










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión On-line ISSN 1819-4087
cultrop vol.36 no.2 La Habana abr.-jun. 2015
Indicadores del crecimiento inicial y del estado nutricional para la selección temprana de genotipos de arroz (Oryza sativa L.) tolerantes a la salinidad
Indicators of initial growth and nutritional status for early selection genotypes of rice (Oryza sativa L.) salt tolerant
Alexis Lamz Piedra, Dra.C. María C. González Cepero
Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700.
RESUMEN
Con el objetivo de determinar posibles indicadores del crecimiento inicial y nutricionales para la selección temprana de genotipos de arroz tolerantes a la salinidad, se desarrollaron ensayos con dos cultivares con diferente grado de tolerancia a la salinidad ('Pokkali' y 'Amistad- 82') en condiciones controladas, con soluciones salinas ajustadas a conductividades eléctricas (CE) de 8 y 12 dSm-1 mediante la adición de NaCl y un control con agua destilada. Se evaluó el porcentaje de germinación a los 15 días después de la siembra (DDS) y los parámetros del crecimiento altura de la planta, longitud de la raíz, masa seca de la parte aérea (MSPA), masa seca del sistema radical (MSSR) a los 21 DDS. En tanto, en hojas y raíces de ambos cultivares, en condiciones semicontroladas y utilizando soluciones nutritivas para la simulación del estrés salino, con CE de 8 y 12 dSm-1, se determinó en estado de plántula, el contenido de nitrógeno (N), fósforo (P), potasio (K), sodio (Na), así como la relación sodio/potasio. Los resultados evidencian un comportamiento diferencial de los cultivares en cuanto a su grado de tolerancia a la salinidad, lo que permite usar algunas de las variables evaluadas, como indicadores para la discriminación de genotipos de arroz tolerantes a salinidad.
Palabras clave: arroz, salinidad, crecimiento, indicadores.
ABSTRACT
In order to identify possible indicators of initial growth and nutrition for early selection of rice genotypes tolerant to salinity, tests with two cultivars with different degree of salinity tolerance ('Pokkali' and 'Amistad-82') were developed under controlled conditions, saline solutions tailored to electrical conductivities (EC) of 8 and 12 dSm-1 by the addition of NaCl and a control with distilled water. The percentage of germination at 15 days after sowing (DAS) and the growth parameters: plant height, root length, dry mass of the aerial part (MSPA), dry mass of the root system (MSSR) to 21 DAS were evaluates. Meanwhile, in leaves and roots of both cultivars under controlled conditions and using nutrient solutions for the simulation of salt stress (CE 8 and 12 dSm-1), was determined at seedling stage, the content of nitrogen (N), phosphorus ( P ) , potassium ( K ), sodium ( Na ) , and sodium / potassium ratio. The results show a differential behavior of the cultivars in their degree of tolerance to salinity, allowing use some of the variables evaluated as indicators for discrimination of rice genotypes tolerant to salinity.
Key words: rice, salinity, growth, indicators.
INTRODUCCIÓN
El área de nuestro planeta, afectada por la salinización, es de alrededor de 800 millones de hectáreas, lo que la convierte en uno de los problemas más serios que enfrenta la agricultura a nivel mundial (1, 2, 3), de lo cual Cuba no está exenta, presentando alrededor de 7,08 millones de hectáreas como superficie agrícola, de las que, un millón de hectáreas están afectadas, y 1,5 millones presentan problemas potenciales de salinización. Esto representa para nuestro país, alrededor de un 14 % de áreas afectadas con diferentes grados de salinización (4).
El exceso de iones salinos en la solución de suelo, provoca daños que implican una afectación en el crecimiento y desarrollo de los cultivos y, por tanto, una disminución de su potencial de rendimiento (5).
Estos daños son el resultado de los trastornos provocados en el metabolismo de las plantas, principalmente por los cambios en el potencial osmótico del suelo, el desbalance nutricional por la interacción entre los iones tóxicos y los nutrientes esenciales para el crecimiento y desarrollo, así como el estrés oxidativo, inducido a partir de los diferentes efectos por el aumento de las especies reactivas de oxigeno (ERO) en los componentes celulares (5).
En la política de recuperación y manejo de estos suelos, el uso de especies y variedades tolerantes al estrés salino se considera como el acontecimiento más pragmático para obtener buenos rendimientos en condiciones salinas (6) y ha recibido especial atención por diferentes investigadores en Cuba y en diversas regiones del mundo (5, 7). A pesar de que esta temática ha sido objeto de múltiples investigaciones, particularmente en los últimos 30 años, aún no se cumplen todas las expectativas y se tendrá que seguir trabajando en la mejora genética de los diferentes cultivos para enfrentar este problema.
El arroz es uno de los cultivos de mayor importancia económica a nivel mundial. Este constituye uno de los cereales de mayor consumo para más de la mitad de la población, después del trigo (8). Sin embargo, su productividad es seriamente afectada por la salinidad en cada etapa del desarrollo.
Por lo anteriormente planteado, el mejoramiento para la tolerancia a la salinidad en el cultivo del arroz, requiere de técnicas de evaluación de los diferentes materiales, las que deben ser rápidas para mantener un gradiente de evaluación correspondiente con la gran cantidad de material que se mejora. Se ha planteado que la selección en estadios tempranos del crecimiento para el desarrollo de nuevas variedades de arroz tolerantes a la salinidad, proporciona una evaluación rápida, que resulta difícil en estados vegetativos avanzados o en estado reproductivo (9) y contribuye a disminuir el periodo de obtención de los nuevos cultivares, a la disminución del número de materiales para las próximas etapas del mejoramiento, donde sería engorroso trabajar con grandes volúmenes de genotipos y reduce otros efectos medioambientales, debido a que se trabaja en condiciones controladas o semicontroladas.
El éxito de cada una de las metodologías de selección y evaluación de nuevos materiales en un programa de mejora genética, está relacionado con la definición de los objetivosA, así como la determinación de los indicadores viables para la selección de genotipos con un comportamiento adecuado en estas condiciones desfavorables, constituyen herramientas de gran importancia (9). Por ello, ha sido objeto de estudio la variación de parámetros bioquímicos, ya que, en alguna medida, se conocen las bases bioquímicas de la tolerancia y cuáles de estas pudieran constituir mecanismos adaptativos que se inducen en las plantas y que, a su vez, presenten una estrecha vinculación con el estrés (10). La necesidad de contar con indicadores para la selección temprana de genotipos de arroz tolerantes a la salinidad impone a los investigadores, fitomejoradores del cultivo del arroz, nuevos retos en los cuales tienen que desarrollarse. De esta manera se estaría contribuyendo a la utilización de zonas afectadas con problemas de salinidad en el territorio nacional. El presente trabajo tuvo como objetivo determinar indicadores del crecimiento inicial y del estado nutricional en la fase vegetativa de plántulas, para la selección temprana de genotipos de arroz tolerantes a la salinidad.
MATERIALES Y MÉTODOS
En el estudio fueron utilizadas semillas de dos cultivares de arroz con diferentes respuestas a la salinidad: Pokkali testigo internacional tolerante a la salinidad (7) y Amistad 82 (A-82) testigo susceptible a la salinidad (9). Las semillas fueron desinfestadas con hipoclorito de sodio al 3 % durante cinco minutos; posteriormente, se lavaron tres veces con agua destilada.
Para los indicadores del crecimiento inicial, se colocaron 50 semillas en placas (10,5 x 0,8 cm) que tenían papel de filtro como soporte a las que se le aplicó 10 mL de soluciones salinas empleándose NaCl hasta obtener CE de 8 y 12 dSm-1, según los tratamientos. Al tratamiento control se le aplicó solo agua destilada. Se sembraron tres placas por tratamiento, las que se colocaron en condiciones de oscuridad por 48 horas para propiciar la germinación. Posteriormente, se mantuvieron a una temperatura de 25± 2 °C y un fotoperíodo de 16 h de luz y 8 h de oscuridad, en un diseño completamente aleatorizado. El experimento se repitió dos veces en el tiempo en iguales condiciones de trabajo.
A los quince días después de la siembra (DDS) se contaron las semillas germinadas y se determinó el porcentaje de germinación y a los 21 DDS se evaluó la altura de la planta (AP), la longitud de la raíz (LR), la masa seca de la parte aérea (MSPA) y del sistema radical (MSSR), así como se determinó la relación entre la parte aérea y el sistema radical (PA/SR).
Para los indicadores nutricionales, tres semillas pregerminadas se trasplantaron en recipientes plásticos (200 mL) que contenían suelo Hidromórfico Gley Nodular Ferruginoso (11) y fueron colocadas en cajas que contenían agua destilada. El experimento se desarrolló en condiciones de invernadero. Quince días después del trasplante (DDT), se les indujo el estrés salino mediante la aplicación de solución nutritiva Murashige y Skoog al 50 % (MS ½) (12) ajustadas a CE de 8 y 12 dSm-1 obtenidas por la adición de NaCl y un control con solución nutritiva MS ½.
Quince días después de impuesto el estrés salino, se tomaron muestras foliares y del sistema radical de tres recipientes de cada cultivar para determinar el porcentaje de N por determinación colorimétrica con el reactivo de Nessler, el porcentaje de P por determinación colorimétrica de la formación del color azul del complejo molibdato-fosfórico y el porcentaje K+ y Na+ por fotometría de llamas (13). Con los valores del K+ y Na+ se determinó la relación Na+/ K+. Se empleó un diseño completamente aleatorizado con seis repeticiones y el experimento se repitió dos veces en el tiempo en iguales condiciones de trabajo.
Los datos se procesaron mediante análisis de varianza simple, basado en un modelo lineal de efectos fijos, utilizando el paquete estadístico SPSS versión 11.5 y las diferencias significativas entre tratamientos se verificaron por la prueba de rango múltiple de Tukey para un nivel de significación del 5 %.
RESULTADOS Y DISCUSIÓN
Determinación de indicadores morfofisiológicos durante la germinación y estados tempranos de crecimiento
Los resultados revelaron un comportamiento diferente de los genotipos estudiados en cuanto a la germinación de las semillas en las diferentes concentraciones salinas evaluadas (Figura 1).
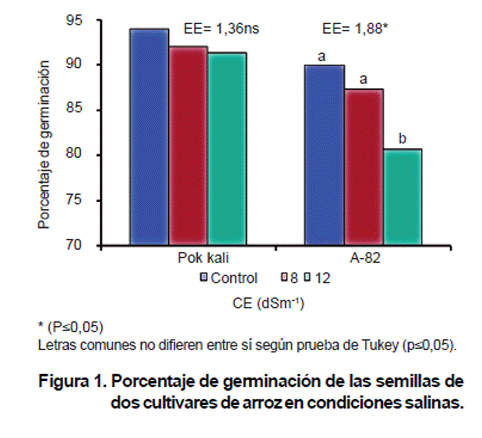
Se pudo apreciar una disminución de la germinación de las semillas en el cultivar A-82 a medida que aumentó la CE del medio de crecimiento, mostrando diferencias significativas (p=0,05) respecto al control a CE de 12 dSm-1, no siendo así para la CE de 8 dSm-1.
El cultivar Pokkali no mostró diferencias significativas (p=0,05) en cuanto a la germinación de las semillas para ninguna de las CE estudiadas. En este caso, el buen comportamiento de la germinación de las semillas en Pokkali, le ofrece ventajas para un mejor establecimiento, crecimiento y desarrollo efectivo de sus plántulas en condiciones de estrés salino.
El comportamiento del cultivar Pokkali, puede estar asociado a que en estas condiciones, pudo mantener la absorción de agua a través del tegumento, por lo que los procesos físicos como la imbibición de las semillas, así como la hidrólisis de los polisacáridos de reserva no se ven afectados, lo que proporciona una buena germinación.
La disminución de la germinación de las semillas en el cultivar A-82 en condiciones salinas, se puede atribuir a una menor absorción de agua que pudo afectar la imbibición de estas, ya que la imbibición es uno de los primeros eventos que ocurre durante la germinación de las semillasB.
El comportamiento diferente observado entre ambos cultivares en CE de 12 dSm-1, permite discriminar entre genotipos, convirtiéndose la germinación de las semillas en un indicador eficiente para la selección temprana.
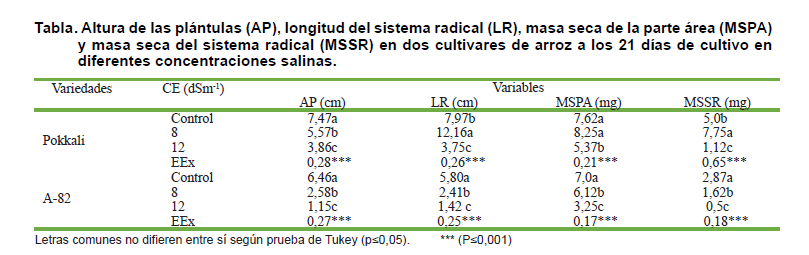
De forma general, los parámetros del crecimiento inicial evaluados, se vieron influenciados por el estrés salino en ambos cultivares (tabla). A pesar de la influencia del estrés, un comportamiento diferente de los cultivares se detectó en la CE de 8 dSm-1 en la longitud de la raíz y la acumulación de masa seca del sistema radical.
En el cultivar Pokkali, se evidenció una estimulación significativa (p=0,05) respecto a su control de LR y de MSSR, en la CE de 8 dSm-1, mientras que el cultivar A-82, mostró una reducción significativa de la LR y MSSR. La masa seca de la parte aérea no mostró variaciones significativas en el cultivar Pokkali en CE de 8 dSm-1 respecto a su control, reduciéndose en CE 12 dSm-1; sin embargo, en A.82 la masa seca de la parte aérea fue más afectada, ya que disminuyó con el aumento de CE en el medio de crecimiento. Estos parámetros pueden ser indicadores para la selección de genotipos tolerantes al estrés salino en la etapa inicial del crecimiento en el cultivo del arroz.
La estimulación de la LR detectada en el cultivar tolerante (Pokkali) es una característica importante de este cultivar. Esta respuesta por lo general, se atribuye a la necesidad de las plantas de absorber agua en condiciones de elevada salinidad y sequía, lo que provoca un alargamiento del sistema radical.
Un sistema radical más alargado en condiciones de estrés osmótico, que puede ser provocado, tanto por falta de agua como por una elevada concentración de sales en el medio de crecimiento, pudiera contribuir a la búsqueda de agua, así como a un mejor desarrollo en estas condiciones de crecimiento. Por otro lado, la menor longitud de la raíz en A-82 puede ser el resultado de la pérdida de turgencia celular (5).
Las disminuciones en la acumulación de biomasa en condiciones salinas, se ha evidenciado en una amplia gama de cultivos (14, 15, 16). El aumento de la MSSR que manifestó el cultivar Pokkali, puede estar relacionado con el aumento de la longitud del sistema radical (tabla), así como, con la tolerancia de este cultivar al estrés salino, ya que un sistema radical más desarrollado en estas condiciones puede mejorar la absorción de agua. El incremento en la absorción de agua cuando existe un desarrollo mayor del sistema radical pudiera contribuir a mantener la actividad fotosintética expresada en masa secaB.
Se detectó que el estrés salino tuvo una influencia importante en la reducción de la altura de las plantas de ambos cultivares (tabla). La influencia del estrés salino sobre la altura de las plantas, no permitió detectar comportamiento diferente de los cultivares evaluados, por lo tanto, no se considera un indicador eficiente para la selección de genotipos de arroz tolerantes a la salinidad en estadíos tempranos del crecimiento.
Las disminuciones en la altura de las plántulas es el resultado de una pérdida de turgencia a nivel celular, provocada por la disminución del potencial osmótico en el medio de crecimiento de los cultivares. La presión de turgencia en las plantas es indispensable para el crecimiento celular en expansión y para la apertura estomática. La presencia de sales en el suelo disminuye el potencial osmótico, causando un estrés hídrico en plantas que lleva a una pérdida de dicha turgencia con su considerable efecto en el crecimientoC.
Indicadores nutricionales (N, P, K+, Na+ y Na+/ K+)
Se pudo apreciar un comportamiento diferente de los cultivares al analizar su estado nutricional según su grado de tolerancia al estrés salino.
En la Figura 2 se puede apreciar el porcentaje de nitrógeno en la raíz de ambos cultivares de arroz. Resultó interesante el resultado encontrado en relación con la no influencia de la salinidad sobre el nitrógeno en hojas, lo que pudiera indicar que el aumento de las concentraciones salinas, no afectan en gran medida la translocación de este macro-nutriente a la parte aérea en el cultivo del arroz. Sin embargo, para el sistema radical, un aumento significativo del contenido de N se aprecia en el cultivar Pokkali en CE de 12 dSm-1, mientas que en el cultivar A-82 el aumento fue en la CE de 8 dSm-1. Esto indica una respuesta diferente de los cultivares en función de la tolerancia a la salinidad.
Las diferencias significativas (p=0,05) del contenido de N observadas en la CE de 12 dSm-1 en la raíz del cultivar Pokkali respeto a su control, demuestran que este cultivar mantiene una mejor absorción de este nutriente para las condiciones evaluadas, lo que ratifica su mejor comportamiento ante el estrés salino respecto al cultivar A-82 por lo que el contenido de N en raíz en CE de 12 dSm-1 pudiera constituir un indicador eficiente para la selección de genotipos de arroz tolerantes a la salinidad en estado vegetativo de plántulas.
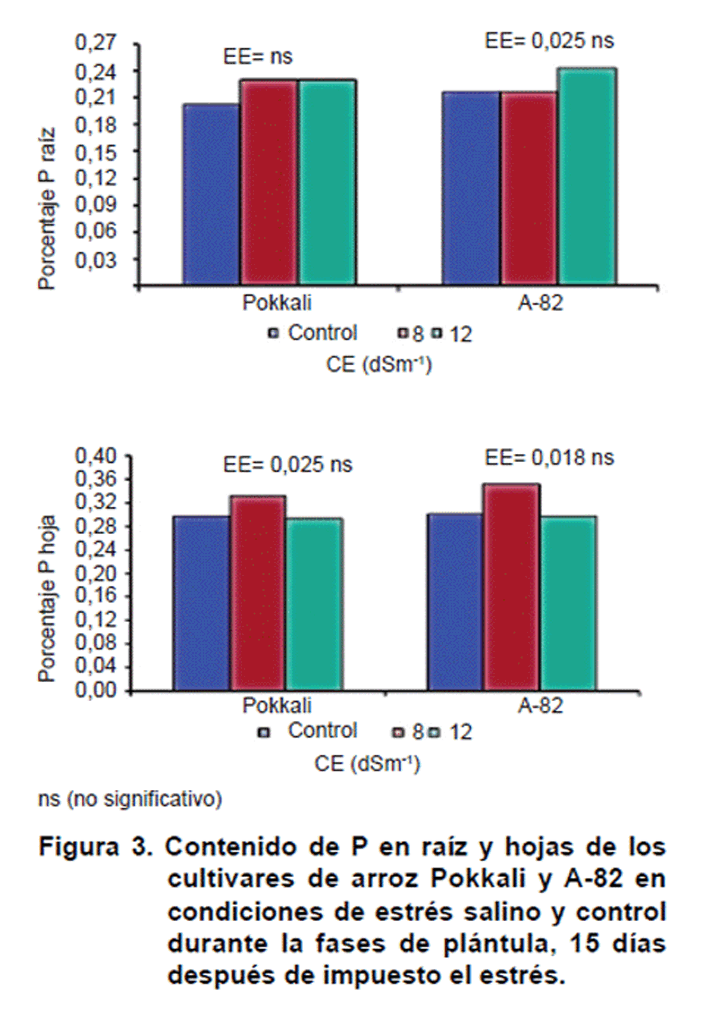
El contenido de fósforo (Figura 3) no manifestó variaciones significativas con el aumento de las concentraciones salinas (como las empleadas en el experimento), para los cultivares estudiados. Esto sugiere que este nutriente no se afecta en condiciones salinas para el cultivo del arroz.
Los resultados detectados, no concuerdan con lo reportado en otros cultivos de interés económico. En este sentido, en el cultivo del tomate se observaron disminuciones del contenido de P con el aumento de las concentraciones salinas en el medio de crecimientoD.
Por otra parte, se detectó un comportamiento semejante al encontrado en este trabajo en el cultivo del arroz, al plantear que el contenido de P no varió en hojas con el aumento de las concentraciones salinas (17).
Teniendo en cuenta el comportamiento de los cultivares en cuanto al contenido de P, se puede asumir que este no constituye un indicador discriminante entre cultivares de arroz tolerantes y susceptibles a la salinidad.
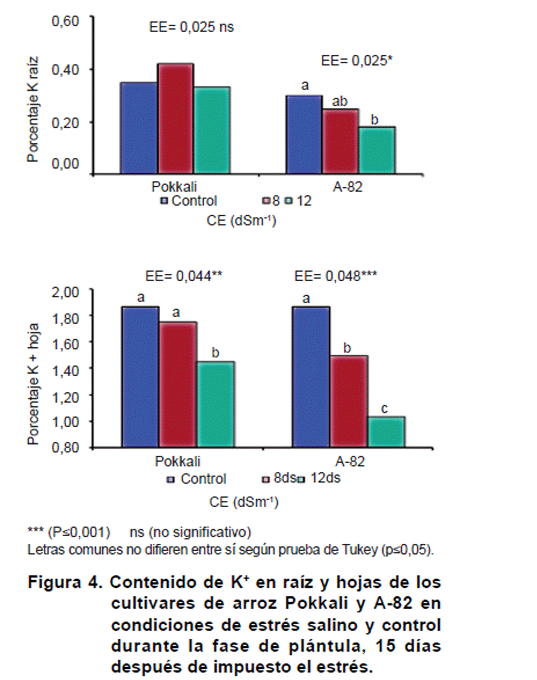
Tanto en hojas como en raíces una disminución del contenido de potasio fue visible con el aumento de las concentraciones salinas en el medio de crecimiento en el cultivar A-82. Sin embargo, el cultivar Pokkali no manifestó reducciones significativas (p=0,05) del contenido de K+ en su raíz (Figura 4).
El contenido de K+ fue más afectado en A-82, observándose diferencias estadísticas en hojas a partir de la CE de 8 dSm-1 y raíz en CE de 12 dSm-1. En el cultivar Pokkali solo se detectó un efecto del estrés salino en hojas a CE de 12 dSm-1, siendo este un relevante resultado debido a la importancia del potasio en la tolerancia a la salinidad, evidenciándose un comportamiento diferencial entre ambos cultivares.
La traslocación del potasio a la parte aérea, también manifestó reducciones con el aumento de la CE en el medio de crecimiento. No obstante, el cultivar Pokkali en hojas, manifestó menor reducción de este elemento que el cultivar A-82 a medida que aumentó la CE del medio de crecimiento.
Como soluto en las vacuolas, el K+ juega un importante rol en el control de las relaciones hídricas ayudando a la turgencia celular (14). Es bien conocida la importancia del mantenimiento de la turgencia en el crecimiento celular, por tanto, el crecimiento de las plantas puede ser inhibido por reducción de la concentración de K+ que reduce la capacidad de ajuste osmótico y el mantenimiento de la turgencia celular.
En este caso, el mejor mantenimiento del estado normal del contenido de potasio en Pokkali, tanto en la raíz como en la parte aérea, pudiera constituir un mecanismo de tolerancia al estrés salino, que contribuiría con el mantenimiento de la turgencia celular y del crecimiento en condiciones salinas. En este sentido se sostiene que la tolerancia de las plantas a la salinidad es atribuida a su habilidad para evitar toxicidad por Na+ y al mantenimiento de las concentraciones de K+ (14). Por lo anteriormente planteado, el contenido de potasio en hoja y raíz de cultivares de arroz en un diseño de estudios similar, constituye un indicador eficiente para la selección de genotipos tolerantes a la salinidad.
En cuanto al sodio (Figura 5), se pudo apreciar un aumento significativo (p=0,05) de su contenido, tanto en hojas como en el sistema radical de ambos cultivares a medida que aumentó la CE en el medio de crecimiento. El cultivar Pokkali en estas condiciones acumuló menor contenido de Na+ en raíz y hojas, así como translocó menos contenido de este ión toxico al área foliar que el cultivar A-82. Sin embargo, no se detectó comportamiento diferente de los cultivares debido a que, tanto Pokkali como A-82 difieren significativamente de su control correspondiente en CE de 8 y 12 dSm-1.
La acumulación de Na+ en tejido vegetal en un medio de crecimiento salino se atribuye a una disminución de la integralidad de la membrana celular, debido a la sustitución del Ca2+ por el Na+ que influye directamente en sus funciones biológicas (5).
En el presente trabajo, el contenido de Na+ en tejido vegetal, no permite la discriminación entre cultivares de arroz sensibles y tolerantes al estrés salino, por lo que no constituye un indicador eficiente para la selección de genotipos tolerantes a la salinidad.
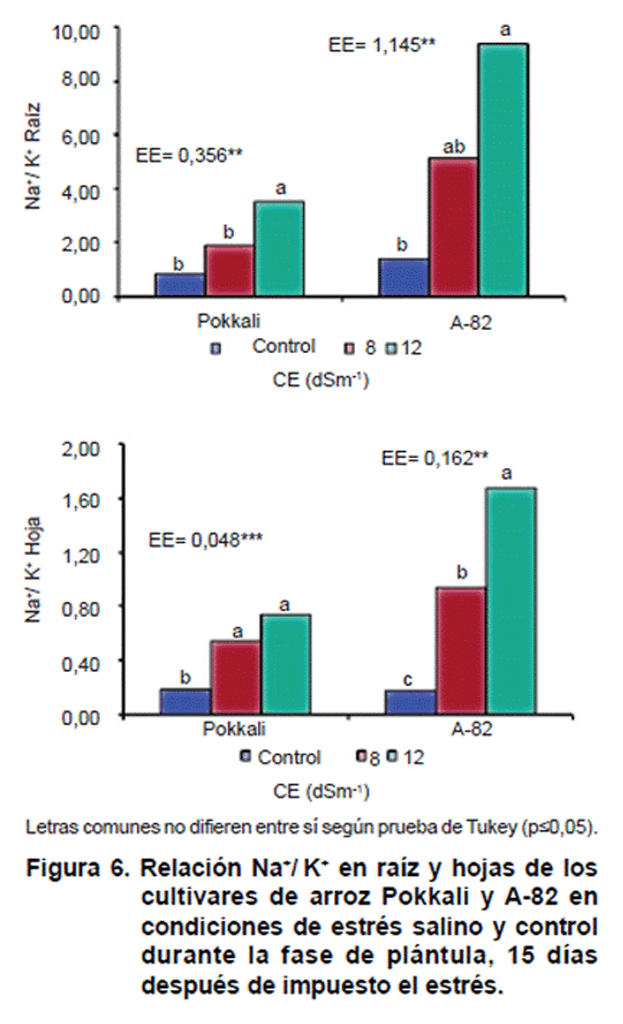
La relación Na+/K+, se afectó a medida que aumentó la CE en el medio de crecimiento para ambos cultivares, siendo más marcada en el sistema radical respecto al área foliar para ambos cultivares, observándose un aumento significativo (p=0,05) de la misma (Figura 6). El comportamiento diferencial entre los cultivares fue apreciable, ya que el cultivar Pokkali mantuvo menor relación Na+/K+ que A-82 con el aumento de la CE del medio, lo cual está dado por una menor absorción de Na+, así como un mejor mantenimiento de la absorción de K+. Esto sugiere que Pokkali es capaz de mantener una adecuada absorción de K+ en medio salino.
En este caso, en el cultivar Pokkali se apreciaron diferencias significativas (p=0,05) de la relación Na+/K+ en hojas en la CE de 8 dSm-1, pero esta no difiere del la CE de 12 dSm-1. Sin embargo, en hojas de A-82 las diferencias estadísticas se observaron en 8 y 12 dSm-1 respecto a su control.
En la raíz, la relación Na+/K+ aumentó en menor medida en el cultivar Pokkali y se detectaron diferencias estadísticamente significativas en la CE de 12 dSm-1, mientras que A-82, la relación Na+/K+ manifestó tendencia al aumento desde la CE de 8 dSm-1 que difiere de su control en CE de 12 dSm-1.
Resultados similares han sido observados en cuanto a la relación Na+/K+ en otros cultivos de interés económico (14). El aumento en relación Na+/K a menudo se atribuye al efecto de competencia entre los iones Na+ y K+ en el medio de desarrollo de la raíz (18). La menor relación Na+/K+ en Pokkali está altamente relacionada con su tolerancia al estrés salino. La tolerancia al estrés por salinidad en plantas es atribuida a su habilidad de evitar la toxicidad por Na+ y al mantenimiento de la concentración de Ca2+ y K+ (14).
CONCLUSIONES
Se concluye, que los cambios ocasionados por el estrés salino en la fase inicial del crecimiento y en el estado nutricional en la fase de plántulas permitió determinar indicadores discriminantes entre los genotipos evaluados. Tal es el caso del porcentaje de germinación, LR, MSPA, MSSR, el contenido de N en raíz, el contenido de K+ en raíz y hojas y la relación Na+/K+. Estos pueden ser indicadores eficientes para la selección temprana de genotipos de arroz tolerantes a la salinidad, los que pueden ser incorporados en los programas nacionales de mejoramiento genético de este cultivo.
BIBLIOGRAFÍA
1. FAO. FAO Land and Plant Nutrition Management Service. 2010. [en línea]. Disponible en: <http://www.fao.org/ag/agl/agll/spush> .
2. Sheng, M.; Tang, M.; Zhang, F. y Huang, Y. Influence of arbuscular mycorrhiza on organic solutes in maize leaves under salt stress. Mycorrhiza, 2011, vol. 21, pp. 423-430. ISSN: 1432-1890.
3. Azad, I.; Branch, B. y Iran, B. The Effects of Salt stress on plant growth. Technical Journal of Engineering and Applied Sciences, 2012, vol. 2, no.1, pp. 7-10. ISSN: 2051-0853.
4. ONE. Anuario Estadístico Cuba. Edición 2011. [en línea]. [Consultado: 14-11-2012]. Disponible en: <http://www.one.cu/PublicacionesDigitales/FichaPublicacion.asp?CodPublicacion=23&CodDireccion=2> .
5. Nawaz, K.; Hussain, K.; Majeed, A.; Khan, F., Afghan, S. y Ali, K. Fatality of salt stress to plants: Morphological, physiological and biochemical aspects. African Journal of Biotechnology, 2010, vol. 9, no. 34, pp. 5475-5480. ISSN: 1684-5315.
6. Kumar, S.; Titov, S.; Mofazzal, M.; Siddika, A.; Sultana, A. y Shahidul, M. D. Phenotypic and genotypic screening of rice genotypes at seedling stage for salt tolerance. African Journal of Biotechnology, 2009, vol. 8, no. 23, pp. 6490-6494. ISSN: 1684-5315.
7. Argentel, L., López, R. D., González, L. M., López, R. C., Gómez, E. y Fonseca, I. Evaluación de la tolerancia a la salinidad en estadios tempranos y finales del desarrollo en triticales (X Triticum secale). Cultivos Tropicales, 2010, vol. 31, no. 1, pp. 48-53. ISSN: 1819-4087.
8. Ali, Y.; Aslam, Z.; Awan, A. R.; Hussain, F. y Cheema, A. A. Screening rice (Oryza sativa L.) lines/cultivars against salinity in relation to morphological and physiological traits and yield components. International Journal Of Agriculture & Biology, 2004, vol. 6, no. 3, pp. 572-575. ISSN: 1684-5315.
9. Cristo, E.; González, M. C.; Cárdenas, R. M. y Pérez, N. Evaluación de la tolerancia a la salinidad en el estado juvenil de tres nuevas líneas de arroz (Oryza sativa L.) utilizando marcadores morfoagronómicos. Cultivos Tropicales, 2001, vol. 22, no. 2, pp. 43-45. ISSN: 1819-4087.
10. Lamz, A., González, M. C. y Reyes, Y. Indicadores bioquímicos para la selección temprana de genotipos de arroz (Oryza sativa L.) con tolerancia a la salinidad. Cultivos Tropicales, 2013, vol. 34, no. 1, pp. 10-17. ISSN: 1819-4087.
11. Minagri. Instituto de Suelos. Nueva Versión de Clasificación Genética de los Suelos de Cuba. Instituto de Suelos. 1999, 64 pp.
12. Murashige, T. y Skoog, F. A revised medium for rapid growth and bioassay with tobacco tissue culture. Plant Physiol., 1962, vol. 13, no. 3, pp. 473-479. ISSN: 0019-5502.
13. Paneque, V.; Calaña, J. M.; Calderón, M.; Borges, Y.; Hernández, T. C. y Caruncho, M. Manual de técnicas analíticas para análisis de suelo, foliar, abonos orgánicos y fertilizantes químicos. San José de las Lajas, La Habana, Cuba: Instituto Nacional de Ciencias Agrícolas, 2010. 156 pp.
14. Amirjani, M. R. Effect of salinity stress on growth, mineral composition, proline content, antioxidant enzymes of soybean. American Journal of Plant Physiology, 2010, vol. 5, no. 6, pp. 350-360. ISSN: 1557-4539.
15. Bibi, A.; Ellahi, N.; Ali, A.; Hussain, F.; Hussain, N. y Ahmad, M. Nutritional influence of salt stress on the growth and nodule formation of Vicia faba. Pakistan Journal of Nutrition, 2012, vol. 11, no. 7, pp. 562-567. ISSN: 1680-5194.
16. Beltrano, J.; Ruscitti1, M.; Arango, M. C. y Ronco, M. Effects of arbuscular mycorrhiza inoculation on plant growth, biological and physiological parameters and mineral nutrition in pepper grown under different salinity and p levels. Journal of Soil Science and Plant Nutrition, 2013, vol. 13, no. 1, pp. 123-141. ISSN: 0718-9508.
17. Naheed, G. Shahbaz, M. y Akram, N. A. Interactive effect of rooting medium application of phosphorus and nacl on plant biomass and mineral nutrients of rice (Oryza sativa L.). Pak. J. Bot., 2008, vol. 40, no. 4, pp. 1601-1608. ISSN: 2070-3368.
18. Heidari, M. y Jamshid, P. Interaction between salinity and potassium on grain yield, carbohydrate content and nutrient uptake in pearl millet. ARPN Journal of Agricultural and Biological Science, 2010, vol. 5, no. 6, pp. 39-46. ISSN: 1990-6145.
Recibido: 24 de abril de 2014
Aceptado: 3 de septiembre de 2014
Alexis Lamz Piedra, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700. Email: alamz@inca.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}