










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.35 no.1 La Habana ene.-mar. 2014
Ultrastructural alterations into chloroplasts and root nodules of cowpea plants grown under saline stress conditions
Alteraciones ultraestructurales en los cloroplastos y nódulos de las raíces de plantas de frijol Caupí desarrolladas bajo condiciones de estrés salino
M.Sc. Ernesto Gómez Padilla,I Dr.C. Raúl López Sánchez,II Dra.C. Beatriz Ruiz-Diez,III Dra.C. Mercedes Fernández-Pascual,IV Lic. Susana Fajardo,V Dra.C. Bettina Eichler-LoebermannVI
IAspirante Investigador, Profesor Asistente, Centro de Estudios de Biotecnología Vegetal, Facultad de Ciencias Agrícolas, Universidad de Granma, Cuba.
IIInvestigador Agregado, Profesor Titular, Centro de Estudios de Biotecnología Vegetal, Facultad de Ciencias Agrícolas, Universidad de Granma, Cuba.
IIIInvestigadora Contratada, Instituto de Ciencias Agrarias (CSIC), Madrid, España.
IVInvestigadora Científica, Instituto de Ciencias Agrarias (CSIC), Madrid, España.
VEspecialista, Instituto de Ciencias Agrarias (CSIC), Madrid, España.
VIDr. Hab. Investigadora, Universidad de Rostock, Facultad de Ciencias Agrícolas y Ambientales, Rostock, Alemania.
ABSTRACT
The ultrastructure of leaves chloroplast and root nodules from cowpea plants (Vigna unguiculata (L.) Walp.), var. IT 86 D-715, subjected to saline stress was evaluated by Transmission Electron Microscopy (TEM). Plants were exposed at 150 and 0,02 mM of NaCl treatments, considering 0,02 mM salt level as control. Native strain VIBA-1 (Bradyrhizobium liaoningense) isolated from saline soils of Cauto Valley (Cuba) was inoculated at sowing. Forty days after germination, samples of the same age and position from leaves and root nodules were taken for microscopy observations. Some ultrastructural modifications were detected by the salt effect in chloroplasts, mainly triggered by the great increase in the size of the starch granules. These modifications produced altered grana distribution. In nodule structure, when saline stress was applied, a weakness of peribacteroid membrane and high number of vesicles into infected cells were noted. Symbiosomes started deteriorating with some broken peribacteroidal membranes. A high vesiculation and degradation of some cellular organelle into uninfected cells were observed.
Key words: saline stress, ultrastructure, starch, cowpea.
RESUMEN
Se ha evaluado el efecto del estrés salino sobre la ultraestructura de los cloroplastos y los nódulos radicales de plantas de frijol Caupí (Vigna unguiculata (L.) Walp.), var. IT 86 D-715, mediante Microscopía Electrónica de Transmisión (TEM). Las plantas se sometieron a 150 y 0,02 mM de NaCl, siendo considerado este último como control. La cepa VIBA-1 (Bradyrhizobium liaoningense), aislada de suelos salinos del Valle del Cauto (Cuba), fue inoculada en el momento de la siembra. Cuarenta días después de la germinación se tomaron muestras de hojas y nódulos de la misma posición y edad para realizar observaciones microscópicas. Se han puesto en evidencia modificaciones ultraestructurales en los cloroplastos, provocadas principalmente por el incremento de tamaño de los gránulos de almidón. Estas modificaciones produjeron alteraciones en la distribución normal de los granas. En la ultraestructura del nódulo se produjo un debilitamiento de la membrana peribacteroidal y se incrementó el número de vesículas de las células infectadas, al ser aplicado el tratamiento salino. Los simbiosomas comenzaron a degradarse al romperse la membrana peribacteroidal. Dentro de las células no infectadas se observó una alta vesiculación y la degradación de algunos orgánulos celulares.
Palabras clave: estrés salino, ultraestructura, almidón, caupí.
INTRODUCTION
Legumes with wildness and adaptive characteristics to environmental stresses like cowpea (Vigna unguiculata (L.) Walp.) developed different mechanisms to reach the ionic and osmotic homeostasis into the cell (1, 2). Some of them, like increase of abscisic acid (ABA) synthesis (3), the rise of K+, Ca2+ concentration, amino acid and other ions and compatible solutes (4), the intracellular compartmentalization of Cl- and Na+ (5), the movement of them from the leaves to the roots system among others, can be mentioned (1, 6).
However, in parallel to physiological, biochemical and molecular changes, ultrastructural alterations may occur. Into the chloroplasts, the plastids and tilakoids membranes disappeared and grana are disorganized (7, 8, 9).
At the same time, into the nodule the cytoplasm disintegration, the loss of cell wall stiffness, the decrease in the packaging of inner cortex cells and the appearance of lobulated nuclei, took place. Other damages like the variations in chromatin condensation, a decrease in the volume of intercellular spaces, the increase of vesicles number and the weakening of peribacteroid membrane have been reported too (10, 11).
The analysis of cellular ultrastructure has been used as a tool to explain some response mechanisms to abiotic stress conditions in nodules and leaves of several crops like Lupin, Medicago truncatula Gaertn., Pisum sativum L. (11, 12, 13). Nevertheless, reports of ultrastructural changes that reflect damages or adaptive responses of cowpea subjected to salt stress are still unknown.
In this context, the aim of the present study was to determine the ultrastructural modifications that take place into the leave chloroplasts and root nodules of cowpea plants grown under saline stress conditions.
MATERIALS AND METHODS
Cowpea seeds (Vigna unguiculata (L.) Walp.), var. IT 86 D-715), were inoculated with a native Bradyrhizobium liaoningense strain VIBA-1 (GeneBank accession number FJ941843), isolated from nodules of cowpea grown in saline soils of Jiguaní Municipality, Granma Province, Cuba (14).
The bacteria were grown in YEM medium (15). The seeds were surface-sterilized (16), then sown in pots with vermiculite Asfaltex S.A, previously washed with distilled water and sterilized in Autoclave SC-500 Matachana for 20 min, at 120°C and 2 180 hPa. The Hoagland nutrient solution nitrogen-free was used to irrigate the plants (17). Seedlings were grown in a growth chamber at 26/19°C (day/night) temperature, 60-70 % relative humidity, 16/8 h light/dark photoperiod, a quantum irradiance of 190 µE m-2, s-2. Forty days after germination, plants were collected and samples of the same age and position from the leaves and nodules were taken for ultrastructural analyses by means of Transmission Electron Microscopy.
Sections of 1 or 2 mm2 were cut near the central nerve of the leaves with a blade of stainless steel; likewise, the nodules were cut in four little sections around 1 mm2 wide, then the samples were embedded in LR-White resin (Sigma) (18).
Nodules and leaves samples were cut with ultramicrotome Reichter Ultracut S. (Leica, Vienna, Austria) equipped with Diatome diamond blades 45°.
Ultrathin sections (70 nm) were collected on copper grills of 120-200 windows and post-stained with lead citrate and examined with a STEM LEO 910 electron microscope with an integrated digital camera Gatan Bioscan (model 792) at an accelerating voltage of 80 kv.
RESULTS
Analysis of leave chloroplasts ultrastructure
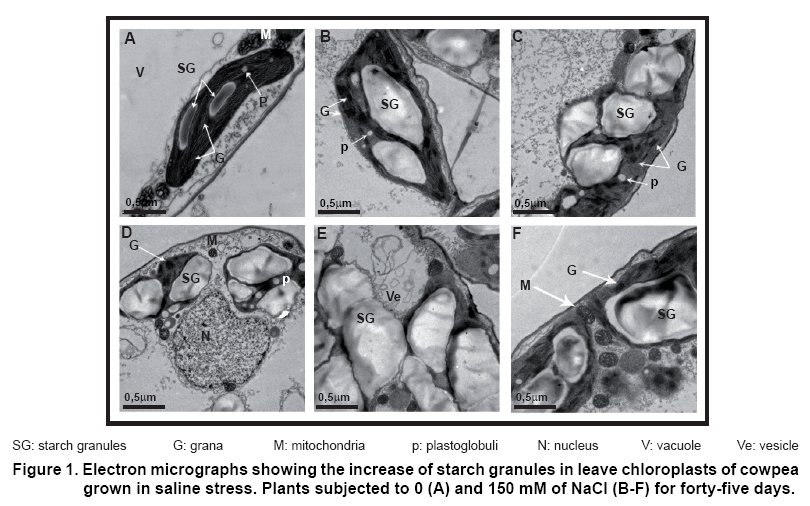
The chloroplasts of mesophyll cells from controls plants presented an organized structure with numerous grana and intergrana aligned in parallel with the chloroplast axis.
One or two elongated starch granules (amiloplasts) with normal size, some plastoglobuli (droplets of lipids) in the stroma and big mitochondria at the chloroplast ends were also observed (Figure 1 A). The size and number of starch granules increased considerably in leave chloroplasts from plants subjected to saline stress (Figure 1B-1F).
The starch granules augmented in such way that changed their original elongated form (Figure 1A), to take circular or oval shape (Figure 1B and C) with irregular borders (Figure 1C and F).
This phenomenon caused modification of normal grana distribution (Figure 1E and F), which were moved to the chloroplast edges. The grana’s appearance changed, being observed like amorphous mass very dense to the electrons, caused as a consequence of tilacoid membranes rupture, which is a senescence signal (Figure 1B and D). The major damages produced by the starch granules increased, occurred when chloroplast membranes were broken (Figure 1E).
Cellular organelle, as nucleus and mitochondrias, did not have structural modifications under saline stress (Figure 1 D and F). The appearance of vesicles were also noted, which is sign of early senescence (Figure 1H).
Analysis of nodular ultrastructure
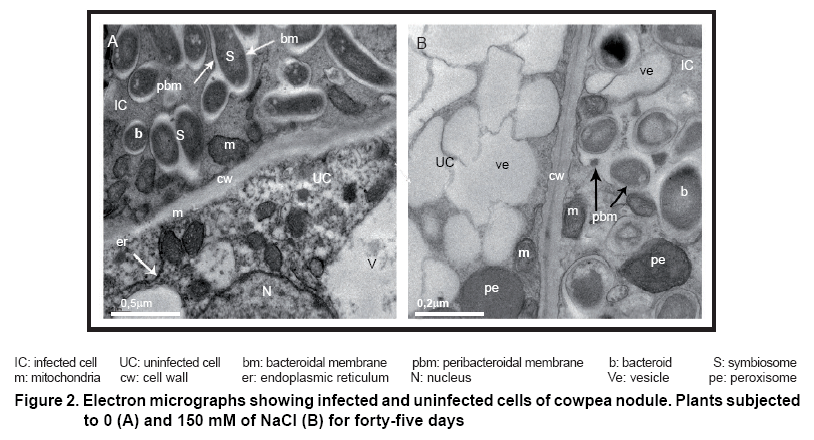
Two types of cells were found in the infection zone of cowpea nodules, infected and uninfected cells. Infected cells were plenty of bacteroids which carries out the biological nitrogen fixation. Uninfected or interstitial cells, without bacteroids, were responsible for the exchange of nitrogen products to the rest of the plant (Figure 2 A-B).
Into infected cells of nodules from plants grown in control treatment, many bacteroids surrounded by peribacteroidal membrane, and symbiosomes formed by two or three bacteroids were highlighted, (Figure 2 A).
Into uninfected cells, several mitochondria’s, endoplasmic reticulum, the nucleus and big vacuole with normal characteristics were noted. However, some variations in cell of nodules from plants subjected to saline stress were evidenced (Figure 2B).
In this sense, symbiosomes look affected by a weakness of peribacteroidal membrane, some of them become broken, being this characteristic the most important damage observed into infected cell (Figure 2B arrowheads). The uninfected cell emphasized unaltered peroxisomes, however the high number of vesicles and some signs of mitochondria deterioration pointed out senescence symptoms.
DISCUSSION
It is a well-known the fact that the starch granules of leaves, synthesized through the day and degraded at the night, need the activity of hydrolytic enzymes, which may be inhibited by adverse conditions (19, 20). A theory related with the mechanism through the plants increase in starch accumulation was explained using Arabidopsis thaliana L. and Nicotiana tabacum L. species. In these plants, the increase of starch granules in leaves correlated with a block in starch degradation, induced mainly by a loss of amylases within the plastids (chloroplasts, amiloplasts) (21).
In this experiment, the micrographs at cellular level highlighted a big accumulation of starch granules in the chloroplasts when the plants were subjected to saline stress, by resulting in some disorders in their structures. For this reason, the starch accumulation in chloroplasts seemed to be more closely related with the initial damage provoked by saline stress at cellular level, than the beneficial effect of the starch as reserve carbohydrate, as have been suggested by several authors (19, 22). This hypothesis could be attributable to the inability of cowpea plants to degrade the accumulated starch by hydrolysis during the night, exhibiting in some problems related with starch metabolism, specifically in the loss or blocked of hydrolytic amylases. Taking into consideration that synthesis and degradation of starch within the plants cells are quick processes (day-night, respectively) then, the starch accumulation could be a mechanism used by plants to mitigate the saline stress. However, this behavior maintain until the saline stress affects the enzymatic hydrolysis, by resulting in a disproportionate starch accumulation, which alter chloroplasts ultrastructure in the same way.
In our results, that phenomenon possibly took place some days before the harvest (45 days) because damages were observed at this moment, and others evidenced of harmful effect at tissue or plant level were not highlight.
Our data agreed with those obtained by several researchers who reported an increase (three-fold) of the leaf starch content in M. truncatula and Lotus japonicus L. grown under salinity stress in comparison with the control without salt (20).
On the other hand, the little variation in nodule structure (as the vesicles appearance) and the weakening of peribacteroidal membrane could be caused due to the nodule senescence, and the beginning of the harmful effect provoked by saline stress. Anyway, other morphological and ultrastructural disorders were not observed in the cells of the nodules, which indicate deep damages caused by salinity. Several authors have reported other deep lesions as cytoplasm disintegration, the stiffness loss of the cell wall, the diminishing of intercellular space and the increase of epidermis membrane and the cortex, due to the increase of vesicles number (22, 23).
The fact that in our result the infected zone of the nodule from plants subjected to salinity, was not affected in a marked way, is very beneficial to symbiotic nitrogen fixation. In the same way, this may indicate also that the nodule as a new structure formed, has developed some mechanisms to protect the symbiosome, and at the same time, to avoid the inactivation of nitrogenase enzyme.
Some mechanisms can be mentioned, such as compatible solutes accumulation as proline, sucrose, and also some structural adaptations as the swollen cell cortex which is used as a barrier to avoid the entry of Na+ ions to the infected zone (23, 24, 25). Others as the presence of a vacuolar antiporter Na+/H+, which play a relevant role in the saline tolerance, and ionic homeostasis, probably due to the induction of the accumulation of Na+ into the vacuole (26, 27) have been reported too.
ACKNOWLEDGEMENTS
This work was carried out as part of the AECID project, reference A/8500/07 and A/019119/08, developed between CSIC (Madrid, Spain) and the University of Granma, Cuba. BRD was supported by contracts of Junta de Comunidades de Castilla-La Mancha (POII09-0182-3834 and POII10-0211-5015).
REFERENCES
1. Praxedes, S. C.; de Lacerda, C. F.; DaMatta, F. M.; Prisco, J. T. and Gomes-Felho, E. Salt tolerance is associated with differences in ion accumulation, biomass allocation and photosynthesis in cowpea cultivars. J. Agron. Crop. Sci., 2010, vol. 196, p. 193-204.
2. Sprent, J. I.; Odee, W. D. and Dakora, D. F. African legumes: a vital but under-utilized resource. J. Exp. Bot., 2010, vol. 61, no. 5, p. 1257-1265.
3. Raghavendra, A. S.; Gonugunta, V. K.; Christmann, A. and Grill, E. ABA perception and signaling. Trends Plant Sci., 2010, vol. 15, no. 7, p. 395-401.
4. Maathuis, F. J. M. The role of monovalent cation transporters in plant responses to salinity. J. Exp. Bot., 2006, vol. 57, no. 5, p. 1137-1147.
5. Turkan, I. and Demiral, T. Recent developments in understanding salinity tolerance. J. Envir. Exp. Bot., 2009, vol. 67, p. 2-9.
6. Manchanda, G. and Garg, N. Salinity and its effects on the functional biology of legumes. Act. Physiol. Plant., 2008, vol. 30, p. 595-618.
7. Hashemi, A.; Abdolzadeh, A. and Sadeghipour, H. R. Beneficial effects of silicon nutrition in alleviating salinity stress in hydroponically grown canola, Brassica napus L. plants. Soil Sci. Plant. Nut., 2010, vol. 56, no. 2, p. 244-253.
8. Abdalla, M. M. Impact of diatomite nutrition on two Trifolium alexandrinum cultivars differing in salinity tolerance. Int. J. Plant Physiol. Bioch., 2011, vol. 3, no. 13, p. 233-246.
9. Amina, A. A.; Ahmed, F. K. and Gadalla, N. O. Improvement the adverse effect of salt stress in Egyptian clover (Trifolium alexandrinum L.) by AsA aplication through some biochemical and RT-PCR markers. J. Appl. Phyto. Envir. Sanit., 2012, vol. 1, no. 2, p. 91-102.
10. Perez-Guerra, J. C.; Coussens, G.; De Keyser, A.; De Rycke, R.; De Bodt, S.; Van De Velde, W.; Goormachtig, S. and Holsters, M. Comparison of developmental and stress-induced nodule senescence in Medicago truncatula. Plant Physiol., 2010, vol. 152, no. 3, p. 1574-1584.
11. Borucki, W. and Sujkowska, M. The effects of sodium chloride salinity upon growth, nodulation, and root nodule structure of pea (Pisum sativum L.) plants. Acta Physiol. Plant., 2008, vol. 30, p. 293-301.
12. Fernández-Pascual, M.; De Lorenzo, C.; de Felipe, M. R.; Rajalakshmi, S.; Gordon, A. J.; Thomas, B. J. and Minchin, F. R. Possible reasons for relative salt stress tolerance in nodules of White lupin cv. Multolupa. J. Exp. Bot., 1996, vol. 47, no. 304, p. 1709-1716.
13. Perez-Guerra, J. C.; Coussens, G.; De Keyser, A.; De Rycke, R.; De Bodt, S.; Van De Velde, W.; Goormachtig, S. and Holsters, M. Comparison of developmental and stress-induced nodule senescence in Medicago truncatula. Plant Physiol., 2010. vol. 152, no. 3, p. 1574-1584.
14. Gómez-Padilla, E.; Ruiz-Díez, B.; Fajardo, S.; Gómez-Buzón, E.; López-Sánchez, R. and Fernández-Pascual, Mercedes. Experimentos de campo en suelos salinos cubanos con plantas de Vigna unguiculata inoculadas con cepas de Bradyrhizobium nativas de la zona. In: Editores: Bonilla, I.; Zornoza, P.; Gárate, A.; Marco, E.; Rivilla, R. y Lucena, J. J. La Nutrición Mineral de las Plantas, como base de una agricultura sostenible. España. Edición libro digital 2012, p. 284-289. ISBN: 84-695-5571-5; ISBN-13: 978-84-695-5571-2.
15. Vincent, J. M. A manual for the practical study of root-nodule bacteria /J.M Vincent. In: International Programme Handbook. No. 15. Blackwele scientific publications, Oxford, England. 1970, p. 164.
16. Somasegaran, P. and Hoben, H. J. Methods in legume-Rhizobium technology. University of Hawaii NifTAL Project and MIRCEN, Department of Agronomy and Soil Science, Hawaii. Institute of Tropical Agriculture and Human Resources. 1985. 296 p.
17. Hoagland, D. R. and Arnon, D. I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Sta. Circ. 1950. vol. 347, p. 1-32.
18. de María, N.; de Felipe, M. R. and Fernández-Pascual, M. Alterations induced by glyphosate on lupin photosynthetic apparatus and nodule ultrastructure and some oxygen diffusion related proteins. Plant Physiol. Bioch., 2005, vol. 43, no. 985-996.
19. Geigenberger, P. Regulation of starch biosynthesis in response to a fluctuating environment. Plant Physiol., 2011, vol. 155, p. 1566-1577.
20. López, G. M. Metabolismo carbonado nodular en Medicago truncatula-Sinorhizobium meliloti, Lotus japonicus-Mesorhizobium loti en estrés salino: implicación de la trehalosa. Tesis de Doctarado. Universidad de Granada. Facultad de Ciencias. 2007, 315 p. ISBN: 978-84-338-4466-8.
21. Humme, E.; Osterrieder, A.; Robinson, D. G. and Hawes, C. Inhibition of Golgi function causes plastid starch accumulation. J. Exp. Bot., 2010, vol. 61, no. 10, p. 2603-2614.
22. Redondo, F. J.; de la Pena, T. C.; Morcillo, C. N.; Lucas, M. M. and Pueyo, J. J. Overexpression of flavodoxin in bacteroids induces changes in antioxidant metabolism leading to delayed senescence and starch accumulation in alfalfa root nodules. Plant Physiol., 2009, vol. 149, no. 2, p. 1166-1178.
23. Mudgal, V.; Madaan, N.; Mudgal, A.; Singh, A. and Kumar, P. Comparative Study of the Effects of Salinity on Plant Growth, Nodulation, and Legheamoglobin Content in Kabuli and Desi Cultivars of Cicer arietinum (L.). KBM J. Biol., 2010, vol. 1, p. 1-4.
24. Larrainzar, E.; Wienkoop, S.; Scherling, C.; Kempa, S.; Ladrera, R.; ArreseIgor, C.; Weckwerth, W. and González, E. M. Carbon metabolism and bacteroid functioning are involved in the regulation of nitrogen fixation in Medicago truncatula under drought and recovery. Mol. Plant Micro. Interac., 2009, vol. 22, p. 1565-1576.
25. Hussain, K.; Farrukh, M.; Majeed, A.; Nawaz, K.; Hayat, K.; Afghan, S. and Shahazad Zia-ul-Hussnian, S. What molecular mechanism is adapted by plants during salt stress tolerance? Afr. J. Biot., 2010, vol. 9, no.4, p. 416-422.
26. Coba de la Peña, T. and Pueyo, J. J. Legumes in the reclamation of marginal soils, from cultivar and inoculant selection to transgenic approaches. Agro. Sust. Develop., 2012, vol. 35, no. 1, p. 65-91.
27. Tang, R.; Li, C.; Xu, K.; Du, Y. H., and Xia, T. Isolation, functional characterization, and expression pattern of a vacuolar Na+/H+ antiporter gene TrNHX1 from Trifolium repens L. Plant Mol. Biol., 2010, vol. 28, p. 102-111.
Recibido: 27 de marzo de 2013
Aceptado: 12 de julio de 2013
M.Sc. Ernesto Gómez Padilla, Aspirante Investigador, Profesor Asistente, Centro de Estudios de Biotecnología Vegetal, Facultad de Ciencias Agrícolas, Universidad de Granma, Cuba. Email: egomezp@udg.co.cu


{kind=link}
{kind=link}