










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Salud Animal
versión impresa ISSN 0253-570X
Rev Salud Anim. vol.34 no.3 La Habana sep.-dic. 2012
ORIGINAL ARTICLE
Antimicrobial Resistance and genes associated to the host-microbe interaction of Pasteurella multocida isolates from swine in Western Cuba
Resistencia antimicrobiana y genes asociados a la interacción hospedero-microorganismo en aislados de Pasteurella multocida de origen porcino de la región occidental de Cuba
Ivette Espinosa, M. Báez, J. Vichi, Siomara Martínez
Division of Molecular Biology. National Center for Animal and Plant Health (CENSA), Apdo.10, San José de las Lajas, Mayabeque, Cuba. E-mail: espinosa@censa.edu.cu.
ABSTRACT
Pasteurella multocida is an important veterinary pathogen that causes a range of animal diseases, including fowl cholera, hemorrhagic septicemia in cattle and atrophic rhinitis in swine. P. multocida is generally recognized as a secondary invader contributing substantially to respiratory diseases in pigs by aggravation of lung lesions. Five capsule serogroups are routinely identified in P. multocida (A, B, D, E, and F) and each is generally associated with, but not completely restricted, to a specific host. A total of 16 isolates of Pasteurella multocida capsular genotype A and two isolates capsular genotype D were characterized for their susceptibilities to 10 antibiotics and the presence of four genes for virulence factors associated to adherence. The use of PCR showed that colonization factors (ptfA), sialidases (nanH) and outer membrane proteins (ompH) occurred in 100% porcine strains, while nanB as a further colonization-related gene was detected in 22% of isolates. The 94% of the isolates showed multiple-drug resistance. It was observed that 100% were resistant to amoxicillin and spectinomicin. The resistance profiles suggested that cephalosporins and sulphametozaxole were the drugs most likely to be active against P. multocida in vitro.
Key words: P. multocida, virulence factor, PCR, antimicrobial susceptibility.
RESUMEN
Pasteurella multocida es un patógeno de amplia distribución mundial, responsable de diversos procesos clínicos en varias especies de animales, produce el cólera aviar en aves, septicemia hemorrágica en bovinos y la rinitis atrófica en cerdos. En esta especie animal P.multocida se considera, además como un patógeno secundario que contribuye sustancialmente a los procesos clínicos respiratorios, agravando las lesiones pulmonares. Existen 5 serogrupos capsulares en P. multocida (A, B, D, E, and F) y cada uno es generalmente asociado, pero no completamente restringido a un hospedero específico. En este estudio se realizó la caracterización de la resistencia antimicrobiana frente a 10 antibióticos y la detección de cuatro genes asociados a la virulencia en 16 aislados de P. multocida tipo capsular A y 2 del tipo capsular D, todos procedentes de cerdos. Los ensayos de PCR evidenciaron la presencia de factores de colonización (ptfA), sialidasa (nanH) y la proteína de membrana externa ompH en 100% de los aislados, mientras el gen nanB asociado a la colonización solamente se detectó en 22% de los aislados. El 94% de los aislados mostraron multirresistencia a los antimicrobianos evaluados, el 100% fueron resistentes a amoxicillin y spectinomicin. Los perfiles de resistencia detectados señalan que las cephalosporina y sulphametozaxole fueron las drogas más activas contra P. multocida in vitro.
Palabras clave: P.multocida, factor virulencia, PCR, susceptibilidad antimicrobial.
INTRODUCTION
Pasteurella multocida is a worldwide-distributed pathogen responsible for a broad range of diseases in livestock (hemorrhagic septicemia, shipping fever), poultry (fowl cholera), wildlife (avian cholera) and laboratory rabbits (snuffles) (1). P. multocida is the type species of the genus including the three subspecies; P. multocida subsp. multocida, P. multocida subsp. gallicida and P. multocida subsp. Septic (2,3). This bacterium is a major pathogen of pigs, being involved in a number of different disease conditions. The main diseases associated with this organism are progressive atrophic rhinitis and pneumonic pasteurellosis, one of the most common and widespread diseases of intensively housed pigs and also playing a role to increasing the severity of primary lung lesions in pigs (4, 5).
P. multocida shows a high antigenic variability and a wide host spectrum, as well as different courses of infection which also imply enormous difficulties in producing vaccines (6). Strains of P. multocida are grouped into five capsular serogroups (serogroups A, B, D, E, and F) and are further classified into 16 somatic serotypes (serotypes 1 to 16), which are primarily based on lipopolysaccharide antigens (7). The recognized virulence factors of this organism that have been identified to date are the capsule and lipopolysaccharide (1,7). Other key factors also include diverse adhesins, toxins, siderophores, sialidases and outer membrane proteins (6,8,9).
Although antimicrobial therapy is a widely available tool for the prevention and control of clinical infections (10,11), the antibiotic resistance in pathogenic bacteria from food-producing animals and environmental sources is recognized as a global problem for public health (10, 12). Numerous previous studies have reported that the imprudent use of antimicrobials bears a high risk for the selection of resistant bacteria and promotes the spread of resistance genes located on plasmids, integrons and transposons thought the process of horizontal transfer gene (13).
The prevalence of serotypes and pathotypes of P. multocida can vary considerably according to the geographical distribution and over time in a given region. The use of control methods such as antimicrobial or autogenous vaccines requires studies in each geographical area (13). These studies are useful for identifying the presence of pathogenic strains, which are predominant , and establishing the persistence of these strains over time. A successful approach for the identification of virulent strains is the differentiation of capsular serotypes using specific antisera or the corresponding cps types by genotypic assessment, with the subsequent detection of virulence associated factors. To date, in Cuba there are no studies concerning the presence of virulence associated genes known so far in porcine P. multocida strains. The aim of this study was thus to research the presence of so far known virulence associated genes in P. multocida isolates and to evaluate the antibiotic susceptibility.
MATERIALS AND METHODS
The selected isolates of P. multocida (nP=18) were recovered from 2004 (n=10), 2010 (n=3) and 2011 (n=5) in the course of routine diagnostic procedures from diseased pigs lung. Pigs included in the study were classified into two production stages: post-weaning (4-10 weeks) and transition (10-18 weeks) belonging to five farrow-to-finish farms in several geographic areas within the western region. Bacteria were cultured on Columbia agar plus 5% sheep blood plates. All the isolates were identified by standard methods as api NE (biomeriuex), not grown in Mac Conkey agar. After identification, bacteria were lyophilized and stored at 4oC until they were used.
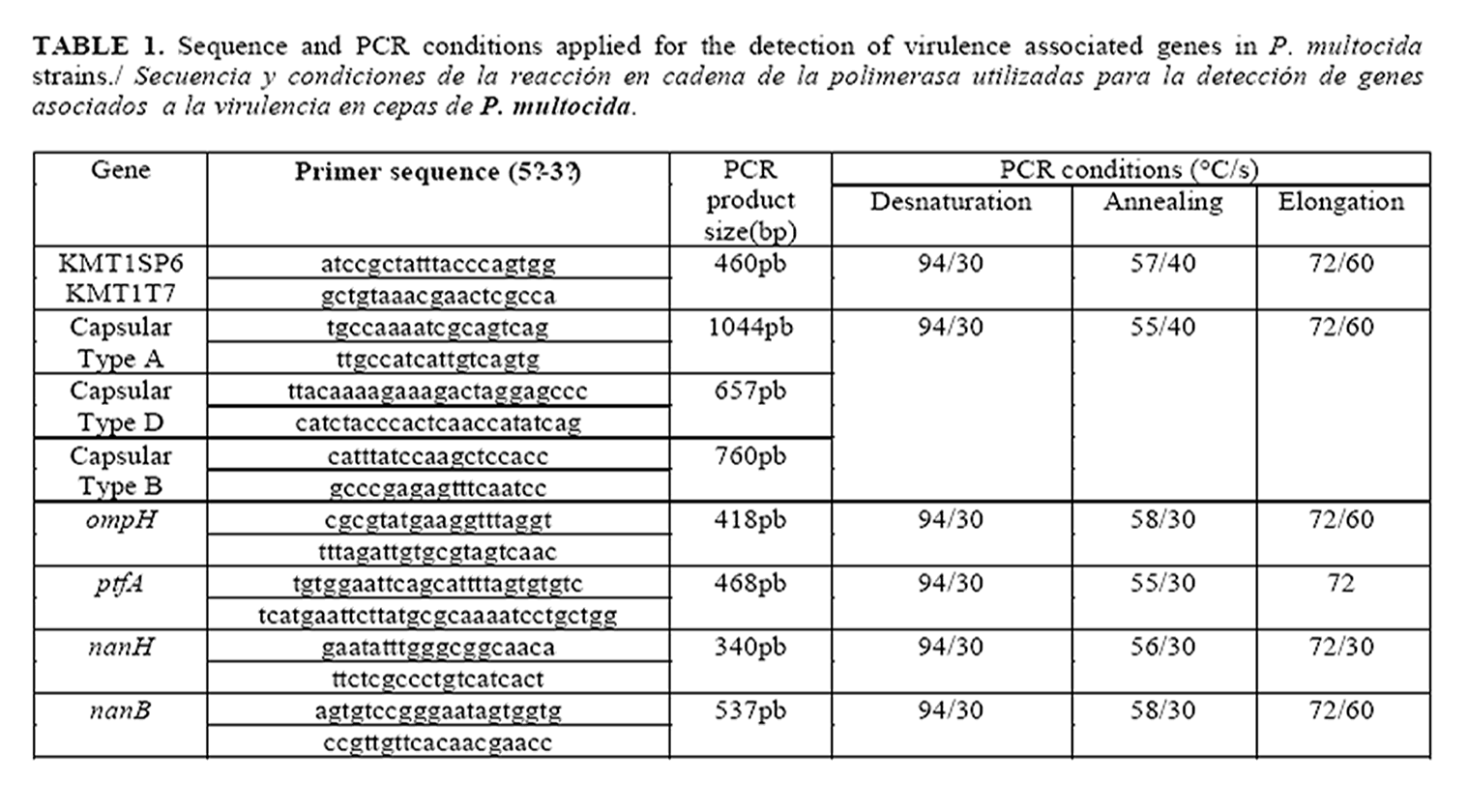
Identification of P. multocida isolates was confirmed using species-specific PCR with primers KMT1T7 and KMT1SP6, as previously described (14). The capsular types of the isolates were determined by capsule PCR typing with the capsule-specific primer pairs (primers specific for capA, capD and capB) described by Townsend et al. (15). The primer sequences used in the capsule PCR typing assay for P. multocida are listed in Table 1. Four pairs of oligonucleotide primers were used for the detection of virulence genes (VG) ompH, ptfA, nanH and nanB (6). Details of all oligonucleotide primers used and PCR conditions applied for the detection are listed in Table 1. Each isolate was tested under the same conditions twice. All oligonucleotides were custom synthesized (CIGB, Cuba).
The bacterial lysates used as templates for PCR were prepared as follows. A loopful of a bacteria colony from a fresh overnight culture on a Columbia agar plate was resuspended homogeneously in 50 µL of sterile water and the mixture was boiled at 100°C for 5 min and frozen at 20oC for 10 min to release the DNA and centrifuged to 3000g (14). A 5 µL volume of the supernatant was used as a template for each 25 µL PCR mixture containing 20 pmol of each primer, 0,4 mM of dNTPs, 1x PCR buffer, 3 mM MgCl2 and 1 U Taq DNA polymerase (Invitrogen).
The amplified DNA was visualized in 2% agarose gels in TBE buffer (90 mM Tris, 90 mM borate, 2.5 mM EDTA [pH 8]) for 1h at a constant voltage of 125V. Amplified products were stained with ethidium bromide (0.5µg/µL) and detected by UV transillumination. The 50pb ladder (Promega) was used as standard.
Antimicrobial susceptibility was determined by disk diffusion methods by following CLSI guidelines (16). The antimicrobial agents used and their respective concentration were as follows: amoxicillin (30µg); neomycin (30µg); erythromycin (15µg); tetracycline (30µg); trimethoprim/sulphamethoxazole (25µg), spectinomycin (25µg); amikacin (30µg), chloramphenicol (30µg), nalidixic acid (30µg) and cepholetin (30µg). They were supplied by the Oxoid. The inoculums were prepared from a 24 hour Columbia blood agar plate by suspending several colonies in 5 mL of saline solution and were adjusted to 0.5 McFarland standards. One hundred microliters of the adjusted inoculums were deposited on the surface agar Muller Hinton plate and further incubated at 37oC for 24 h. For the antimicrobials, the breakpoint values were taken from the CLSI guidelines (16).
RESULTS AND DISCUSSION
The P. multocida isolates were researched for kmt, the presence of three capsule biosynthesis genes and four virulence associated genes by PCR analyses. Pasteurellosis is one of the most common diseases of pigs worldwide (17,18). It is remarkable that P. multocida, one of the first agents specifically identified in pneumonic swine lung, continued throughout the past over 100 years as a major agent in porcine pneumonia, often mixed with viral agents and continues today without any evidence of being diminished (19). In this study the isolates of P. multocida were from post-weaning pigs and transition with lesions in the lungs and clinical signs of pneumonia. Xibiao et al 2009 detected strains of P. multocida throughout the year without seasonal variation from growing pigs 60 to 110 days old (20). A study in tonsils using high throughput bar-coded 454-FLX pyrosequencing defined the core microbiome dominated by Pasteurellaceae, in theses structures of pig that are constantly exposed to both ingested and inhaled microorganisms including both bacteria and viruses (21).
The whole 18 isolates were confirmed as P. multocida based on the kmt species specific gene PCR amplification product. The identification of P. multocida depends on standard culture methods and serotyping. These methods are labor intensive and results can be inconclusive. In recent years, genotypic methods of bacterial identification have proved beneficial in overcoming some limitations of traditional phenotypic procedures. For ease and rapidity, PCR was performed directly from single colonies, grown on agar plates in primary cultures from contaminated sites on the animal, such as tonsil and lung (14). A pipette tip was lightly touched onto a colony, and this sample was then resuspended in PCR amplification mixture. The primer pair KMT1SP6-KMT1T7 amplified a product of approximately 460 bp from all isolates of P. multocida.
P. multocida in pigs can cause two different clinical presentations: the atrophic rhinitis and pneumonia as a secondary invader. P. multocida plays a secondary in combination with other viral or bacterial infections contributing substantially to respiratory diseases in pigs by aggravation of lung lesions. These two clinical presentations have been predominantly linked to the capsule antigen D and capsule antigen A respectively (18). In this study, sixteenth isolates belong to A capsular serogroup and two isolates correspond to D capsular serogroup. The capsular PCR assay also provides a rapid and highly specific alternative to conventional capsular serotyping, there are currently only two laboratories worldwide that make and maintain the antisera required for capsular typing (15).
The atrophic rhinitis is less frequent as reported data from different countries, in the United States (17), England and Wales (18), thus the prevalence of strains of capsular serogroup D is lower than that of serogroup A. However, in Australian (22) and Asia (20,23), the prevalence of strains of capsular serogroup D is higher than that of serogroup A although atrophic rhinitis is seldom reported in these countries (20). There was a high numerous of isolates capsular type A, the two isolates capsular type D corresponding to 2011. Our findings suggest that the capsule type A and D isolates were to the same extent associated with pneumonic lesions in swine, that have been predominantly linked to the capsule antigen A in the past (6). None of the isolates amplified the fragment with capB-specific primers corresponding to the capsule biosynthetic loci of B serogroup. The strains of serogroup B have occasionally been implicated as a cause of a pneumonic disease in swine in China (20) or a septicemic disease in India, Sri Lanka and Vietnam (15,19). P. multocida is rarely considered the primary agent, but rather constitutes a part of the porcine respiratory disease complex (PRDC). However the findings of pericarditis and positive cultivation of P. multocida from the pericardial sac indicate that bronchopneumonia associated with P. multocida is a part of a more complex disease pattern affecting other organ systems in addition to the respiratory tract (24).
The ability to cause pathogenesis in such a wide range of host species suggests that P. multocida must have an armamentarium of adhesins to allow tissue colonization. However, the ways in which these molecules interact with the host have not been elucidated (25). All the isolates amplified the fragments of 418pb and 468pb corresponding to ompH and ptfA genes respectively (Figure 1). Escribe texto o la dirección de un sitio web, o bien, traduce un documento.Cancelatraducción del español al inglésThe outer membrane proteins (OMPs, Outer Membrane Proteins) are present in many Gram-negative bacteria, developing a role in pathogen-host interactions involved in nonspecific adhesion, immunomodulation, cytokine secretion, cytoskeletal changes and chemotaxis neutrófilos (26). The DNA sequence analyses of ptfA genes from various P. multocida strains showed a high degree of variation that could limit the potency of a vaccine based on this fimbrial antigen to cause a heterologous, capsule- and serotype independent protection considerably (6).

Sialidase is the only extracellular glycolytic enzyme produced by P. multocida, suggesting that this enzyme probably plays a major role in the ability of P. multocida to colonize animals. Many P. multocida isolates possess two sialidase genes (nanH and nanB) that encode enzymes with different substrate specificities (27). Of the sialidase-encoding genes studied, nanH (100%) was more frequent than nanB (22%), this last one only was present in A type isolate (n=4) while nanH in A and D types isolates. Tang et al (20) also found that of the sialidase-encoding genes studied, nanH (97.0%) was more prevalent than nanB (81.5%), in contrast Ewers et al. (6) found that of the sialidase-encoding genes, nanB was present in 100% of the P. multocida strains tested while nanH in 98.1%. These genes are part of the operon encoding two sialidases whose function is to release sialic acid from different host compounds, for example, to glycoproteins and glycolipids, thus, used as carbon source, also involved in virulence as they discover receptor cells of the mucous membranes of the host and / or reduce their defenses (27).
About the metabolism of the sialic acids, a structurally diverse group of nine-carbon keto sugars, has hypothesized that the interplay between its metabolism and the host-microbe interaction is a unifying theme for understanding diseases caused by a wide range of invasive pathogens or commensals. Other pathogens, such as Neisseria gonorrhoeae and Haemophilus spp, do not synthesize sialic acid but scavenge host-derived (environmental) sialic acids for cell surface decoration (modification) involving endogenous sialyltransferases. In addition to de novo synthesis or scavenging of sialic acid, some pathogens may also catabolize environmental sialic acid for nutrition (carbon, nitrogen and energy) or as a source of amino sugars for cell wall biosynthesis (28,29). In P. multocida , nanH or nanB genes could confer the bacterium not only growth benefit but also the property to adhere and invade host cells by unmasking host receptors (30). The nanH sialidase-deficient mutants of P. multocida have a reduced ability to replicate with host glycoconjugants as carbon sources (31).
Productive infection by bacterial pathogens relies on the expression of virulence factors that have wide ranging functions like competence, adherence, capsule synthesis and export, evading host immune responses. One of the difficulties associated with the development of a heterologous vaccine for P. multocida is that strains of this organism exhibit extensive genetic heterogeneity within and between serotypes (32). Thus, identification of an antigenic factor or DNA region conserved across capsular types or pathogenic strains irrespective of geographic origin may help to obviate this problem. The presence of the VF genes examined, which encode colonization factors (ptfA), sialidases (nanH), and outer membrane proteins (ompH), was equally in both (A and D) capsular type. These distribution patterns would support some lines of evidence suggesting the factors involved in adhesion may potentially serve as vaccine candidates that can elicit homologous protective immunity against all serogroup of P. multocida (32).
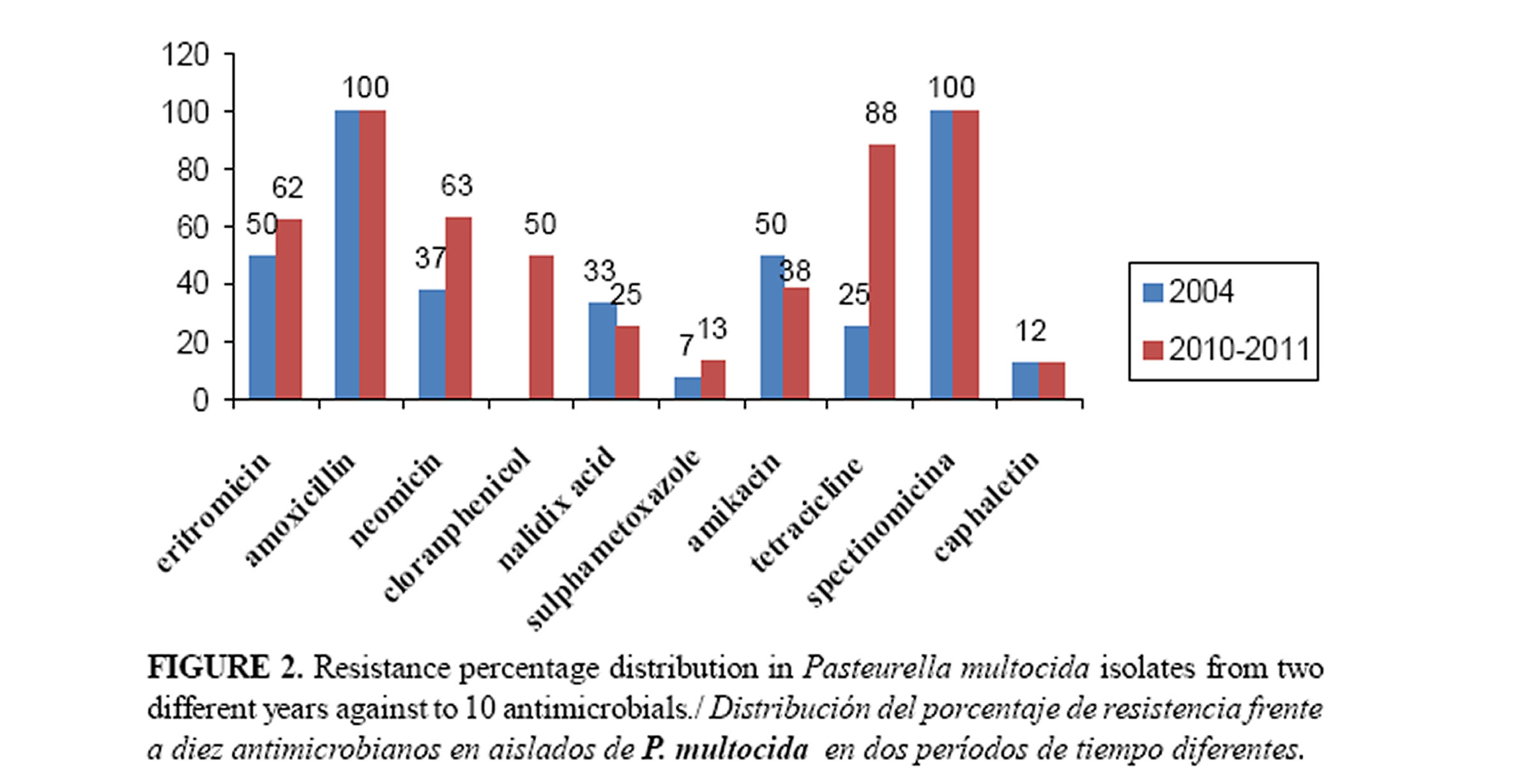
Eigteen isolates of P. multocida recovered from diseased swine were tested for resistance to 10 antibiotics. The prevalence of resistance to conventional antibiotics, including amoxicillin, spectinomicin and tetracycline among the P. multocida isolates was found to be in excess of 100-88% for each antibiotic (Figure 2). The 25% of isolates from 2004 and 88% of isolates from 2010-2011 showed a progressive increase of resistance to tetracycline. Therefore, preventive and therapeutic effects on porcine P. multocida strains should no longer be expected from these antibiotics.
Susceptibility of P. multocida isolates to eritromicin, neomicin, nalidixic acid and amikacin was variable. Of the P. multocida isolates tested corresponding to 2004, all of them were inhibited effectively by cloranphenicol; however the isolates corresponding to 2010-2011 showed a slight increase in resistance. In the present study, nalidixic acid, neomicine and amikacin exhibited moderate activity against all strains tested. Therefore, these antimicrobials could be still considered as primary therapeutic agents for the empiric treatment of P. multocida infections.
It was observed that 94% of isolates were multiresistant (resistant to from 3 to 10 antibiotics). Most of the resistance genes to date encountered among P. multocida isolates are associated with either small plasmids or with conjugative and nonconjugative transposons (33,34). Alvaro et al (35) found coexistence of multiple antimicrobial resistance plasmids in P. multocida strains from Spain and explained the resistant to tetracyclines and/or streptomycin as well as to sulfonamides.
Our results about antibiotic susceptibility, like the findings of Kehrenberg et al. (13) in France, Salmon et al. (36) in North America and Yoshimura et al. (37) in Japan, indicated that cephalosporin (cephaletin) was one of the most active drug, however in our study trimethoprim/sulphametoxazole were also an active drug. The high proportion of strains susceptible to sulphonamides (sulphathiazole) indicates that P. multocida is mainly susceptible to this antimicrobial agent. Bacteria respond to antibiotic stress with a transcriptional reflex to counteract the assault on essential processes such as cell wall synthesis, translation, transcription and replication. The transcriptional responses of P. multocida to different antibiotics with known mode of actions were collected to create a compendium of transcriptional profiles; the trimethoprim caused a marked reduction in the transcription of capsule genes, which correlated with a concomitant reduction of the capsular layer on the surface of P. multocida (38).
To the authors' knowledge, this is the first extended study of genetic diversity within a collection of P. multocida isolates from pigs using molecular methods including the amplification of virulence genes in Cuba. This is the first report indicating the occurrence of at least two capsular types A and D and the presence of gene fragments associated to the host-microbe interaction among the P.multocida isolates tested. A larger scale sampling of several pig farms from different geographical regions of the country could enable us to design a strategy to minimize the risk of exposure to P. multocida.
Studies on antibiotic susceptibility and resistance are very few and hence this study on in vitro susceptibility test on P. multocida isolates would be helpful in formulating control measures for P. multocida infection in swine. Therefore it is important to evaluate the efficacy of various antimicrobial drugs currently used in swine and to determine patterns of antibiotic sensitivity. The high prevalence of multiresistant strains of P. multocida in pigs and the association of such strains with serious disease strongly suggest that more attention should be paid to the prudent use of antimicrobials. The resistance development in isolates is a continuous process in which novel resistance genes may be acquired or developed under the selective pressure imposed by the use of new drugs (19).
REFERENCES
1. Harper M, Boyce JD, Adler B. Pasteurella multocida pathogenesis: 125 years after Pasteur. FEMS Microbiol Lett. 2006;265:1-10.
2. Christensen H, Bisgaard M, Angen O, Frederiksen W, Olsen JE. Characterization of sucrose negative variants of Pasteurella multocida including isolates from large cat bitewounds. J Clin Microbiol. 2005;43:259-270.
3. Pijoan C. Pneumonic pasteurellosis. Progressive Atrophic Rhinitis. In: Leman AD, Straw BE, Mengeling WL, D'AIliare S, De Jong MF (Eds.), Diseases of Swine. Iowa State University Press, Ames 1992:414-435.
4. Pijoan C. Pneumonic pasteurellosis. In: Straw BE, D'Allaire S, Mengeling WL, Taylor, DJ. (Eds.), Diseases of Swine. Iowa State University Press, Ames, Iowa. 1999;511-520.
5. De Jong MF. Progressive and nonprogressive atrophic rhinitis. In Diseases of Swine 9 th edition. Straw B, Zimmerman JJ, Dállaire S and Taylor DJ (eds.). Blackwell Publishing, Ames, Iowa, USA. 2006;577-602.
6. Ewers CA, Becker L, Bethe A, Kießling S, Filter M, Wieler LH. Virulence genotype of Pasteurella multocida strains isolated from different hosts with various disease status. Vet Microbiol. 2006;114(3-4):304-317
7. Harper M, Cox A, Michael FS, Parnas H, Wilkie I, Blackall PJ, et al. Decoration of Pasteurella multocida lipopolysaccharide with phosphocholine is important for virulence. J Bacteriol. 2007;189:7384-7391.
8. Adler B, Bulach D, Chung J, Doughty S, Hunt M, Rajakumar K, et al. Candidate vaccine antigens and genes in Pasteurella multocida. J Biotechnol. 1999;73:83-90.
9. Fuller TE, Kennedy MJ, Lowery DE. Identification of Pasteurella multocida virulence genes in a septicemic mouse model using signature-tagged mutagenesis. Microb Pathog. 2000;29:25-38.
10.Lion C, Conroy MC, Carpentier AM, Lozniewski A. Antimicrobial susceptibilities of Pasteurella strains isolated from humans. Int J Antimicrob Agents. 2006;27:290-293.
11.Brogden KA, Nordholm G, Ackermann M. Antimicrobial activity of cathelicidins BMAP28, SMAP28, SMAP29 and PMAP23 against Pasteurella multocida is more broad-spectrum than host species specific. Vet Microbiol. 2007;119:76-81.
12.White DG, Zhao S, Simjee S, Wagner DD, McDermott PF. Antimicrobial resistance of foodborne pathogens. Microbes Infect. 2002;4:405-412.
13.Kehrenberg C, Schulze-Tanzil G, Martel JL, Chaslus-Dancla E, Schwarz S. Antimicrobial resistance in Pasteurella and Mannheimia: epidemiology and genetic basis. Vet Res. 2001;32:323-339.
14.Townsend KM, Frost AJ, Lee CW, Papadimitriou JM, Dawkins HJ. Development of PCR assays for species- and type-specific identification of Pasteurella multocida isolates. J Clin Microbiol. 1998;36:1096-1100.
15.Townsend K, Boyce M, Chung JD, Chung AJ, Adler B. Genetic organization of Pasteurella multocida cap Loci and development of a multiplex capsular PCR typing system. J Clin Microbiol. 2001;39:924-929.
16.National Committee for Clinical Laboratory Standards. Performance Standards for antimicrobial disk and dilution susceptibility test for bacteria isolated from animals: approved standard, second ed., NCCLS document 2002.M31-A2, Wayne, USA.
17.Pijoan C. Pneumonic Pasteurellosis. In Diseases of Swine, 9 th edition. Straw B, Zimmerman JJ, Dállaire S,Taylor DJ (eds.). Blackwell Publishing. Ames, Iowa, USA.2006;719-726.
18.Davies RL, Mac Corquodale R, Baillie S, Caffrey B. Characterization and comparison of Pasteurella multocida strains associated with porcine pneumonia and atrophic rhinitis. J Med Microbiol. 2003;52:59-67.
19.Ross RF. Pasteurella multocida and its role in porcine pneumonia Anim Health Res Rev. 2007;7(1/2):13-29.
20.Tang X, Zhanqin Z, Junyong H, Bin W, Xuwang C, Qigai H, Huanchun C. Isolation, Antimicrobial Resistance, and Virulence Genes of Pasteurella multocida Strains from Swine in China. J Clin Microbiol. 2009;47(4):951-958.
21.O'Sullivan T, Friendship R, Blackwell T, Pearl D, McEwen B, Carman S, et al. Microbiological identification and analysis of swine tonsils collected from carcasses at slaughter. Can J Vet Res.2011;75:106-111.
22.Djordjevic SP, Eamens GJ, Ha H, Walker MJ, Chin JC. Demonstration that Australian Pasteurella multocida isolates from sporadic outbreaks of porcine pneumonia are non-toxigenic (toxA-) and display heterogeneous DNA restriction endonuclease profiles compared with toxigenic isolates from herds with progressive atrophic rhinitis. J Med Microbiol. 1998;47:679-688.
23.Choi C, Kim B, Cho WS, Kim J, Kwon D, Cheon DS, et al. Capsular serotype, toxA gene and antimicrobial susceptibility profiles of Pasteurella multocida isolated from pigs with pneumonia in Korea. Vet Rec. 2001;149:210-212.
24.Pors SE, Hansen MS, Bisgaard M, Jensen HE. Occurrence and associated lesions of Pasteurella multocida in porcine bronchopneumonia, Vet Microbiol. 2011;150(1-2):160-166
25.Mullen LM, Nair SP, Ward JM, Rycroft AN, Williams RJ. Novel Adhesin from Pasteurella multocida That Binds to the Integrin-Binding Fibronectin FnIII9-10 Repeats_ Infect Immun.. 2008;76(3):1093-1104.
26.Giles R, Nicky J, Mordan L Henderson B, Lin J, Huang S, Zhang, Q. Outer membrane proteins: key players for bacterial adaptation in host niches. Microb Infect. 2002;4:325-331.
27.Mizan S, Henk A, Stallings A, Maier M, Lee MD. Cloning and characterization of sialidases with 2-6' and 2-3' sialyl lactose specificity from Pasteurella multocida. J Bacteriol. 2000;182:6874-6883.
28.Vimr E R, Kalivoda K A, Deszo EL, Steenbergen S M. Diversity of Microbial Sialic Acid Metabolism. Microbiol Mol Biol Reviews. 2004;68(1):132153.
29.Chen X and Varki A. Advances in the Biology and Chemistry of Sialic Acids. ACS Chem Biol. 2010;5(2):163-176.
30.Mizan S, Henk A, Stallings A, Maier M, Lee MD. Cloning and characterization of sialidases with 2-62 and 2-32 sialyl lactose specificity from Pasteurella multocida. J Bacteriol. 2000;182:6874-6883
31.Steenbergen SM, Lichtensteiger CA, Caughlan R, Garfinkle J, Fuller TE, Vimr ER. Sialic Acid Metabolism and Systemic Pasteurellosis. Infect Immun. 2005;73(3): 1284-1294.
32.Al-Hasan K, Boyce1 J, McCarl VP, Bottomley S, Identification of novel immunogens in Pasteurella multocida Microbial Cell Fact. 2007;6(3):1-5.
33.Wilkie I, Adler B, Naas T, Benaoudia F, Lebrun L, Nordmann P. Molecular identification of TEM-1 beta-lactamase in a Pasteurella multocida isolate of human origin. Eur J Clin Microbiol Infect Dis. 2001;20:210-213.
34.Wu JR, Shieh HK, Shien JH, Gong SR, Chang PC. Molecular characterization of plasmids with antimicrobial resistant genes in avian isolates of Pasteurella multocida. Avian Dis. 2003;47:1384-1392.
35.Alvaro SM, Escudero JA, Gutierrez B, Hidalgo L, Garcia N, Llagostera M, et al. Multiresistance in Pasteurella multocida Is Mediated by Coexistence of Small Plasmids. Antimicrob agents Chemother. 2009; 3399-3404.
36.Salmon SA, Watts JL, Case CA, Hoffman LJ, Wegener HC, Yancey RJ. Comparison of MICs of ceftiofur and other antimicrobial agents against bacterial pathogens of swine from the United States, Canada and Denmark. J Clin Microbiol. 1995;33:2435-2444.
37.Yoshimura H, Ishimaru M, Endoh YS, Kojima A. Antimicrobial susceptibility of Pasteurella multocida isolated from cattle and pigs. J Vet. Med B. 2001;48:555-560.
38.Lin JT, Connelly MB, Amolo C, Otani S, Yaver DS. Global transcriptional response of Bacillus subtilis to treatment with subinhibitory concentrations of antibiotics that inhibit protein synthesis. Antimicrob Agents Chemother.2005; 49:1915-26.
Recibido: 28-10-2011.
Aceptado: 30-7-2012.

{kind=link}
{kind=link}