presentar con la reflectancia se deben a la concentración de compuestos extraíbles que inciden en un cambio en la reflectancia en el espectro visible y en el infrarrojo, por lo cual recomienda para la identificación de patrones espectrales de especies correlacionarlos más con la parte química de la especie.
Según los resultados del presente estudio, se descartó la relación de que la anatomía de la madera de esta especie, al analizar el aspecto colorimétrico y encontrar únicamente relaciones con la longitud de 580 nm, que es donde se da la reflectancia de los colores considerados amarillos, aspecto que se denota con la correlación lineal con la variable b* y a la inversa con a*. Según Hall et al., (2017), en el sistema CIELab la coordenada b* es la que infiere en la coloración amarilla-azul, lo cual produce valores elevados en la longitud de 580 nm que infieren que la coloración amarilla aumente, mientras la coloración en 460 nm tiende a longitudes verduzcas, de ahí que la relación sea inversa con la coordenada de color a*. Por otra parte, la poca relación con la luminosidad según Holm et al., (2018) se debe a que es una variable que tiene mayor relación con la absorción de la luz por parte de la superficie, en caso de maderas es cercana al 55-65 % de la luz que incide, mientras la reflectancia es inferior al 30 %; de ahí que la luminosidad no tienda a presentar significancia con la longitud de reflectancia obtenida.
Potencialidad de la reflectancia hiperespectral como diferenciador de especies
Si bien se conoce que la reflectancia es funcional para la identificación y diferenciación de especies a nivel foliar, según mencionan Kiat et al. (2018), a nivel de maderas, los estudios han sido limitados, pero la potencialidad de aplicación es amplia. Liu et al., (2019) mencionan que la reflectancia y absorción de la luz en madera no solo permitiría la identificación de especies, posibilitaría estimar de forma no destructiva las concentraciones de celulosa, hemicelulosa y lignina, y a su vez la potencialidad de coloración de la madera al estar seca. Por su parte, Schmidt et al. (2019) destacan la potencialidad de la espectrometría como indicador de calidad de la madera para desarrollo de pellets como combustible sólido; el sistema facilitaría la evaluación de la composición química del pellets y estimación mediante modelos de potencial calórico de las especies.
Powers et al., (2009) destacan que se debe ver la reflectancia como más que un indicador de especies o composición química, debe ser entendido como una metodología de evaluación de la calidad de madera, identificación de puntos de falla del material (evidencia espectral de microfallas) y de sistemas de secado de madera. Se debe incentivar este tipo de investigaciones y enfocarlas al entendimiento de la presencia de elementos como calcio, nitrógeno, potasio, fósforo, entre otros; elementos que, según los científicos, son de relevancia en el manejo silvicultural de plantaciones y que al momento de extracción de madera se da una pérdida nutricional, que debe ser analizada para el desarrollo de paquetes de fertilización.
Se encontraron diferencias significativas a nivel anatómico entre la sección maderable de albura y duramen.
La albura presentó una mayor cantidad de vasos solitarios (38,4 %), una densidad de vasos mayores con 27,07 poros mm-2 y un diámetro de vasos promedio de 148,53 ìm.
El duramen presentó una densidad de madera significativamente mayor (0,78 g cm-3).
En colorimetría, la luminosidad y b* fue mayor en albura, pero con valores significativamente menor en a*, obteniendo un diferencial del color superior a 14,55 (cambio total del color). Con la reflectancia se encontraron tres puntos de diferenciación significativa que fueron a los 460, 580 y 1 020 nm; al correlacionar estos tres segmentos no se encontró correlación alguna con los aspectos anatómicos, pero sí con la densidad de la madera (superiores a 0,69), a nivel de color no se encontraron relaciones con el parámetro de color L*, mientras que a* solo mostró correlaciones inversas y de tipo lineal con b* a los 580 nm. Finalmente, no se encontraron relaciones colorimétricas con aspectos anatómicos.

