










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.35 no.3 La Habana jul.-set. 2014
COMUNICACIÓN CORTA
Respuesta de plantas de papaya silvestre (Carica cubensis Solms) al estrés hídrico y su recuperación: aspectos fisiológicos y del crecimiento
Response of papaya silvester plants (Carica cubensis Solms) to water stress and rewatering: growth and physiological aspects
Yusnier Díaz Hernández,I Dr.C. Walfredo Torres de la Noval,II Ms.C. Jesús Rodríguez Cabello,III Dr.C. Pedro Rodríguez HernándezIV
IReserva Científica del Departamento de Fisiología y Bioquímica Vegetal. Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32 700.
IIInvestigador Titular del Departamento de Fisiología y Bioquímica Vegetal. Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32 700.
IIIAspirante a Investigador del Departamento de Fisiología y Bioquímica Vegetal. Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32 700.
IVInvestigador Agregado del Departamento de Fisiología y Bioquímica Vegetal. Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32 700.
RESUMEN
Carica cubensis Solms es una especie silvestre endémica de Cuba. Son poco conocidos los aspectos fisiológicos de esta especie en condiciones de humedad limitada del suelo, por lo que el presente trabajo tuvo como objetivo estudiar la respuesta de la misma en la fase vegetativa ante la deficiencia hídrica en el suelo y su recuperación. El estudio se condujo en condiciones semi-controladas, donde las plantas fueron sometidas a 21 y 15 días de supresión y recuperación del riego respectivamente. Se monitoreó la humedad en el suelo, y se evaluaron la altura de las plantas, numero de hojas, superficie foliar, las masas secas de la raíz, tallo y hojas y el contenido foliar de clorofilas. Además, se calculó la Tasa Absoluta de Crecimiento (TAC) y Área Foliar Específica (AFE). El contenido de humedad en el suelo disminuyó hasta alcanzar valores de un 12 %, se afectó la altura, la superficie foliar, la biomasa de los órganos y el contenido de clorofilas. La relación biomasa de la raíz/biomasa de la parte aérea tendió al aumento en las plantas estresadas, lo que constituye un posible mecanismo de tolerancia a la sequía. El desarrollo de las plantas, por el comportamiento en el tiempo de la TAC, y el AFE, no sufrió modificaciones. Al establecerse el periodo de rehidratación, todas las variables mostraron una tendencia a la recuperación y, al final del periodo, la superficie foliar, el contenido de contenido de clorofilas totales, así como la TAC de la biomasa de raíz, alcanzaron valores similares al control. Se comprueba que en períodos cortos de estrés, seguido de una recuperación, no se afecta de manera general el crecimiento vegetativo de plantas de Carica cubensis.
Palabras clave: riego, superficie foliar, clorofilas.
ABSTRACT
Carica cubensis Solms is a wild species endemic from Cuba. Little is know about physiological aspects of this species under limited soil water content. The present study was conducted with the objective of evaluate the response of plant to water deficit and recovery, during vegetative stage. Assay was conducted under semi-controlled conditions and withholding water supply for 21 days and after this period, re-watering for 15 days. Water soil content was monitored periodically as well as plant height, leaves number, foliar surface, organs biomass, and total chlorophylls content. Also, Absolute Growth Rate (AGR), and Specific Leaf Area (SLA) were estimated. Soil water content decreased to values of 12 %; and plant height, foliar surface, organs biomass and total chlorophylls content were affected the most. In stressed plants, root to shoot biomass ratio increased, this constitutes a possible drought tolerant mechanism of the specie. Plant development was not affected (as evaluated by AGR) as well as SLA. During re-watering all variables tended to recover at the end, stressed plant foliar surface and total chlorophylls content reached similar values to control plants. It was conclude that short period of water deficit fallowed by re-watering do not affect Carica cubensis vegetative growth.
Key words: irrigation, foliar surface, chlorophylls.
INTRODUCCIÓN
La papaya (Carica papaya L.) es una fruta tropical que en los últimos cinco años ha tenido un crecimiento productivo a nivel mundial (1). Este cultivo se desarrolla en casi todas las áreas tropicales del mundo. Entre los principales países productores se encuentran la India, Brasil y México (2).
Numerosos autores han señalado la presencia, en nuestros ecosistemas, de especies silvestres que crecen de forma aislada, fundamentalmente en cuencas hidrográficas o de difícil acceso al hombre (3), las cuales producen abundantes frutos por planta de pequeño a mediano tamaño, y que son aprovechados por la población local.
Carica cubensis Solms es una especie silvestre endémica de Cuba pariente de las especies de papayas (Carica papaya), con un alto potencial agronómico. Esta especie puede ser utilizada particularmente en zonas marginales como reforestación, donde han sido seleccionadas para resistir condiciones de estrés, ya que contribuye a una producción sostenible con bajos insumos (4).
El conocimiento de la interacción de la papaya con factores ambientales como la luz, temperatura, humedad relativa, contenidos de humedad en el suelo y las propiedades físicas y biológicas del mismo, son necesarios para maximizar la productividad y limitar los efectos de estos factores en el proceso fotosintético, por lo que el conocimiento de la respuesta a estos factores ambientales provee las bases científicas para el desarrollo de estrategias de manejo para optimizar la calidad y productividad de las cosechas (5).
Algunos autores plantean la necesidad de un manejo adecuado en el cultivo durante la etapa de crecimiento vegetativo, frecuentemente las plantas de papaya presentan esterilidad femenina al inicio de la floración y es un aspecto normal que se corrige por sí mismo durante el desarrollo de las primeras flores. Sin embargo, esta situación puede continuar por más tiempo y convertirse en un problema económico si dichas plantas no reciben una adecuada fertilización o riego (6).
El déficit hídrico es uno de los principales factores ambientales que afecta el desarrollo de las plantas, este se pone de manifiesto de varias maneras, uno de los principales aspectos es la reducción del crecimiento (7). En plantas del género Carica, condiciones de baja humedad en el suelo han tenido una influencia negativa en el crecimiento y desarrollo de las mismas, viéndose afectado el crecimiento y la productividad (8). Otros autores han señalado las afectaciones de un período de déficit hídrico en parámetros del intercambio gaseoso en las hojas, así como la incidencia en la abscisión foliar (9).
De manera general el déficit hídrico provoca el aumento en crecimiento de la raíz respecto al tallo y parte aérea (10), el cual se considera como mecanismo de tolerancia al estrés hídrico.
Tomando como base lo planteado anteriormente, y el desconocimiento que existe sobre la fisiología de esta especie silvestre endémica de Cuba fuera de su hábitat natural es que se planteó como objetivo el estudio de la respuesta fisiológica de esta especie en la fase vegetativa frente a un estrés moderado por deficiencia hídrica y su posterior recuperación al restablecerse el riego.
MATERIALES Y MÉTODOS
El experimento se llevó a cabo en el Instituto Nacional de Ciencias Agrícolas (INCA), ubicado en San José de las Lajas, provincia Mayabeque en condiciones semicontroladas.
Para la obtención de semillas se utilizaron frutos de la especie silvestre del género Carica (Carica cubensis), recolectados en la cuenca Almendares-Vento en el noreste de La Habana. Luego de haber alcanzado una altura promedio de 60 cm en la fase de semillero, alrededor de los 40 días después de la siembra (DDS), las plantas fueron trasplantadas a recipientes de 1,67 m3 de capacidad (0,250x0,6x0,44), los cuales contenían un suelo Ferralítico Rojo Lixiviado según la nueva versión de clasificación de los suelos de Cuba (9). Todas las plantas se mantuvieron por un período de 30 días en condiciones óptimas de humedad en el suelo para su acondicionamiento, donde el agua fue aplicada manteniendo una humedad constante. Luego de esta fase se establecieron dos tratamientos, un tratamiento con riego (T0) al cual se aplicó agua manteniendo una humedad constante en el suelo, y al segundo tratamiento (T1) no se le aplicó agua.
Las plantas se sometieron a 21 días de supresión del riego y posteriormente a un periodo de recuperación durante 15 días, donde el suelo se llevó a un nivel de humedad similar al control. El monitoreo del contenido de humedad en el suelo se realizó mediante la metodología de Reflectrometría de Dominio Temporal (TDR). Para ello se utilizó un TDR Fieldscout con dos electrodos de 12 cm de longitud. Se evaluaron indicadores fisiológicos de respuesta de la planta frente al estrés hídrico como la altura, medida desde la base del tallo hasta el ápice, así como la superficie foliar mediante un medidor portátil de área foliar AM 300. El contenido de clorofilas totales fue estimado a través de un medidor portátil de clorofilas MINOLTA SPAD* 502 plus (Soil Plant Analysis Development), las mediciones se realizaron en la parte central de la tercera hoja, utilizando dos hojas por planta y tres evaluaciones por hoja. Los datos se expresaron en unidades SPAD.
Se determinaron las dinámicas de masa seca en los órganos raíz, tallo y hojas. Con los datos obtenidos se calculó como indicador de crecimiento la Tasa Absoluta de Crecimiento (TAC). Los datos de la superficie y masa seca foliar se utilizaron para calcular el Área Foliar Específica (AFE) como componente morfológico de crecimiento.
Se utilizó un diseño en bloques al azar, con dos tratamientos, cada uno de cuatro réplicas, las que estuvieron conformadas por seis plantas. Para el análisis de los datos cuantitativos se comprobó la normalidad y homogeneidad de la varianza mediante la prueba de Kolmogorov-Smirnov y la prueba de Cochran C, Hartley y Bartlett. Posteriormente, se calcularon el intervalo de confianza y el error estándar de las medias. Los análisis fueron realizados utilizando el paquete estadístico STATGRAPHICS Plus 5.1.
RESULTADOS Y DISCUSIÓN
El contenido de humedad presente en el suelo en los tratamientos estudiados se muestra en la Figura 1. Se pudo apreciar la pérdida de agua mediante los mecanismos naturales de evaporación y transpiración durante el tiempo de exposición al estrés, siendo más considerable al final del mismo (92 días después de la siembra, DDS). Luego de restablecer el riego se observa una rápida recuperación en el contenido volumétrico de agua en el suelo, tomando valores cercanos al control.
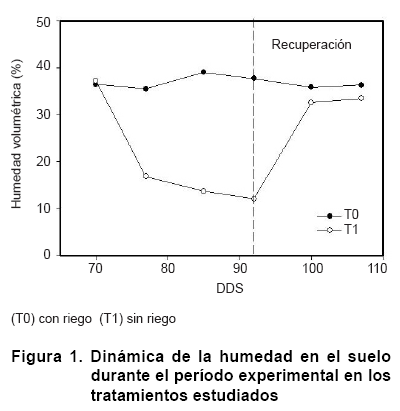
Los niveles más bajos de humedad en el suelo alcanzados en este estudio se consideran como estrés moderado para plantas de papaya (10).
En ambos tratamientos, las plantas presentaron un incremento de la altura en el período evaluado, respondiendo a la fase de crecimiento vegetativo; sin embargo, las plantas sometidas a estrés mostraron una reducción y mostraron síntomas de recuperar esta, la intensidad en el crecimiento, a partir de que la humedad del suelo resultó similar a la del tratamiento control, durante la recuperación. No obstante, las plantas con restricción hídrica se vieron afectadas en su altura, indicando la incapacidad de la planta de reponerse en esta variable (Figura 2).
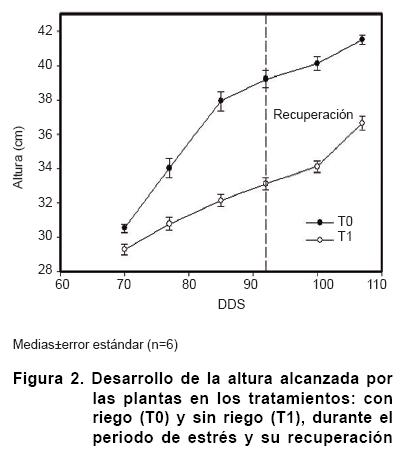
Estudios en especies del género Carica han señalado afectaciones en el crecimiento de las plantas durante la fase de crecimiento vegetativo en condiciones de déficit hídrico, teniendo como consecuencia una disminución posterior en la productividad (11).
El impacto de la supresión del riego durante la etapa evaluada provocó una reducción de hasta un 15 % en el crecimiento respecto al control a los 12 días de suprimir el riego; sin embargo, se observa una tendencia a la recuperación, donde, se aprecia una reducción de la afectación hasta el 12 %, luego de restablecer el riego.
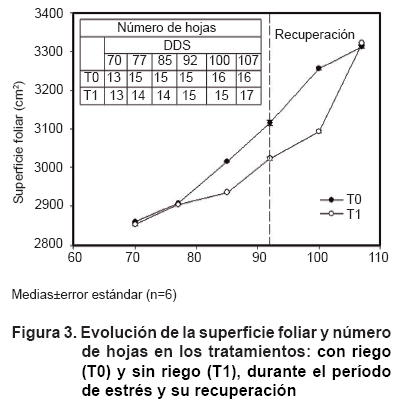
En el presente estudio el efecto de una deficiencia hídrica en el suelo tuvo su impacto en el desarrollo foliar de las plantas, encontrándose diferencias significativas a partir de los siete días de imposición del estrés en las plantas sin suministro hídrico (Figura 2), este comportamiento se mantuvo similar durante el desarrollo del estrés, alcanzando valores parecidos al control en el último momento de la recuperación, lo cual indica que las plantas de papaya pueden llegar a recuperar su superficie foliar después de un período de estrés (9).
Se ha señalado que a niveles de humedad en el suelo de hasta 23 % ocurre abscisión foliar en plantas del género Carica (10). En este trabajo la afectación no está dada por la disminución del número de hojas, sino por la reducción de la superficie foliar de las hojas nuevas que se fueron emitiendo (Figura 3).
Las condiciones de estrés por sequía influyen en los procesos del intercambio de gases a nivel de hoja. Por lo que una planta sometida a estrés de sequía presenta disminución de la fotosíntesis y de transpiración, así como reducción de la superficie foliar (14).
Las masas secas durante el estrés tuvieron un comportamiento similar al control, aunque hubo diferencias en la magnitud de los datos. Al restablecerse el riego el tallo alcanzó valores cercanos al control, no siendo así para la raíz y las hojas, aunque siguieron un patrón tendiente a la recuperación (Figura 4 A, C y E). Las TAC de los órganos evaluados presentaron patrones similares de desarrollo al control durante la imposición del estrés, aunque todos alcanzaron valores inferiores. Durante la recuperación del riego, las plantas estresadas mantuvieron una tendencia a la recuperación, siendo más favorable para la raíz (Figura 4 B, D y F).
En la tabla se observó un menor crecimiento en hojas y tallo respecto a la raíz, lo que sugiere un mecanismo de respuesta de la planta para extraer agua de las capas más profundas del suelo.
Como parte del crecimiento activo de la raíz en búsqueda de agua, el crecimiento en otras partes de la planta pudo verse afectado (10), aspecto que se pone de manifiesto en este trabajo.
El comportamiento de la TAC en las plantas en estudio concuerda con estudios realizados en especies hortícolas donde el máximo valor se alcanza en un tiempo corto o puede mantenerse por un periodo mayor, según la especie y variedad (8).
Durante la fase de crecimiento vegetativo, se han informado reducciones en el crecimiento de todos los órganos de plantas de papaya bajo déficit hídrico (17).
Se pudo observar en la tabla que el estrés provocó un aumento en la acumulación de masa seca de la raíz en comparación con la parte aérea, alcanzando valores superiores al control. Durante la recuperación se mantiene un comportamiento similar, aún con valores mayores al control pero en menor proporción, lo que puede estar dado por el traslado de reservas a la parte aérea para su recuperación (16).
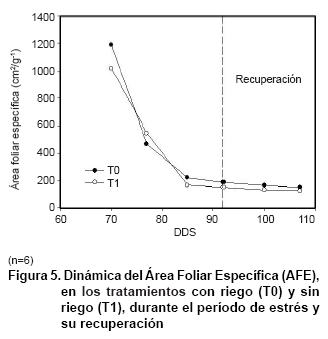
El AFE tuvo una tendencia a disminuir en ambos tratamientos, observándose un comportamiento similar entre ellos durante todo el desarrollo experimental. Este índice se refiere a consideraciones relativas al espesor de las hojas de la planta en cada período de crecimiento (18), por lo que las plantas bajo condiciones de estrés respondieron de igual forma a aquellas plantas que no presentaban restricciones hídricas.
Existe una gran correlación entre el contenido de carbohidratos totales en las hojas de las plantas y el IAF (15, 19), lo que sugiere que las plantas no estresadas en este estudio tengan una mayor eficiencia fotosintética dada por la superficie foliar y por consiguiente una mayor producción de carbohidratos totales, traducido en una alta acumulación de materia seca en los órganos (Figura 4 A, C y E). Bajo condiciones de baja humedad en el suelo, plantas de papaya sometidas a estrés han tenido afectación en la acumulación de masa seca total durante el periodo experimental, aunque no se ha evaluado la respuesta durante una fase de recuperación al estrés (20).
Cambios en el espesor de la lámina foliar han sido considerados como una respuesta adaptativa frente a condiciones de escasez de agua en plantas del género Carica (18), dicha modificación puede estar asociado el tipo de respuesta de esta especie bajo condiciones de estrés al relacionarse con el AFE (Figura 5).
Las investigaciones sobre el efecto que provocan las condiciones de estrés abiótico como la salinidad y el déficit hídrico en la concentración de pigmentos fotosintéticos son abundantes y coincidentes, y tienden a revelar que los daños ocasionados se deben fundamentalmente a la destrucción de los cloroplastos y a un aumento de la actividad de la enzima clorofilasa, lo cual afecta la síntesis de clorofilas (19).
La Figura 6 muestra las afectaciones antes mencionadas, donde las plantas sometidas a restricciones hídricas presentaron un menor contenido de clorofilas totales con diferencias significativas a los 15 días de suspendido el abastecimiento de agua, coincidiendo con datos obtenidos anteriormente (20).
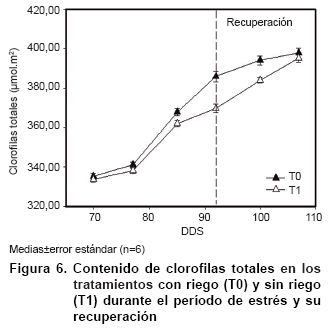
Las afectaciones en el contenido de clorofilas totales en plantas de papaya se han observado cuando estas han sido sometidas a restricciones hídricas, donde tales afectaciones son más severas a partir de los 15 días de suspendido el abastecimiento de agua (20). Estos posibles daños en el aparato fotosintético pueden haber sido una de las causas de la menor acumulación de biomasa en el tratamiento sin suministro hídrico (21).
En cultivares de cebada sensibles y tolerantes al estrés hídrico se observó un descenso rápido en el contenido de clorofilas a partir de los tres y cinco días de imposición del estrés respectivamente, lo que pudiera suponer una reducción en la actividad fotosintética por una afectación en el aparato fotosintético (20).
La respuesta en el contenido de clorofilas frente a un déficit hídrico está dada por la especie y el tiempo de exposición al estrés, así como la fase fenológica del cultivo (21).
Las plantas estresadas durante el restablecimiento del riego aumentaron su contenido de clorofilas totales respecto al periodo de estrés, lo que podría permitirle a las plantas un aumento en la eficiencia fotosintética para producir biomasa foliar y así alcanzar un desarrollo similar al control.
CONCLUSIONES
Se considera que el trabajo debe repetirse con mayor profundidad, debiendo evaluarse aspectos como la asimilación del carbono e indicadores de relaciones hídricas. Además de que sería importante determinar el impacto del déficit hídrico y su recuperación sobre el desarrollo reproductivo de las plantas.
REFERENCIAS
1. García, J. C.; Vázquez, M. E.; Torres, M. A.; Dávila, S. I. y Sánchez, D. Métodos de extracción de semilla en papaya Golden y la relación con la longevidad. Revista Mexicana de Ciencias Agrícolas, 2011, vol. 2, pp. 281-288.
2. FAO. Statistics Agricultural productions. [Consultado: 14 de octubre de 2012] Disponible en: http://www.fao.org/economic/ess/ess-publications/ess-yearbook/en/.
3. Feldberg, K.; Hentschel, J. y Wilson, R. Phylogenetic biogeography of the leafy liverwort Herbertus based on nuclear and chloroplast DNA sequence data: Correlation between genetic variation and geographical distribution. Journal of Biogeography, 2007, vol. 34, pp. 688-698.
4. Rodríguez, J.; Rodríguez, P.; González, M. E. y Martínez, P. Molecular characterization of Cuban endemism Carica cubensis Solms using random amplified polymorphic DNA (RAPD) markers. Agricultural Sciences, 2010, vol. 1, pp. 95-101.
5. Campostrini, E. y Glenn, D. M. Ecophysiology of papaya: a review. Brazilian Journal of Plant Physiology, 2007, vol. 19, pp. 413-424.
6. Mentaberry, A. Tolerancia a estreses abióticos. [Consultado: 12 de noviembre de 2012] Disponible en:
http://www.fbmc.fcen.uba.ar/materias/agbt/teoricos/2011_15%20Resistencia%20a%20estres%20abiotico.pdf.
7. Marler, T. y Clemente, H. Papaya seedling growth responses to wind and water deficit is additive. HortScience, 2006, vol. 41, pp. 96-98.
8. Aiyelaagbe, I.; Fawusi, M. y Babalola, O. Growth, development and yield of pawpaw (Carica papaya L.). Homestead selection in response to soil moisture stress. Plant and Soil, 1986, vol. 93, pp. 427-435.
9. Mahouachi, J.; Arbona, V. y Gómez-Cadenas, A. Hormonal changes in papaya seedlings subjected to progressive water stress and re-watering. Plant Growth Regulation, 2007, vol. 53, pp. 43-51.
10. Ivonne, A. y Miranda, D. Efecto de cinco sustratos sobre índices de crecimiento de plantas de papaya (Carica papaya L.) bajo invernadero. Revista Colombiana de Ciencias Hortícolas, 2007, vol. 1, pp. 22-27.
11. Hernández, A.; Pérez, J. M.; Bosch, D. y Rivero, L. Nueva versión de la clasificación genética de los suelos de Cuba. La Habana: Instituto de Suelos. MINAGRI. 1999. 64 pp.
12. Mahouachi, J.; Argamasilla, Rosa y Gómez-Cadenas, A. Influence of Exogenous Glycine Betaine and Abscisic Acid on Papaya in Responses to Water-deficit Stress. Journal Plant Growth Regulation, 2012, vol. 31, pp. 1-10.
13. Masri, M.; Razak, A. S. y Ghazalli, M. Z. Response of papaya (Carica papaya L.) to limited soil moisture at reproductive stage. MARDI Research Journal, 1990, vol. 18, pp. 191-196.
14. Nava, A.; Ramírez, I.; Peña, C.; Díaz, G. y González, V. Características del intercambio de gases en hojas de guayabo (Psidium guajava L.). Revista Chapingo Serie Horticultura, 2009, vol. 15, pp. 119-126.
15. González, M. y Peters, J. Strategies of leaf expansion in Ficus carica under semiarid conditions. Plant Biology, 2009, vol. 12, pp. 469-474.
16. Almodares, A.; Taheri, R. y Adeli, S. Inter-relationship between growth analysis and carbohydrate contents of sweet sorghum cultivars and lines. Journal of Environmental Biology, 2007, vol. 28, pp. 527-531.
17. Alok, T.; Srivastava, N. y Khalid, K. Effect of Vesicular Arbuscular Mycorrhizal Fungi on Papaya (Carica papaya L.) Plant Under water Stress Conditions. Indian Journal of Scientific Research, 2011, vol. 2, pp. 65- 67.
18. Appels, A. y Lagudah, H. E. Manipulation of chromosomal segments from wild wheat for the improvement of bread wheat. Aust. J. Plant Physiol, 1990, vol. 17, pp. 253-266.
19. Fang, X.; Turner, N.; Li, F.; Li, W. y Guo, X. Caragana korshinskii seedlings maintain positive photosynthesis during short-term, severe drought stress. Photosynthetica, 2011, vol. 49, pp. 603-609.
20. Assumpção, F.; Campostrini, E.; Torres, A. y Hespanhol, L. Relationship between photochemical efficiency (JIP-Test Parameters) and portable chlorophyll meter readings in papaya plants. Brazilian Journal Plant Physiology, 2012, vol. 23, pp. 295-304.
21. Guo, P.; Baum, M.; Grando, S.; Ceccarelli, S.; Bai, G.; Li, R.; Von Korff, M.; Graner, A. y Valkoun, Jan. Differentially expressed genes between drought-tolerant and drought-sensitive barley genotypes in response to drought stress during the reproductive stage. Journal of Experimental Botany, 2009, vol. 60, pp. 3531-3544.
Recibido: 26 de diciembre de 2012
Aceptado: 31 de julio de 2013
Yusnier Díaz Hernández, Reserva Científica del Departamento de Fisiología y Bioquímica Vegetal. Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32 700. Email: yusnier@inca.edu.cu

{kind=link}
{kind=link}
{kind=link}