










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión On-line ISSN 1561-3054
Rev Cubana Med Trop v.61 n.3 Ciudad de la Habana sep.-dic. 2009
ARTÍCULO ORIGINAL
Fenotipo antenal de dos especies cubanas del complejo flavida: Triatoma flavida y Triatoma bruneri (Reduviidae: Triatominae)
Antennal phenotype of two Cuban species of flavida: Triatoma flavida y Triatoma bruneri (Reduviidae: Triatominae) complex
Jinnay Rodríguez RodríguezI; Silvia CataláII; Omar Fuentes GonzálezIII; Jorge Fraga NodarseIV
I Licenciada en Biología. Máster en Entomología Médica y Control de Vectores. Departamento de Control de Vectores, Instituto de Medicina Tropical "Pedro Kourí" (IPK). Ciudad de La Habana, Cuba.
II Doctora en Ciencias Biológicas. Centro Regional de Investigaciones Científicas y Técnicas de La Rioja (CRILAR). La Rioja, Argentina.
III Licenciado en Biología. Departamento de Control de Vectores, IPK. Ciudad de La Habana, Cuba.
IV Licenciado en Bioquímica. Máster en Parasitología. Departamento de Parasitología, IPK. Ciudad de La Habana, Cuba.
RESUMEN
INTRODUCCIÓN: el fenotipo antenal ha mostrado ser una herramienta útil para la comparación taxonómica entre especies de Triatominae.
OBJETIVO: estudiar el fenotipo antenal en adultos de T. flavida y T. bruneri para clarificar el estado taxonómico de estas especies así como conocer la adaptación específica de cada una a su hábitat. Métodos: fueron contadas y analizadas 4 tipos de sensilla: bristles (BR), tricoide de pared fina (TPF), tricoide de pared gruesa (TPG) y basicónica (BA).
RESULTADOS: el pedicelo de T. flavida mostró solamente 2 tipos de receptores mientras que T. bruneri tuvo un mayor número de quimiorreceptores de 3 tipos diferentes. Entre las hembras de T. flavida y T. bruneri fueron observadas diferencias en los BR y BA, para los machos las diferencias fueron observadas en los receptores TPF, TPG y BA sobre el pedicelo y en los BR, TPG y BA en el flagelo 1. El análisis discriminante permitió la separación de las 2 especies, sin embargo, en el análisis de acuerdo con el sexo no se observaron diferencias.
CONCLUSIONES: como consecuencia de la adaptación de cada especie a su hábitat se observaron modificaciones morfológicas en el fenotipo antenal. Futuros estudios a los niveles ecológico, morfométrico y molecular son necesarios para clarificar el estado taxonómico de estas especies.
Palabras clave: Triatoma flavida, Triatoma bruneri, Triatominae, sensilla antenal, Cuba.
ABSTRACT
INTRODUCTION: the antennal phenotype has proven to be a useful tool for the taxonomic comparison between Triatominae species.
OBJECTIVE: to study the antennal phenotype in adults of T. flavida and T. bruneri adults to clarify the taxonomic status of these species as well as to learn about the specific adaptation of each of them to its habitat.
METHODS: four types of sensilla were counted and analyzed: Bristles (BR), thin walled trichoidea (TH), thick walled trichoidea (TK) and basiconic (BA).
RESULTS: the pedicel of T. flavida showed only two different types of receptors whereas T. bruneri males had a greater number of chemoreceptors of three different types. Among T. flavida and T. bruneri females, there were differences in BR and BA; differences in males were found in the receptors TH, TK and BA on the pedicel and BR, TK and BA in the flagellum 1. Discriminatory analysis allowed the separation of the two species; however, the analysis by sex did not show any difference.
CONCLUSIONS: as a consequence of the adaptation of each species to its habitat, morphological changes of the antennal phenotype occurred. So, further studies at ecological, morphometric and molecular levels are needed to make their present taxonomic status clear.
Keywords: Triatoma flavida; Triatoma bruneri; Triatominae; antennal sensilla; Cuba.
INTRODUCCIÓN
T. flavida es la especie más ampliamente distribuida en Cuba, es considerada selvática que habita en cuevas y, en ocasiones, ha sido encontrada en las casas atraída por la luz.1 Esta especie forma parte del complejo flavida junto con T. bruneri y T. obscura de Jamaica.2
T. flavida fue descrita por Neiva en 1911,3 más tarde Usinger en 19444 la redescribió como Nesotriatoma flavida y describió otra especie N. bruneri que luego en 1946 la consideró sinonimia de N. flavida.5 Lent y Wygodzinsky6 concuerdan con esta sinonimia, aunque no reconocen al género Nesotriatoma, sin embargo, plantean la necesidad de estudiar más material y la oportunidad de criarlos en el laboratorio, que permita validar la variabilidad poco usual del complejo citado.
En 1981, Lent y Jurberg7 redescriben las 2 especies T. flavida y T. bruneri por sus caracteres externos y por el estudio de la morfología de la genitalia externa de los machos. A pesar de eso, Marquetti8 señaló que en Cuba la taxonomía de estos insectos merece una revisión porque existen dudas de la veracidad de algunas de las especies reportadas.
La biología de T. flavida ha sido estudiada en condiciones de laboratorio,9 sin embargo, no existen estudios sobre la biología de T. bruneri. Además, la distribución geográfica de estas 2 especies no está bien delimitada y existen contradicciones en la literatura.
El número, tipo y distribución de sensilla sobre los segmentos antenales constituye el fenotipo antenal. Su estudio ha mostrado ser una herramienta útil para la comparación taxonómica entre género y especie de Triatominae y para ilustrar diferencias en la complejidad del nicho ecológico entre especies estrechamente relacionadas.10-12
El objetivo de este trabajo fue estudiar el fenotipo antenal en adultos de T. flavida y T. bruneri, para que unido a futuros estudios ecológicos, genéticos y morfométricos, permitan clarificar el estado taxonómico de estas especies, además de conocer las adaptaciones específicas de cada una relacionada con su hábitat.
MÉTODOS
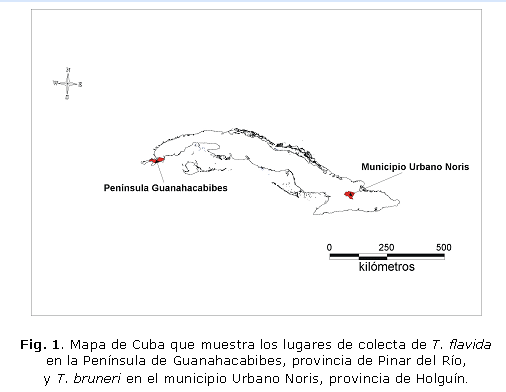
Insectos
Se analizó un total de 12 individuos de T. flavida, 6 colectados directamente en cuevas de la Península de Guanahacabibes (2 machos y 4 hembras) y 6 obtenidos en el laboratorio (4 machos y 2 hembras). Los ejemplares de T. bruneri (3 machos y 4 hembras) fueron colectados en casas del municipio Urbano Noris, Provincia de Holguin (Fig. 1). Estas especies fueron identificadas según la descripción de Usinger.4 Además fueron analizados 6 ejemplares selváticos de T. dimidiata, 3 machos colectados en la selva de Petén (Guatemala) y 3 hembras en Yucatán, México, los cuales se utilizaron como grupo externo en el análisis discriminante.
Preparación de la antena
Una antena por insecto fue cortada de la cabeza de cada individuo al nivel del escapo y almacenada en alcohol 70 %. Fueron aclaradas con KOH 4 %, neutralizadas con 5 % de ácido acético y montadas individualmente en glicerina para su observación al microscopio óptico a 400x. La superficie ventral del pedicelo y los 2 segmentos del flagelo fueron dibujados usando una cámara lúcida, identificando y contando de acuerdo con Catala y Schofield10 los receptores siguientes: mecanorreceptores (BR, Bristles), quimiorreceptores (TPF, tricoide de pared fina; TPG, tricoide de pared gruesa y BA, basiconica).
Análisis de los datos
Medias y desviación estándar del número de sensilla por tipo y segmento antenal de T. flavida y T. bruneri fueron calculados. Mediante la prueba de Levene se determinó la homogeneidad de las varianzas y después se aplicó un ANOVA para la comparación entre especies. En las variables que mostraron heterocedasticidad se usaron pruebas no paramétricas de U Mann-Whitney.
La antena de los especímenes de T. flavida, T. bruneri y T. dimidiata fueron analizadas usando un análisis discriminante multivariado (PADWIN) con 4 variables: los BR y el TPF del pedicelo y los TPG y BA del flagelo 1. PADWIN es un software desarrollado por JP Dujardin (http:///www.mpl.ird.fr/morphometrics) que permite hacer un análisis discriminante pero estima la significación estadística de forma no paramétrica, por una prueba de permutaciones. El número de permutaciones usadas fue de 1 000.
RESULTADOS
El número de sensilla, por tipo y por segmento, es mostrado en la tabla 1 según la especie y el sexo. No se observaron diferencias significativas entre los individuos selváticos y los del laboratorio, por lo que se consideró como una sola población.
Tabla 1. Número de sensilla antenal (media y desviación estándar) por segmentos de la antena de T. flavida, T. bruneri y T. dimidiata
| Especie | Sexo | n | Pedicelo | Flagelo I | Flagelo II | ||||||||||
| BR | TPF | TPG | BA | BR | TPF | TPG | BA | BR | TPF | TPG | BA | ||||
| Triatoma | Macho | 6 | 0 DE | 74,66 7,55 | 45,66 18,07 | _ | 1,00 1,09 | 14,50 2,58 | 62,66 10,42 | 113,83 9,86 | 15,50 4,59 | 8,20 1,09 | 36,80 7,59 | 122,20 11,73 | 32,00 8,80 |
| Hembra | 6 | 0 DE | 78,60 9,31 | 20,20 13,36 | _ | 0,80 0,83 | 15,20 1,30 | 63,40 11,12 | 125,60 13,64 | 10,40 4,87 | 7,40 1,94 | 30,60 6,10 | 120,60 17,38 | 22,20 4,96 | |
| Triatoma | Macho | 3 | 0 DE | 68,66 4,50 | 111,33 23,09 | 1,33 1,52 | 2,66 0,57 | 10,00 2,00 | 83,33 19,85 | 145,66 16,50 | 36,33 3,78 | 7,00 0,00 | 38,00 14,14 | 127,00 19,79 | 22,00 5,65 |
| Hembra | 4 | 0 DE | 71,60 7,09 | 32,60 5,77 | _ | 2,40 2,07 | 12,40 1,51 | 62,40 3,50 | 137,80 21,69 | 29,80 17,62 | 5,50 3,53 | 22,50 4,94 | 102,50 50,20 | 28,00 12,72 | |
| Triatoma dimidiata | Macho | 3 | 0 DE | 35,33 1,15 | 223,66 43,73 | 42,66 29,69 | 12 1,73 | 11,66 0,57 | 80,33 32,14 | 77,66 8,50 | 37 9,53 | ND | ND | ND | ND |
| Hembra | 3 | 0 DE | 37,66 3,05 | 173,33 17,78 | 64,33 22,14 | 12,33 4,16 | 13,66 1,15 | 72 2 | 66,33 19,55 | 22 6,08 | ND | ND | ND | ND | |
BR: Bristles, TPF: tricoide de pared fina, TPG: tricoide de pared gruesa, BA: basicónica, ND: datos no disponibles.
Caracteres generales de T. flavida y T. bruneri
T. bruneri presentó mayor cantidad de sensilla que T. flavida con un promedio de 1 160 y 1 022, respectivamente. El mayor número de sensilla se presentó en el flagelo 1 para ambas especies.
La antena de T. flavida y T. bruneri muestra todos los tipos de sensilla en el flagelo 1 y 2. En el pedicelo tanto hembras como machos tiene BR, TPF y BA.
En el caso de T. bruneri las hembras tienen BR, TPF y BA, mientras que los machos tienen BR, TPF, TPG y BA.
Dimorfismo sexual
Los machos de ambas especies presentaron antenas con mayor cantidad de sensilla que las hembras, con un promedio de 1 180 y 1 003, respectivamente.
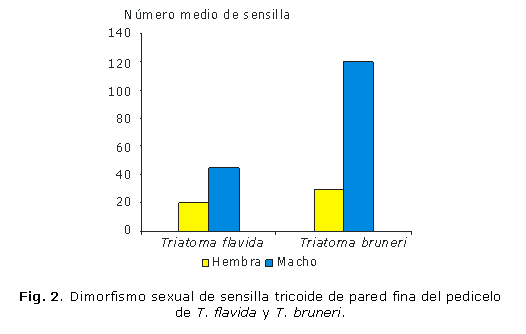
El número de TPF en el pedicelo es significativamente mayor en los machos que en las hembras (p< 0,05), tanto en T. flavida como en T. bruneri, aunque T. bruneri presentó un mayor dimorfismo sexual (p= 0,000004) que T. flavida (p= 0,015355) (Fig. 2). No se observaron diferencias en el número de los otros tipos de sensilla entre los sexos por especie.
Comparación sexual entre T. flavida y T. bruneri
Entre las hembras de T. flavida y T. bruneri se observaron diferencias en los BR y en la BA del flagelo 1. El número de BR de T. flavida es significativamente mayor que en T. bruneri (p< 0,05), mientras que el número de BA de T. bruneri es significativamente mayor que en T. flavida (p< 0,05). Se destaca que las hembras de ambas especies no poseen TPG en el pedicelo.
En relación con los machos, ambas especies difieren en los receptores siguientes: TPF, TPG y BA del pedicelo y en los BR, TPG y BA del flagelo 1. El número de TPF, TPG y BA del pedicelo, al igual que el número de TPG y BA del flagelo 1 en T. bruneri es significativamente mayor que en T. flavida (p< 0,05). Sin embargo, con respecto al número de BR del flagelo 1 de la especie T. flavida es significativamente superior que T. bruneri (p< 0,05).
Comparación entre T. flavida, T. bruneri y T. dimidiata
El análisis discriminante permitió separar significativamente las 3 especies (Fig. 3). Este estudio reclasificó 95 % de todos los individuos. La primera función discriminante representó 90 % de la variación total, mientras que la segunda función representó 10 % remanente. La distancia de Mahalanobis (Dm), calculada como la distancia entre grupos centroides entre las 3 especies fue significativa, con una reclasificación para T. flavida, T. bruneri y T. dimidiata de 100 %, 85 % y 100 %, respectivamente. La distancia más pequeña fue entre las especies T. flavida y T. bruneri (tabla 2).
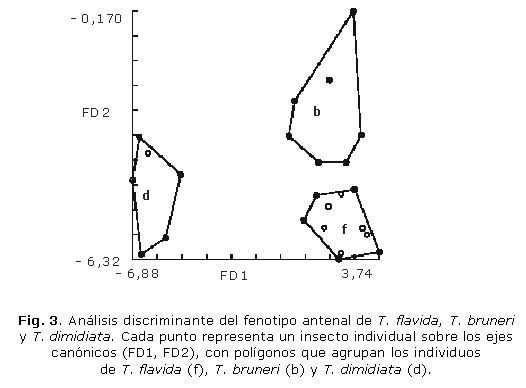

Un segundo análisis discriminante al analizar de manera separada hembras y machos de cada especie mostró que el fenotipo antenal de las hembras de T. flavida y T. bruneri no difieren significativamente (p> 0,05).
DISCUSIÓN
Especies de triatominos que viven en diferentes hábitats, muestran gran cantidad de quimiorreceptores de 3 tipos diferentes en el pedicelo, mientras aquellos adaptados a uno o pocos hábitats estables muestran ninguno o pocos quimiorreceptores en el pedicelo antenal.11 Este estudio muestra que T. flavida tiene pocos quimiorreceptores de solo 2 tipos (TPF y BA) en este segmento, lo cual pudiera estar relacionado con la explotación de un nicho espacial estable, como son las cuevas. Sin embargo, los machos de T. bruneri presentan mayor cantidad de quimiorreceptores de 3 tipos (TPF, TPG y BA), lo que sugiere mayor capacidad de dispersión que T. flavida y requiere mayor disponibilidad sensorial.
Un resultado notable obtenido en este estudio es la presencia del tipo de sensillum TPG solo en machos de T. bruneri y su ausencia en hembras de la misma especie y en T. flavida. La falta de este quimiorreceptor ocurre también en otras especies de Triatominae como T. platensis, T. rubrofasciata, T. infestans, T. protracta y T. brasiliensis.11 Resultados similares fueron reportados por otros autores,13 al encontrar sensillum de tipo TPG solo en machos alados de Mepraia sp., pero no fueron encontrados en hembras y machos de los insectos micropteros, lo cual sugiere la idea de que la ausencia de TPG en estos últimos se considera una pérdida secundaria ligada a la pérdida de la capacidad de vuelo.
En relación con las especies analizadas en este trabajo se pudiera inferir, que la ausencia de TPG en T. flavida estaría ligada a una disminución en la capacidad de dispersión, al considerar la cueva como su principal hábitat. En el caso de T. bruneri, su fenotipo antenal parece indicar que los machos de esta especie tendrían mayor capacidad de dispersión y colonización de nuevos hábitats, unido también a la mayor cantidad de sensilla del tipo TPF en el pedicelo. Estas sensilla fueron además observadas en estrecha relación con la capacidad de dispersión de Mepraia spinolai.13
Los sexos de ambas especies fueron distinguidos por un mayor número de TPF sobre el pedicelo de los machos, que coincide con resultados obtenidos en otros Triatominae y reafirma la hipótesis de que este tipo de sensillum está involucrada en la percepción de moléculas relacionada con la conducta sexual.14-16
La mayoría de las especies de Triatominae muestran mayor número de sensilla en el flagelo 1,12 lo cual fue demostrado en este estudio, al presentar los 2 sexos de T. flavida y T. bruneri mayor cantidad de sensilla en este segmento antenal en relación con los otros 2 segmentos estudiados.
El uso del análisis discriminante multivariado del fenotipo antenal ha mostrado ser útil para diferenciar géneros, especies y estadios de desarrollo.12 El presente estudio permitió diferenciar a través de este análisis las 2 especies cubanas del complejo flavida.
A pesar de que existen diferencias en el fenotipo antenal entre T. flavida y T. bruneri, el análisis multivariado según el sexo indicó que las hembras de estas 2 especies no difieren. Resultados similares fueron obtenidos al comparar hembras de Mepraia spinolai y Mepraia gajardoi, especies cercanas que forman parte del complejo spinolai.13
Los patrones de sensilla antenales en insectos hematófagos reflejan modificaciones de los patrones ancestrales, los cuales están modulados por las necesidades de cada especie en particular.17 Los resultados del presente estudio sugieren que las 2 especies cubanas del complejo flavida pudieran haber tenido un ancestro común y como consecuencia de la adaptación de cada especie a su hábitat, se produjeron modificaciones morfológicas del fenotipo antenal como las que han sido descritas en este trabajo. Sin embargo, futuros estudios a los niveles molecular, ecológico y morfométrico son necesarios para clarificar si estas especies deben o no mantener su presente estado taxonómico.
AGRADECIMIENTOS
Al doctor J. P. Dujardin por financiar el entrenamiento sobre fenotipos antenales. Al doctor C. J. Schofield quien amablemente revisó y mejoró el manuscrito. Este estudio fue financiado por la red ECLAT y el proyecto CDIA.
REFERENCIAS BIBLIOGRÁFICAS
1. Jiménez HO. Observaciones sobre la biología de Triatoma flavida Neiva, 1911 en Cuba. Rev Cub Med Trop. 1981;33:42-50.
2. Dujardin JP, Schofield CJ, Panzera F. Les Vecteurs de la Maladie de Chagas. Recherches Taxonomiques, Biologiques et genetiques. Brussels: Academie Royale des Sciences d Outre Mer; 2000.
3. Neiva A. Notas de entomología médica. Tres novas especies de redúvidas norte-americanas. Brasil-Médico. 1911;25:441.
4. Usinger RL. The Triatominae of North and Central America and the West Indies and their public health significance. Public Health Bull. 1944;288:1-83.
5. Usinger RL. Notas on Cuban Triatominae. Pan-Pacific Ent. 1946;22:19-20.
6. Lent H, Wygodzinsky P. Revision of the Triatominae (Hemiptera: Reduviidae), and their significance as vectors of Chagas disease. Bull Am Museum Nat History. 1979; 163:123-520.
7. Lent H, Jurberg J. As especies insulares de Cuba do género Triatoma. Rev Brasil Biol. 1981;431-9.
8. Marquetti MF. Triatomas. En: Llop AH, Valdes-Dapena MV, Suazo JL, editores. Microbiología y Parasitología Médicas. Tomo III. Ciudad de La Habana: Editorial Ciencias Médicas; 2001. p. 501-5.
9. Cabello DR, Lizano E. Biology of T. flavida Neiva, 1911 (Hemiptera: Reduviidae) under laboratory conditions. Mem Inst Oswaldo Cruz. 2001;96(6):879-81.
10. Catalá S, Schofield C. Antennal sensilla of Rhodnius. J Morphol. 1994;219:193-203.
11. Catalá S. Antennal sensilla of Triatominae (Hemíptera: Reduviidae): a comparative study of five genera. Int J Insect Morphol Embryol. 1997;26:67-73.
12. Gracco M, Catalá S. Inter-specific and developmental differences on the array of antennal chemoreceptors in four species of Triatominae. Mem Inst Oswaldo Cruz. 2000;95(1):67-74.
13. Moreno ML, Gorla D, Catalá S. Association between antennal phenotype, wing polymorphism and sex in the genus Mepraia. Infection, Genetics and Evolution. 2006;6(3):228-34.
14. Catala S, Carbajal A, Torres M, Moreno M, Ordoñez R, Montaña F, et al. La antena de los triatominae: caracteres ancestrales y marcadores funcionales. En: Guhl F, Schofield CJ, editores. Proceeding del Cuarto Taller Internacional sobre Genética Poblacional y Control de Triatomíneos, Punta Iguana, Cartagena, Colombia; 2000. p. 125-32.
15. Carbajal de la Fuente AL, Catalá S. Relationship among the habitat and the antennal sensilla pattern of six species of Triatominae (Hemíptera: Reduviidae). Mem Inst Oswaldo Cruz. 2002;97:1073-7.
16. Catalá S, Sachetto C, Moreno M, Rosales R, Salazar-Schettino PM, Gorla D. Antennal phenotype of Triatoma dimidiata populations and its relationship with species of phyllosoma and protracta complexes. J Med Entomol. 2005;42(5):719-25.
17. McIver, S. B., Siemicki, R. Fine structure of antennal putative thermo-hygrosensilla of adult Rhodnius prolixus Stal (Hemíptera: Reduviidae). J Morphol. 1985;183:15-23.
Recibido: 17 de octubre de 2008.
Aprobado: 5 de febrero de 2009.
Lic. Jinnay Rodríguez Rodríguez. Departamento de Control de Vectores. Instituto de Medicina Tropical "Pedro Kourí". Autopista Novia del Mediodía Km 6 ½. AP 601. Lisa, Ciudad de La Habana, Cuba. Correo electrónico: jinnay@ipk.sld.cu

