










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Salud Animal
versión impresa ISSN 0253-570X
Rev Salud Anim. v.29 n.2 La Habana Mayo-ago. 2007
Artículo original
POLIMORFISMO GENÉTICO EN EL LOCUS DE LA KAPPA-CASEÍNA, EN VACAS DE DIFERENTES RAZAS Y CRUCES EN EL TRÓPICO MEXICANO
GENETIC POLYMORPHISM IN THE KAPPA-CASEIN LOCUS IN COWS OF DIFFERENT BREEDS AND CROSSBREEDS IN THE MEXICAN TROPIC
Patricia Cervantes*, Mayra Luna*, A. Hernández*, F. Pérez-Gil*, P. Ponce** y Odalys Uffo**
*Facultad de Medicina Veterinaria y Zootecnia, Universidad Veracruzana. México. Correo electrónico: paty1959@yahoo.com.mx; **CENLAC, CENSA, Apartado 10, San José de las Lajas, La Habana, Cuba. Correo electrónico: pastor@censa.edu.cu
RESUMEN
En este estudio se presentan los resultados sobre la variabilidad genética en el locus de la k-caseína para seis rebaños de ganado bovino del trópico mexicano, basados en análisis de ADN por PCR-RFLP de muestras de sangre de 331 animales. Los valores estimados de frecuencias alélicas para el alelo A de esta proteína, que resultó ser el más abundante en 5 de las 6 poblaciones analizadas se encontraron entre 0.511 para el suizo y 0.702 para el Holstein, con valores de heterocigosidades observadas en todos los casos superiores a las esperadas. Basado en esta información se analizó el cumplimiento de las condiciones de equilibrio, encontrándose que el rebaño Cebú es el único que no muestra características de desequilibrio para el locus analizado. La información obtenida en el presente trabajo muestra una perspectiva de la composición de este locus en los rebaños analizados y la posibilidad de aplicación de esta tecnología para estrategias de selección, que mejoren la respuesta genética de las razas explotadas con propósitos lecheros en el trópico.
Palabras clave: variabilidad genética; proteínas lácteas; recursos genéticos.
ABSTRACT
First results on genetic variability in the k-casein locus are reported for six bovine breeds in the Mexican Tropic. They were based on DNA analysis by PCR-RFLP of blood samples from 331 animals. The estimated gene frequencies for A allele of this protein, that resulted to be most abundant in 5 of the 6 populations analysed, were between 0.511 for Swiss and 0.702 for Holstein, with heterozygosis values observed in all the cases, superior to those expected. Based on this information, the Hardy-Weinberg equilibrium conditions were analysed, being Zebu breed the unique not showing lack of equilibrium in the locus studied. The information obtained in this work gives a perspective of this locus composition in the herds analysed and it evidences possibilities of application of this technology in selection strategies that improve the genetic response of breeds with milk intentions in the tropical region.
Key words: genetic variability; milk proteins; genetic resources.
INTRODUCCIÓN
La selección en el ganado bovino se ha basado mayormente en rasgos cuantitativos tales como rendimiento de grasa, proteína y leche, los cuales se asume que están controlados por loci múltiples. La mejora genética de estos rasgos es relativamente lenta, pues los caracteres productivos solamente pueden ser medidos en un sexo, están afectados por numerosos poligenes y los factores ambientales ejercen una influencia importante en su expresión. Todo esto indudablemente disminuye la exactitud en la evaluación genética de los animales. Además, estos rasgos productivos solamente pueden ser medidos en animales adultos, lo cual incrementa el intervalo entre generaciones y disminuye el progreso genético por año.
La leche puede ser dividida en dos fracciones proteicas, las séricas (a-lactoalbúmina y b-lactoglobulina) y las cuatro caseínas (as1, as2, b, k-caseína) (10 ), cada una de las cuales exhibe al menos dos variantes genéticas. Varios estudios han reportado que algunas de dichas variantes genéticas, particularmente de la b-lactoglobulina y la k-caseína, están asociadas con el desarrollo de la lactación y tienen una influencia fundamental en la composición de la leche y sobre sus propiedades de procesamiento, incluyendo el rendimiento quesero (22, 26, 16, 1).
El polimorfismo de la leche ha recibido un interés investigativo considerable debido a su uso potencial como ayuda a la selección genética y a la caracterización genética del ganado bovino (9, 14, 31, 19).
Se han descrito relaciones entre los genotipos de las diferentes proteínas lácteas y algunas características productivas (21). De estos estudios se conoce que el genotipo AA de la b-lactoglobulina está asociado con un elevado rendimiento de leche, mientras que el BB se relaciona con un elevado contenido de grasa y caseínas, por lo tanto es más deseable para la manufactura de quesos. El alelo B de la k-caseína se asocia con elevados rendimientos de proteína total, el rendimiento de leche en la primera lactancia y en múltiples lactancias (27). La combinación LGBB, CASBB y CSN3B es favorable para la leche destinada a la producción de queso y LGBA, CASBC y CSN3A (29) y para la leche destinada al consumo como leche fluida. Esto ha sido confirmado por Formaggioni et al. (12), quienes refieren evidencias de que las leches caracterizadas por el alelo B de estas tres proteínas presentan un menor contenido de nitrógeno y mejores propiedades de coagulación que aquellas leches tipo A, por lo tanto favorables las primeras para la producción de quesos, la selección a favor de la CSN3B ejerce un efecto de arrastre favorable a los alelos LGBB y CASBB y negativo para el alelo CASA1C (32).
La determinación de los genotipos de las proteínas lácteas, puede realizarse por electroforesis, a partir de muestras de leche directamente, pues la expresión de las caseínas ocurre solamente en las células mamarias, durante la fase de lactancia, lo cual limita su uso solamente a vacas en este estado. Con las tecnologías de análisis de ADN, las cuales incluyen los métodos de la reacción en cadena de la polimerasa y el polimorfismo de longitud de los fragmentos de restricción (PCR-RFLP) ( 23), actualmente es posible determinar los genotipos, en este caso de la k-caseína de todos los individuos de una población bajo selección, independientemente del sexo, la edad o el estado fisiológico. Así, es posible incluir esta información en los programas de selección, los cuales pueden resultar en predicciones más exactas del valor genético de los animales al ser seleccionados y por lo tanto, incrementar la respuesta genética a la selección.
De acuerdo a los recursos utilizados, en México, la productividad en los sistemas de lechería tropical, se consideran bajos, con una tasa de producción anual de leche por vaca de 525 litros, se distinguen por ser fincas con terrenos menores a 140 hectáreas y con menos de 60 vacas (7). Se forma con ganado de cruzas Suizo-Cebú o Holstein-Cebú y ganado Criollo y Cebú utilizado como doble propósito. Los hatos son pequeños y medianos, su explotación es a base de pastos inducidos y en menor grado mejorada, con reproducción por proceso natural y en algunos casos por inseminación artificial. Del número total de ganado productor de leche en México, la raza Holstein es la más popular y ocupaba hasta 1997 el 90 %, el 10% restante correspondía a las razas Pardo Suizo y Jersey (25), con una tendencia de crecimiento de un 25% del total nacional, considerando que la raza Suiza, que se ha extendido de manera importante hacia el trópico, tanto por su capacidad de adaptación, como porque permite utilizarla como doble propósito (13). Las razas cebuínas que se explotan en el trópico, se caracterizan por presentar bajos rendimientos productivos y una alta adaptación a las condiciones de esta área (3). En cuanto a la ganadería criolla, esta posee un papel socioeconómico y ecológico importante en diversas regiones y se asocia a poblaciones rurales de bajos recursos económicos (2). La leche producida en esta región es utilizada en mayor proporción para la elaboración tanto industrial como artesanal de derivados lácteos, la importancia biológica del gen de k-caseína en el rendimiento lechero y el efecto de su polimorfismo constituyen una herramienta de selección asistida por marcadores en animales portadores de los alelos que favorecen una mayor concentración de sólidos de la leche.
El objetivo del presente trabajo fue el estudio de las frecuencias génicas en el locus de la k-caseína en seis hatos de diversas razas de ganado bovino (Holstein, Suizo Pardo, Criollo Lechero Tropical, Cebú, F1 Suizo x Cebú y F1 Holstein x Cebú), explotados en el trópico mexicano, específicamente en el estado de Veracruz.
MATERIALES Y MÉTODOS
Se trabajó un total de 331 animales provenientes de 7 hatos del estado de Veracruz, México pertenecientes a la raza Holstein (42 animales), Suizo (n=92), Cebú (n=25), Criollo Lechero Tropical (n=92), un F1 de cruce de Holstein x Cebú (n=45) y otro de Suizo x Cebú(n=35).
Se tomaron muestras de sangre periférica para extraer ADN genómico individual, el cual fue aislado de leucocitos por tratamiento con proteinasa K, precipitación con NaCl2M y extracción con isopropanol-etanol 70% (28). La cuantificación de ADN de cada muestra se realizó por fluorometría, utilizando el colorante Hoechst, 5ng/mL.
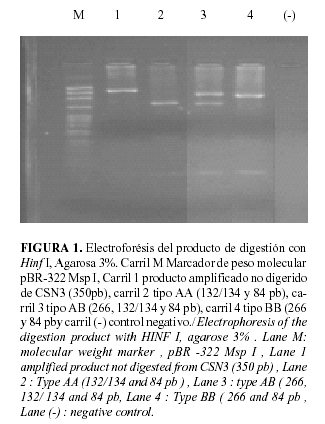
La tipificación se realizó por el método PCR-RFLP (23), utilizando una mezcla de reacción que contenía 50mM KCl, 10mM Tris-HCl, pH 8.8, 0.2mM de dNTPs, 0.5uM oligonucleótidos específicos, 0.15 mg/dl BSA, 1.0% de Tritón X-100, 1.5mM MgCl2, 0.125u/ml de Taq Polimerasa, 50 ng/L de ADN muestra, en un volumen final de 40uL. Como resultado de la amplificación del gen CSN3 se obtuvo un fragmento de 350 pb, que incluye la secuencia correspondiente al sitio de mutación de interés.
El amplicón fue digerido con la endonucleasa de restricción HinfI a 37ºC durante 3h, La digestión con las enzimas específicas se hizo con 10 uL de cada uno de los productos amplificados, en un volumen final de 20.35 uL, con 2U de enzima, 2.0 uL de buffer específico para cada una de ellas y se completó el volumen con agua desionizada. Se incubaron las mezclas de reacción a 37ºC durante toda la noche (23), para distinguir entre los alelos A y B. Tanto los fragmentos amplificados como los de restricción fueron separados en geles de agarosa al 2 y 3%, respectivamente, teñidos con bromuro de etidio, visualizándolos con un documentador de geles Genecam Spectronic.
Para el estudio de la variabilidad genética en la población analizada se utilizó el programa Biosys-1 (30). A partir de los genotipos se obtuvieron las frecuencias alélicas, el número medio de alelos por locus, la heterocigosidad estimada por conteo directo y no sesgada, así como el porcentaje de loci polimórficos, cuando la frecuencia del alelo más común era menor que 0.95 y se analizó el estado de las condiciones de equilibrio de Hardy-Weinberg en cada población.
RESULTADOS Y DISCUSIÓN
El fragmento del gen de la k-caseína amplificado fue de 350pb ubicado entre el exón IV y el intrón IV de dicho gen. Las variantes genéticas más importantes y detectadas aquí, CSN3A y CSN3B, difieren en los aminoácidos en posición 136 y 148 (Lin et al.,). En la primera posición, Thr (ACC) se cambia por Ile (GCT) y en la segunda, Asp (GAT) se sustituye por Ala (GCT) para A y B respectivamente. El cambio en la posición 148 elimina el sitio HinfI en la CSN3B que sí está presente en CSN3A (35). Ver Figura 1.
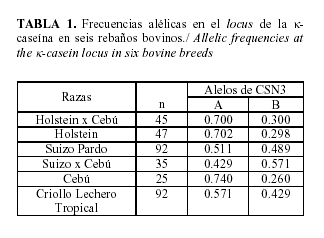
En la Tabla 1, se puede observar que en todas las razas se encontraron tres genotipos (AA, BB y AB). La raza Suizo Pardo posee un valor intermedio en la frecuencia de ambos alelos, mientras que CSN3B en SC y CLT posee valores diferentes (0.429/0.571, p<0,05), pero mayores que en las razas H, C y el cruce HC.
Para CSN3A se encontraron frecuencias elevadas en las poblaciones H, HC y Cebú (0.7 o mayor) coincidiendo con un reporte de Biase et al. (4), en Brasil con valores elevados para CSN3A (0.82) en una población cebuína. Este resultado puede estar relacionado con el origen del Cebú en la región sur de México, el cual fue importado en la década del 40´ del pasado siglo de dicho país.
En la raza Holstein la frecuencia del alelo CSN3B se encontró baja (0.298). Fue encontrado en Canadá para toros de la raza Holstein, una frecuencia de CSN3B de 0.15 (27) , mientras que Ng-Kwai-Hang et al. (26) en vacas Holstein Canadiense encontraron alelo A de CSN3, se ha asociado en la raza Cebú, a una substitución de Ile(135)?Thr y en los cruces Bos taurus x Bos indicus a una transición silenciosa del tercer codón de A/G en el sitio para el aminoacido Pro(150) (CCAÆCCG) y que en ambas razas se denomina variante A de CSN3 (12).
Los resultados de frecuencia de CLT (CSN3A/0.571 y CSN3B/0.429), difieren de forma inversa a los encontrados por Viana et al. (34) en la raza Rubia Gallega, (CSN3A 0.4703 y CSN3B 0.5297). Aún con estas diferencias en las frecuencias y por las características de origen de CLT en América, con un comportamiento productivo similar a la Rubia Gallega en cuanto a un bajo volumen de leche y rendimiento alto de sólidos, se puede asumir que el rebaño CLT analizado aún conserva caracteres genotípicos de sus ancestros europeos.
Otros autores encontraron en otras razas europeas valores similares a los de CLT para CSN3B, (15) para la raza Normando (0.46), (20) en Ayrshire (0.40) y para la raza Parda Alpina (0.48) (16). Este comportamiento fue encontrado también en un rebaño criollo de Cuba, el cual mostró una frecuencia de CSN3B de 0.3 (33), 2003).
Desde el punto de vista de estos resultados, se debe esperar que la leche con más actitud para el procesamiento industrial provenga de los animales Suizo Pardo y el cruce SC, y los menores rendimientos del Criollo, mientras que el Holstein, Cebú y el cruce HC son los menos favorecidos.
La Tabla 2 muestra los valores de heterocigosidades medias observadas y esperadas (Hobs y Hesp), el número medio de alelos por locus (A) y el porcentaje de polimórfismo (P0.95). Del análisis de los parámetros considerados como medida de estructura genética en los rebaños, se observó mayor heterogeneidad que lo esperado. Estos resultados justifican los obtenidos para el cumplimiento de las condiciones teóricas del Equilibrio Hardy-Weinberg (EHW), en las seis poblaciones.
Se encontró un exceso de heterocigosis y al igual que en la mayoría de las poblaciones especializadas, en ninguno de los seis grupos estudiados el locus está en equilibrio. Esto puede deberse a la ocurrencia de apareamientos dirigidos o controlados (no aleatorio), individuos emparentados o migración entre rebaños diferentes cercanos geográficamente. La influencia de la selección artificial ejercida sobre este locus, parece estar relacionada con algún carácter productivo a favor del cual se esta seleccionando y existe la posibilidad de que no sea posible detectar ciertos alelos para el mismo alelo una frecuencia de 0.25, lo que explica en parte los resultados de este trabajo en la raza Holstein y sus cruces HC, ya que la población estudiada deriva en gran medida de vacas y semen de origen canadiense o neocelandés. En Argentina, en una población de vacas Holstein y Criollas, encontraron para CSN3B en Holstein una frecuencia de 0.34 y para Criolla de 0.35 (14), pero ambas poblaciones estaban en equilibrio. La selección artificial que se ha desarrollado para el incremento de la producción de leche, se expresa en elevadas frecuencias del alelo CSN3A, el cual se corresponde con leche de mejor calidad para venta/proceso como leche fluida que para uso en la obtención de quesos. La frecuencia baja del alelo B para CSN3 se asocia a un control de apareamientos que reduce la posibilidad de afectar esta frecuencia en la raza Holstein (6,8,17).
La mayor frecuencia del alelo CSN3B se localiza en el cruce SC seguido del Suizo Pardo, resultado que se explica por la alta presencia de este alelo reportado para dicha raza que la hacen particularmente adecuada para el proceso industrial y con una composición superior en proteína y grasa (16).
En relación a las vacas Cebú, los resultados concuerdan con lo descrito por Escoda et al. (11) y Uffo (33), quienes reportan frecuencias elevadas del alelo A de CSN3 en ganado cebú venezolano (0.90) y cubano (0.84). En este estudio, las frecuencias son semejantes a lo presentado Kemenes (18) para ganado cebú brasileño; esto puede asociarse a deriva genética, dada por el origen y dispersión geográfica de la población cebú analizada. Se reporta en estudios de ADN mitocondrial en ganado Cebú brasileño, una gran proporción de animales derivados de cruzas antiguas de Bos taurus importado de España con Bos indicus de la India (24) , lo que explicaría la elevada frecuencia de CSN3A en cebú. La frecuencia del por PCR/RFLP, los que a su vez posean un efecto poligénico que deberá ser detectado con el uso de otros marcadores moleculares.
El locus analizado resultó ser polimórfico (100%) con igual número medio de alelos por loci (2), sin que aparezcan alelos raros en todos los rebaños (Tabla 2). Esto se debe a que los métodos utilizados para la detección de las variantes genéticas de dichas proteínas son específicos para la diferenciación entre dos alelos determinados, lo que no permite la identificación de alelos raros, poco comunes en poblaciones bajo los efectos de la selección artificial. De estar presente alguno de los alelos mencionados, los resultados obtenidos no se verían afectados, dada la baja prevalencia de aparición de los mismos.
El análisis del polimorfismo molecular de k-caseína en los diferentes genotipos permitirá una aproximación a las características importantes, en relación con la expresión productiva y composicional de la leche.
REFERENCIAS
1. Aleandri R, Butazzoni LG, Schneider JC, Caroli A, Davoli R. The effects of milk protein polymophisms on milk component and cheese-producing ability. J Dairy Sci. 1990;73:241-255.
2. Alonso MR, Ulloa RA. Hacia un proyecto nacional de investigación en genomas de animales domésticos. Veterinaria México. 1997;28(4):365.
3. Ávila S, Gasque R. Grupos genéticos de ganado bovino destinados a la producción de leche. Enciclopedia Temática Pecuaria. Facultad de Medicina Veterinaria y Zootecnia, UNAM. 2002.
4. Biase HF, Del Garnero VA, Becerra AFL, Rosa JMA, Lôbo BR, Martelli M. Analysis of restriction fragment length polymorphism in the kappa-casein gene related to weight expected progeny difference in Nellore cattle. Gen Mol Biol. 2005;28(1):84-87.
5. Bovenhuis H, Van Arandonk JAM, Korver S. Association between milk protein polymorphisms and milk production traits. J Dairy Sci. 1992;75:2549.
6. Bovenhuis H. The value of selection for ê-casein and â-lactoglobuline genotypes in dairy cattle breeding. The relevance of milk protein polymorphism for dairy cattle breeding. Wageningen Agricultural University, Netherlands. 1997;101-126.
7. Castro LCJ. Perspectivas de la red leche de bovino en México. FIRA, México. 2003.
8. Debeljak M, Susnik S, Marinsek-Logar R, Medrano JF, Dovc P. Allelic differences in bovine kappa-casein gene which may regulate gene expression. Pflugers Arch. 2000;439(3 Suppl):R4-6.
9. Del Lama SN, Zago MA. Identification of kappa-casein and beta-lactoglobulin genotypes in Brazilian Bos indicus and Bubalus bubalis population. Braz J Genet. 1996;19:73-77.
10.Eigel WN, Butler JE, Ernstrom CA, Farrel HM Jr, Harkalkar VR, Jennes R, et al. Nomenclature of proteins of cow milk: fifth edition. J Dairy Sci. 1984;67:1599-1631.
11.Escoda AB, Alvarez LO, Yerez S. Estudio de los polimorfismos genéticos de las proteínas de la leche producida en algunas hacienda de la zona de Carora. Rev Facultad Agronomía (LUZ). 1981;6(2):714-716.
12.Formaggioni P, Summer A, Malacarne M, Mariani P. Milk protein polymorphism: detection and diffusion of the genetic variants in bos genus. Università degli Studi di Parma Annali della Facoltà di Medicina Veterinaria, 1999;Vol. XII, disponible online en: http://www.unipr.it/arpa/facvet/annali/1999/
13.García BCM. Perspectivas de la ganadería tropical de México ante la Globalización. Foro de Desarrollo de Ganadería Tropical, SAGARPA. 2002.
14.Golijow CD, Giovambattista G, Ripoli MV, Dulout FN, Lojo MM. Genetic variability and population structure in loci related to milk production traits in native Argentine Creole and commercial Argenine Holstein cattle. Braz J Genet. 1999;22:395-398.
15.Grosclaude F, Mahé MF, Mercier JC, Bonnemaire J, Teissier JH. Polymorphisme des lactoprotéines de Bovinés Népalais. II. Polymorphisme des caséines "as-mineures"; le locus as2-Cn est-il lié aux loci as1-Cn, b-Cn et k-Cn? Annales de Génétique et de Sélection Animale. 1976;8:481-491.
16.Grosclaude F. Le polymorphisme génétique des principales lactoprotéines bovines. INRA: Prod Anim. 1988;1:5-17.
17.Kamiñski S. Association between polymorphism within regulatory and coding fragments of bovine k-casein gene and milk performance traits.J Anim Feed. 2000;S9:435-446.
18.Kemenes PA, Reginato LCA, Rosa AJM, Parker IU, Razook GA, Figuereido LA. et al k-casein, b-lactoglobulin and growth hormone allele frequencies and genetic distances in Nelore, Gyr, Guzerá, Caracu, Charolais, Canchin and Santa Gertrudis cattle. Gen Mol Biol. 1999;22:539-541.
19.Lara MAC, Gama LT, Bufarah G, Sereno JRB, Celegato EML, de Abreu UD. Genetic polymorphism at the k-casein locus in Penteneiro cattle. Arch Zootec. 2002;51:99-105.
20.Lin CY, McAllister AJ, Ng-Kwai-Hang KF, Hayes JF. Effects of milk protein loci on first lactation in dairy cattle. J Dairy Sci.1986;69(3):704-711.
21.Lin CY, Sabour MP, Lee AJ. Direct typing to milk proteins as an aid for genetic improvement of dairy bulls and cows: A review. Animal Breeding Abstracts. 1992;60: 1-10.
22.Marzali SA, Ng-Kwai-Hang KF. Relationship between milk protein polymorphism and cheese yielding capacity. J Dairy Sci. 1986;69:1193-1201.
23.Medrano JF, Aguilar-Córdova E. Genotyping of bovine k-casein loci following DNA sequence amplification. Biotechnology. 1990;8:144-146.
24.Meirelles FV, Artur JM, Rosa AJM, Lôbo RB, Garcia JM, Smith LC, et al. Is the American Zebu really Bos indicus? Gen Mol Biol. 1999;22(4):543-546.
25.Moro MJ, Ruíz LFJ. Mejoramiento genético de características de conformación en ganado Holstein. Vet Mex. 1998;29(4).
26.Ng-Kwai-Hang KF, Hayes JF, Moxley JE, Monardes HG. Relationship between milk protein polymorphism and major milk constituents in Holstein Friesian. J Dairy Sci. 1986;69:22-26.
27.Sabour MP, Lin CY, Keough A, Mechanda SM, Lee AJ. Effects of selection practiced on the frequencies of k-casein and b-lactoglobulin genotypes in Canadian artificial insemination bulls. J Dairy Sci.1993;76:274-280.
28.Sambrook J, Rusell DW. Molecular cloning, a laboratory manual. Cold Spring Harbor Laboratory Press. Principles of gene manipulation. An introduction to genetic engineering. (6º edicción). S.B. Primorose, R.Twyman R. Y B. Old (2001). Blackwell Sciences.
29.Sartore G, Stasio I. Prospects for the selection of dairy cattle on the basis of frequencies of specific genetic variants. Industria del latte. 1984;20:55-56.
30.Swofford DL, Selander RB. BIOSYS-1. A computer program for the analysis of allelic variation in population genetics and biochemical systematic. Release 1.7. Champaing, IL, Ilinois Natural History Survey. 1989.
31.Tambasco MD. Deteção de polimorfismo dos genes de kappa-caseína e beta-lactoglobulina em animais da raça Jersey. Monografía1998: Universidad Federal de São Carlos, SP.
32.Tejedor T, Rodellar C, Zaragoza P. Análisis de la variabilidad genética en razas bovinas, mediante estudios electroforéticos. ITEA.1986;35:133:255-237.
33.Uffo O. Aplicación de los marcadores moleculares al estudio de la biodiversidad del ganado bovino cubano. Tesis Doctoral, CENSA, 2003. Cuba.
34.Viana JL, Fernández A, Iglesias A, Sánchez L, Becerra J. Análisis by PCR/RFLPs of the most frequent k-casein genotypes Rubia Gallega cattle breed. Arch Zootec. 200;50:91-96.
35.Zardowny D, Kühnlein U. The identification of the kappa-casein genotype in Holstein dairy cattle using the polymerase chain reaction. Theor Appl Genet. 1990;80:631-634.
(Recibido 15-9-2006; Aceptado 19-1-2007)


{kind=link}
{kind=link}
{kind=link}