










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión On-line ISSN 1561-3054
Rev Cubana Med Trop vol.50 supl.1 Ciudad de la Habana 1998
ARTÍCULOS ORIGINALES
INSTITUTO DE MEDICINA TROPICAL "PEDRO KOURÍ"Características antigénicas de los virus aislados del líquido cefalorraquídeo de pacientes con neuropatía epidémica
Lic. Luis Sarmiento Pérez,1 Dr. Pedro Más Lago,2 Dra. Ivonne Ávalos Redón,3 Lic. Marité Bello Corredor4 y Téc. Rosa Palomera Puentes5RESUMEN
Durante la epidemia de neuropatía ocurrida en Cuba en los años 1992/1994 se aislaron del líquido cefalorraquídeo de pacientes agentes virales relacionados antigénicamente con los virus Coxsackie. Para establecer una función de estos virus en la etiopatogenia de la enfermedad se seleccionaron las cepas 47/93 IPK identificada como Coxsackie A9 y la cepa 44/93 IPK de efecto citopático ligero (ECP-L) y se realizó un estudio de sus características antigénicas mediante Western Blot. Se comprobó la relación antigénica entre ambas cepas y se demostró la ausencia de proteínas estructurales en su forma nativa en los agentes de ECP-L. A partir de estos resultados se plantea la posibilidad de que la persistencia sea un mecanismo por el cual estos virus participen en la etiopatogenia de la neuropatía epidémica en Cuba.Descriptores DeCS: NEURITIS/ líquido cefalorraquídeo; NEURITIS/ etiología; VIRUS COXSACKIE/ aislamiento & purificación; CUBA.
Aunque determinados agentes biológicos han sido reportados como causa probable de neuropatías epidémicas,1 el primer reporte en que se encontraron virus en el líquido cefalorraquídeo (LCR) de pacientes con neuropatía tropical ocurrió durante la epidemia de Japón en 1971, pero la función de estos virus nunca se aclaró.2
La epidemia de neuropatía ocurrida en Cuba, constituye la segunda evidencia de la presencia de virus en el LCR de pacientes con neuropatía epidémica, pues de éste se aislaron agentes virales que mostraron 2 tipos de efectos citopatogénicos: uno típico de Enterovirus, en un reducido número de casos, que se identificó como Coxsackievirus A9; y otro de progresión lenta y ligera (efecto citopático ligero: ECP-L) en la mayoría de los pacientes, cuyo agente responsable no se pudo identificar pero por prueba de neutralización se relacionó antigénicamente con los Coxsackievirus A9 y B4.3
Aunque los patrones epidemiológico e inmunológico encontrados,4-6 no apoyan una infección aguda con las características clásicamente conocidas y por tanto, muestran poca evidencia de que la enfermedad tuviera carácter infeccioso, el alto porcentaje de aislamientos encontrados en los LCR de los pacientes, así como el obtener aislamientos en 2 sistemas independientes,3 sugieren que estos agentes virales deben participar en la etiopatología de la neuropatía epidémica en Cuba.
Teniendo en consideración estos hallazgos, nos propusimos realizar un estudio antigénico de los agentes virales aislados durante la epidemia mediante Western Blot con los objetivos de profundizar en las características antigénicas de éstos y determinar posibles mecanismos por los cuales estos virus participan en la etiopatogenia de la neuropatía epidémica en Cuba.
MÉTODOS
Antígenos. Para este estudio se seleccionaron la cepa 47/93 IPK identificada como Coxsackievirus A9 y la cepa 44/93 IPK de ECP-L y se procedió a la preparación de los antígenos virales mediante el método descrito por Mertenz y otros.7 Además, se preparó un antígeno control de células Vero no inoculadas utilizando esta misma metodología.Western-Blot. Los diferentes antígenos fueron sometidos a electroforesis en gel de poliacrilamida en presencia de duodecil sulfato de sodio.8 Se preparon 2 corridas electroforéticas con los antígenos agrupados de la forma siguiente:
- Corrida 1-Gel 1: antígeno 44/93 IPK, antígeno 47/93 IPK, antígeno Vero.
- Corrida 2-Gel 2: antígeno 44/93 IPK, antígeno 47/93 IPK, antígeno Vero.
| | | |
| 44/93 IPK | ||
| 1 | 47/93 IPK | Suero 44/93 IPK |
| | ||
| 44/93 IPK | ||
| 2 | 47/93 IPK | Suero 47/93 IPK |
| |
Al concluir esta incubación las membranas fueron lavadas e incubadas con el conjugado anti-Ig de conejo peroxidasa diluido 1/1 000 en PBS-T20-leche. Luego se realizaron lavados similares a los ya descritos y se procedió a revelar la reacción con el sustrato de la enzima.
RESULTADOS
Membrana 1. El suero hiperinmune a la cepa 44/93 IPK reconoció en el antígeno obtenido de la cepa 47/93 IPK (Cox A9) 2 proteínas que por su peso molecular de 30 y 20 kD, respectivamente, son compatibles con las proteínas estructurales VP1 y VP3 de los Enterovirus,mientras que con el antígeno homólogo se ponen de manifiesto proteínas de un peso molecular de 43 kD no compatibles con las proteínas estructurales de los Enterovirus (fig. 1).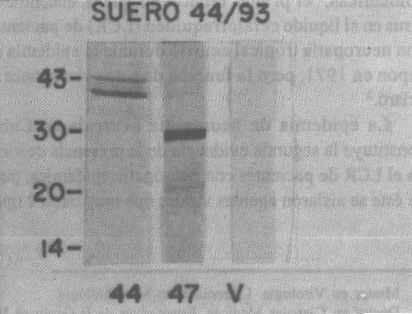
Fig. 1. Prueba de Western Blot con suero hiperinmune a la cepa 44/93 IPK.
44: Antígeno de cepa 44/93 IPK (ECP-L).
47: Antígeno de cepa 47/93 IPK (Cox A9).
V: Antígeno de células Vero.
Membrana 2. El suero hiperinmune a la cepa 47/93 IPK reconoció en su antígeno homólogo las proteínas estructurales VP1 y VP3 de 30 y 20 kD, respectivamente y no se obtuvo señal de reconocimiento con el antígeno obtenido de la cepa 44/93 IPK (fig.2).
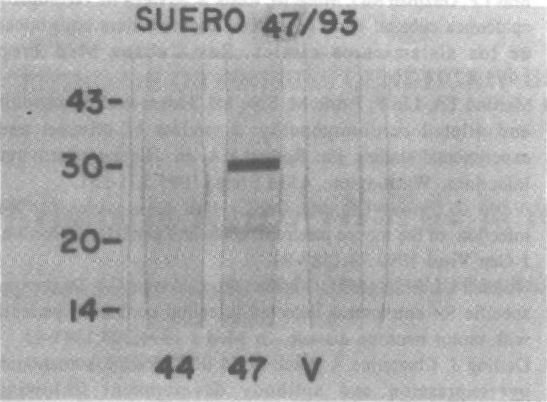
Fig. 2. Prueba de Western Blot con suero hiperinmune a la cepa 47/93 IPK.
44: Antígeno de cepa 44/03 IPK (ECP-L).
47: Antígeno de cepa 47/93 (Cox A9).
V: Antígeno de células Vero.
En ambas pruebas el antígeno preparado con las células Vero, utilizadas como control, mostró resultados negativos (figs. 1 y 2).
DISCUSIÓN
La reactividad cruzada del suero hiperinmune a la cepa 44/93 IPK con los antígenos virales demuestra la relación antigénica entre ambas cepas y concuerdan con los resultados obtenidos por microneutralización al caracterizar los aislamientos de ECP-L como Enterovirus relacionados antigénicamente con Cox A9 y Cox B4.3,9La ausencia de reconocimiento del antígeno preparado con las células Vero utilizadas para realizar los aislamientos permiten considerar que el reconocimiento de los sueros es virus específico y no producto de anticuerpos contra estructuras celulares. Confirman una vez más, que los aislamientos no constituían virus latentes en los sistemas celulares utilizados para obtenerlos.
La ausencia de reconocimiento del suero anti- -47/93 IPK a la cepa 44/93 IPK y la obtención, con su suero homólogo, de proteínas de peso molecular de 43 kD, no compatibles con las proteínas estructurales de los Enterovirus, se corresponde también con la prueba de neutralización cruzada.
De estos resultados no se puede inferir que exista deleción de la parte del genoma que codifica para las proteínas estructurales en la cepa 44/93 IPK, pero sugiere que en las proteínas estructurales de los agentes de ECP--L puedan existir modificaciones que repercutan en la estructura del virión y den lugar a una aparente falta de proteínas estructurales, por lo menos en su forma nativa. Estas modificaciones pueden conllevar a un cambio de los principales sitios antigénicos y de esta forma burlar el sistema inmune y establecer persistencia.10
Los Enterovirus, aunque de forma usual son líticos, bajo ciertas condiciones como en la presencia de un genoma viral defectivo o una replicación defectiva pueden causar infección persistente. La persistencia viral es una hipótesis reciente y altamente atractiva que explica cómo los Enterovirus o la interacción virus-inmunidad pueden causar daños en los tejidos u órganos, en particular durante la fase crónica de muchas enfermedades.11
Varios autores han demostrado este mecanismo en la persistencia de Enterovirus en algunas enfermedades crónicas. Woodall y otros,12 encontraron por análisis de la secuencia de la región 5´no codificante del genoma de Enterovirus detectado en tejido del sistema nervioso central de casos de enfermedades de las neuronas motoras, cambios en la estructura secundaria de la región que actúa como sitio de entrada ribosomal y sugirieron que la persistencia puede estar dada por una ineficiente traducción del polipéptido viral.
Gerling y otros,13 plantearon la posibilidad de que la diabetes mellitus puede ser causada por la infección persistente con Cox B4 defectivo no citopatogénico, en células B pancreáticas humanas. Otro Picornavirus (foot and mouth disease virus) puede causar infección persistente en cabras con viriones inmaduros con deleciones de hasta 3 kb.14
Los resultados obtenidos por estos autores sustentan los nuestros y permiten sugerir que las modificaciones en las proteínas estructurales encontradas en la cepa 44/93 IPK pueden repercutir en la persistencia de estos agentes de ECP-L, y perpetuar de esta manera el estímulo antigénico o producir daño por sí sola. Todos estos resultados nos permiten concluir que:
- Los agentes de ECP-L carecen de proteínas estructurales, por lo menos en su forma nativa.
- La persistencia puede ser considerada como un mecanismo por el cual estos virus participan en la etiopatogenia de la neuropatía epidémica en Cuba.
SUMMARY
During the epidemic of neuropathy occurred in Cuba from 1992 to 1994 viral agents antigenically related to the Coxsackie viruses were isolated from the cerebrospinal fluid of patients. To establish a function of these viruses in the etiopathogeny of the disease, strains 47/93 IPK identified as Coxsackie A9 and the strain 44/93 of light cytopathic effect were selected to conduct a study of their antigenic characteristics by Western Blot. It was proved that there is an antigenic relationship between both strains and it was also demonstrated the absence of structural proteins in their native form among the agents of light cytopathic effect. According to these results, it is stated the possibility that persistance may be a mechanism through which these viruses take part in the etiopathogeny of epidemic neuropathy in Cuba.Subject headings: NEURITIS/cerebrospinal fluid; NEURITIS/etiology; COXSACKIE VIRUSES/isolation & purification; CUBA.
REFERENCIAS BIBLIOGRÁFICAS
- Roman GC, Spencer PS, Schoenberg BS. Tropical myeloneuropathies: the hidden endemias. Neurology 1985;35:1158-70.
- Inoue YK. Inoue-Melnick virus and associated disease in man: recent advances. Prog Med Virol 1991;38:167-79.
- Más LP, Pelegrino JL, Guzmán MG, Comellas MM, Resik S, Álvarez M, et al. Viral isolation from cases of epidemic neuropathy in Cuba. Arch Path Lab Med 1997;121:825-33.
- Alfaro I, González-Quevedo A, Pino M del, Serrano C, Lara R, González H, et al. Immunoglobulins in epidemic neuropathy in Cuba. J Neurol Sci 1994;27:234-35.
- Borrajero I, Pérez JL, Domínguez C, Chong A, Coro RM, Rodríguez H, et al. Epidemic neuropathy in Cuba: morphologycal characterization of peripheral nerve lesions in sural nerve biopsies. J Neurol Sci 1994;127:68-76.
- Pérez AR, Pereira C, Hernández FO, Más LP, Balmaseda A, Menéndez R. Prevalence and factors associated to the epidemic neuropathy in population assisted family doctor in Cuba. Mem Inst Oswaldo Cruz 1996;91:543-50.
- Mertenz TH, Pika U, Eggers HJ. Cross antigenicity among enteroviruses as revealed by immunoblot technique. Virology 1983;129:431-42.
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970;227:680-85.
- Más LP, Guzmán MG, Muné M, Resik S, Álvarez M. Neuropatía epidémica cubana: Parte II: algunas características antigénicas de los aislamientos virales. Rev Cubana Med Trop 1995;47:16-20.
- Martino TA, Liu P, Petric M, Sole MJ. Enteroviral myocarditis and dilated cardiomyopathy: a review of clinical and experimental studies. En: Rotbart HA, ed. Human enterovirus infections. Washington: ASM Press, 1995;291-351.
- Vella C, Brown CL, Mc Carthy DA. Coxsackievirus B4 infection of the mouse pancreas: acute and persistent infection. J Gen Virol 1992;73:1387-94.
- Woodall CJ, Riding HM, Graaham DI, Clements GB. Sequences specific for enterovirus detected in spinal cord from patients with motor neurone disease. Br Med J 1994;308:1541-43.
- Gerling J, Chatterjee A. Lack of 64 000 Mr Islet autoantigen overexpression and antibody development following Coxsackievirus B4 infection in diabetes resistant mice. Autoimmunity 1993;14:197-203.
- Rewers M, Atkinson M. The possible role of enteroviruses in diabetes mellitus. En: Rotbart HA, ed. Human enterovirus infections. Washington: ASM Press, 1995;353-85.
Lic. Luis Sarmiento Pérez. Instituto de Medicina Tropical "Pedro Kourí". Apartado 601, Marianao 13, Ciudad de La Habana, Cuba.
1Master en Virología. Licenciado en Microbiología.
2 Doctor en Ciencias Médicas. Especialista de II Grado en Microbiología. Investigador De Mérito.
3 Especialista de I Grado en Microbiología.
4 Master en Virología. Licenciado en Microbiología. Investigador Auxiliar.
5 Técnico en Microbiología.

