










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El cólera es una enfermedad infecciosa aguda de etiología bacteriana, identificada por Robert Koch en 1833, siendo la primera entidad para la cual se estableció un sistema de vigilancia e informe de brotes a gran escala.1
La Organización Mundial de la Salud (OMS) define el cólera como una infección intestinal aguda causada por la ingestión de agua o alimentos contaminados por la bacteria del género Vibrio la cual produce una enterotoxina que causa diarrea abundante, indolora y acuosa, que conlleva con rapidez a una deshidratación grave y la muerte, si no es tratada oportunamente, principalmente países en vías de desarrollo del continente africano, asiático y el Caribe.2
El agente causal Vibrio cholerae pertenece al género Vibrio, familia Vibrionaceae, siendo el patógeno más reconocido de esta familia por la diarrea que puede producir en los pacientes. Los principales factores de virulencia asociados a la enteropatogenicidad incluye la enterotoxina colérica y el factor de colonización lo que confiere la habilidad de colonizar en el intestino delgado. Además, existen otros factores de virulencia que han sido identificado en este género como son la producción de enzimas extracelulares gelatinasa, elastasa, lecitinasa, citolicinas, citotoxinas, hemolisinas y la producción de polisacárido capsular y hemaglutininas.3,4,5
La enfermedad del cólera requiere de rehidratación tanto por vía oral como endovenosa acompañada de una terapia antimicrobiana; se utiliza como tratamiento de elección tetraciclina (doxiciclina) y azitromicina, y como antimicrobiano alternativo la ciprofloxacina.6 No obstante en estudios internacionales se describen aislados de V. cholerae resistentes a múltiples antimicrobianos de uso frecuente, con un incremento de la resistencia a todos los antibióticos de uso frecuente, incluye ampicilina, quinolonas, ciprofloxacina, tetraciclina y trimetoprim/sulfametoxazol, lo cual pudiera estar debido a mecanismos de resistencia como la producción de betalactamasas de espectro extendido, las mutaciones cromosómicas, la presencia de bombas de eflujo y la adquisición de genes de resistencia por transferencia horizontal.5,6
La vigilancia de la enfermedad diarreica aguda (EDA) provocada por V. cholerae O1 representa un reto importante para el sistema nacional de salud en Cuba. Aunque desde la década de los 80 se han realizado estudios de caracterización fenotípica en esta especie, no se ha estudiado la asociación de la virulencia con la resistencia antimicrobiana en aislados de V. cholerae O1. Por tanto, el objetivo es determinar el comportamiento de la resistencia antimicrobiana frente a los antimicrobianos de interés empleados en el tratamiento, la presencia de factores de virulencia enzimáticos y si existe relación entre ambos.
Métodos
Se realizó un estudio descriptivo observacional de corte transversal durante el periodo de julio de 2012 a diciembre de 2015.
Se estudiaron 500 aislados de V. cholerae O1 identificadas, identificados con antelación, pertenecientes a la colección de cultivos del Laboratorio Nacional de Referencia de Enfermedades Diarreicas Agudas del Instituto de Medicina Tropical “Pedro Kourí” (LNR/EDA/IPK). Los aislados procedían de muestras de heces obtenidas de pacientes incluidos en los eventos de cólera notificados en Cuba, remitidos por la Red Nacional de Laboratorios de Microbiología, en el periodo descrito anteriormente.
Los aislados preservados en medio de conservación para enterobacterias (medio Pasteur), elaborado según las instrucciones del Manual de Medios y Reactivos del Instituto “Pasteur” (1978), se inocularon en agua peptonada alcalina (APA) y se incubaron en condiciones de aerobiosis a 37 ᵒC de 6-8 h. De cuatro a cinco asadas del cultivo en agua peptonada se sembraron por agotamiento en placas con agar MacConkey (BioCen/Cuba) y se incubaron en aerobiosis a 37 ᵒC de 18-24 h. Transcurrido este período, se seleccionaron tres colonias translúcidas, convexas y de bordes regulares y se inocularon por punción y estría en los medios de diferenciación primaria: agar hierro y dos azúcares de Kligler y agar hierro lisina (BioCen/Cuba), se incubó bajo las mismas condiciones descritas con anterioridad. La presencia de la enzima citocromo oxidasa se determinó mediante la prueba oxidasa.4
Se determinó la susceptibilidad de aislados de V. cholerae O1 frente a los agentes antimicrobianos de elección. Se utilizó el método de difusión en agar (Kirby - Bauer). Para la lectura e interpretación de los halos de inhibición se utilizaron los protocolos recomendados por Clinical and Laboratory Standard Institute (CLSI) (tabla 1).7,8
Se emplearon como cepas controles: Escherichia coli ATCC® 25922, Pseudomonas aeruginosa ATCC® 27853 y Staphylococcus aureus ATCC® 25923.
Tabla 1 Antimicrobianos que se emplearan en las pruebas de susceptibilidad
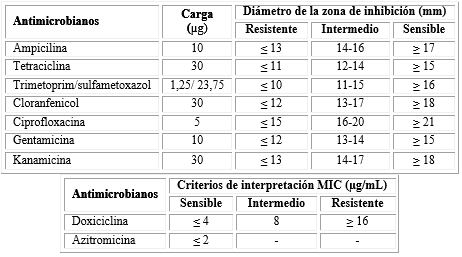
Fuente: CLSI. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. 27th ed. Pennsylvania: Clinical and Laboratory Standards Institute; 2017. CLSI. Clinical and Laboratory Standard Institute. Methods for antimicrobial dilution and disk susceptibility testing of infrequently isolated or fastidious bacteria. Approved Guideline-Second Edition. M45-A2. 2010;30(18):36-7
Criterio de multirresistencia: Se consideró como multirresistente aquellos aislamientos que mostraron resistencia a tres o más familias de antimicrobianos diferentes.9
Determinación de los factores de virulencia:
Los datos fueron almacenados y procesados con el programa Microsoft Office Excel 2015. El análisis de las variables cualitativas se realizó mediante la prueba de comparación de proporciones entre dos o más grupos, con el cálculo previo de las frecuencias absolutas y relativas. Cuando las frecuencias fueron pequeñas se usó la prueba de Fisher. La prueba de correlación de Spearman (r) se usó para determinar si existía una relación lineal entre las frecuencias de aislamientos resistentes, con las frecuencias de factores de virulencia, y si esta relación resultó estadísticamente significativa. En todos los casos los valores de probabilidad (p) se consideraron significativos cuando fueron menores de 0,05.
Todos los análisis fueron desarrollados empleando los paquetes de programas para análisis estadísticos GraphPadPrism versión 5.01 para Windows y Epidat 3.1.10
El protocolo de investigación que dio paso a los resultados de este trabajo fue aprobado por el Comité de Ética Institucional del Instituto de Medicina Tropical “Pedro Kourí” (CEI-IPK-85-17).
Resultados
La figura 1 muestra el comportamiento in vitro de los 500 aislados de V. cholerae O1 frente a los nueve agentes antimicrobianos. Al comparar las frecuencias de aislamientos sensibles, resistentes y con sensibilidad intermedia entre todos los antimicrobianos, se encontraron diferencias estadísticamente significativas (p< 0,05).
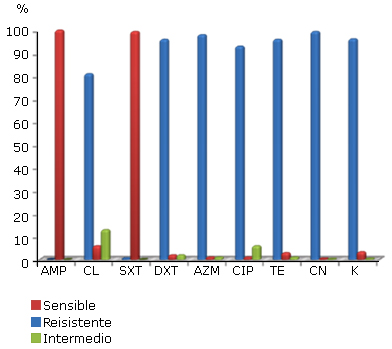 AMP: ampicilina; CL: cloranfenicol; SXT: trimetoprim-sulfametoxazol; DXT: doxiciclina; AZM: azitromicina; CIP: ciprofloxacina; TE: tetraciclina; CN: gentamicina; K: kanamicina.
AMP: ampicilina; CL: cloranfenicol; SXT: trimetoprim-sulfametoxazol; DXT: doxiciclina; AZM: azitromicina; CIP: ciprofloxacina; TE: tetraciclina; CN: gentamicina; K: kanamicina.Fig. 1 Susceptibilidad antimicrobiana en aislados de V. cholerae O1.
Respecto a la ampicilina y trimetoprim-sulfametoxazol, prevalecieron los aislados resistentes a ambos antimicrobianos (p< 0,005). Sin embargo, frente a los otros antimicrobianos investigados la frecuencia de aislamientos sensibles fue mayor (p< 0,005), con valores de susceptibilidad superiores al 95%. Aunque predominaron los aislamientos sensibles al cloranfenicol, este antimicrobiano mostró el mayor porcentaje de aislamientos con sensibilidad intermedia (p< 0,005), seguido por la ciprofloxacina.
En la tabla 2 se ilustra la distribución de los aislados según los patrones de multirresistencia. Del total de aislados estudiados, 44 (8,8 %) mostraron multirresistencia. Se evidenciaron por primera vez en el país, 17 patrones de multirresistencia, lo que constituye un hallazgo novedoso en comparación con estudios anteriores realizados en el LNR-EDA-IPK.
Tabla 2 Frecuencias de patrones multirresistencia de los aislados de V. cholerae O1
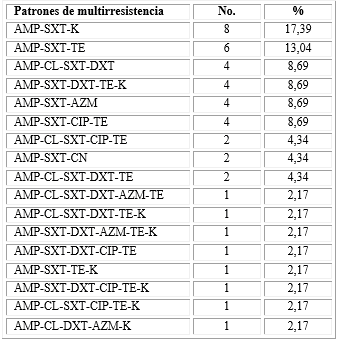
AMP: ampicilina; CL: cloranfenicol; SXT: trimetoprim-sulfametoxazol; DXT: doxiciclina; AZM: azitromicina; CIP: ciprofloxacina; TE: :tetraciclina; CN: gentamicina; K: kanamicina.
Factores de virulencia enzimáticos
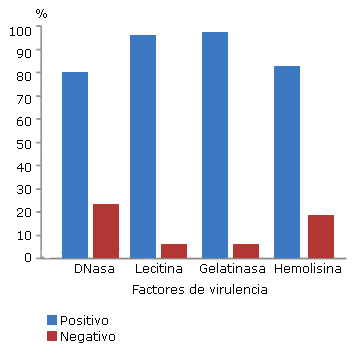
En la figura 2 se muestra la frecuencia de aislados que expresaron los factores de virulencia DNasa, lecitinasa, gelatinasa y hemolisina. Se observa que la frecuencia de aislamientos positivos a gelatinasa (96 %) y lecitinasa (95 %) fue mayor que aquellos que resultaron negativos a estos factores enzimáticos (p< 0,05).
Relación de la presencia de factores de virulencia enzimáticos con la resistencia a los antimicrobianos
Al observar los resultados se desprende que existe una correlación directamente proporcional entre la producción de las enzimas extracelulares y la resistencia antimicrobiana, o sea, a mayor frecuencia de aislamientos resistentes, mayor frecuencia de positividad de los factores de virulencia incrementándose en forma proporcional, lo cual se verifica para todos ellos con la prueba de correlación de Spearman (p< 0,05) (Fig. 3).
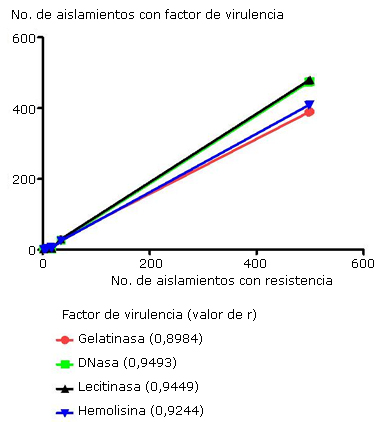
Discusión
Informes recientes indican que en los últimos años se ha constatado un incremento de la resistencia antimicrobiana a nivel mundial entre los aislamientos de V. cholerae frente a antibióticos de uso común, lo cual ha causado serios problemas en el tratamiento de los enfermos con cólera.11
Los resultados del presente estudio relacionados con los hallazgos de resistencia antimicrobiana ante trimetoprim-sulfametoxazol son semejantes a los que se han publicado en varias regiones de Asia, África y otras regiones de América, donde se advierte sobre la alta incidencia de resistencia a este antimicrobiano.11 Por ejemplo, en Ghana, Eibach y otros, en 92 aislamientos procedentes de un brote de cólera en niños mayores de 5 años muestran un 100 % de resistencia ante el trimetoprim-sulfametoxazol; sin embargo, los porcentajes de resistencia para la ampicilina obtenidos en esta investigación difieren de lo publicado por Eibach, pues solo obtienen susceptibilidad intermedia en dicho antimicrobiano.11
Otras investigaciones realizadas en Haití indican que en el análisis de 122 aislados de V. cholerae O1, procedentes de los 10 departamentos de ese país entre los años 2010-2011, obtuvieron elevados porcentajes de resistencia al trimetoprim-sulfametoxazol y valores de susceptibilidad intermedios para ampicilina, ciprofloxacina y cloranfenicol en dos de los aislados.11,12 Por otra parte, en el año 2016 se informó en Nepal que 100 % de los aislamientos de V. cholerae analizados por Gupta y otros, resultaron resistentes a la ampicilina y al trimetoprim-sulfametoxazol, mientras que 96,77 % resultó sensible frente al cloranfenicol.12
Sobre la base de las evidencias anteriores Thapa y otros., 2015, sugieren que los antimicrobianos como la ampicilina y el trimetoprim-sulfametoxazol no deben ser empleados en el tratamiento de la enfermedad del cólera, debido a la evidencia de la existencia en la especie de V. cholerae de diferentes mecanismos de resistencia como la transferencia horizontal de genes de resistencia o las mutaciones espontáneas.9
En el caso de la resistencia antimicrobiana obtenida en los aislamientos de V. cholerae frente a la ampicilina en este estudio podría deberse a la producción de betalactamasa de espectro extendido (BLEE), ya que la presencia de esta enzima se demuestra en varios estudios según la bibliografía, como los de Gosh y otros, en 2011, donde 92,8 % de los aislados existe la presencia de la misma,12 lo que podría deberse a la utilización por la bacteria de otros mecanismos de resistencia a la ampicilina que incluye alteraciones en la permeabilidad de la membrana a este antibiótico o a la presencia de bombas de eflujo que lo exportan. Investigaciones sobre los sistemas de eflujo en V. cholerae han determinado que la presencia de bombas de eflujo RDN en esta especie esté involucrada con la resistencia antimicrobiana.13,14
Así mismo, los porcentajes evidenciados frente al trimetoprim-sulfametoxazol se pudieran atribuir a la posibilidad que tiene V. cholerae de portar el elemento SXT, que pertenece a alguna clase de EIC (elemento integrador conjugativo), descrito por vez primera por Waldor et al., como un elemento que le confiere la habilidad de adquirir genes de resistencia que provienen de enterobacterias resistentes a sulfametoxazol, trimetropim y la estreptomicina. En la actualidad muchos aislados de los serogrupos O1 y O139 alrededor del mundo ha adquirido el elemento SXT a través de diseminación natural.15
En este estudio se evidenciaron aislados con sensibilidad intermedia frente al cloranfenicol y la ciprofloxacina que incluye los aislados con concentraciones mínimas inhibitorias (CIMs) cercanas a los niveles de antibiótico usualmente alcanzados en sangre o en los tejidos y para los cuales el grado de respuesta podría ser menor que con las cepas “sensibles”. Esto pudiera relacionarse con la presencia de mecanismos como resistencia mediada por plásmidos y el elemento integrador conjugativo para el primero y las mutaciones en los genes gyrA y parC, los integrones y las bombas de eflujo para el segundo.16
En Cuba se han realizado varios estudios de susceptibilidad antimicrobiana en V. cholerae. En el 2004 los aislamientos mostraron bajos niveles de resistencia frente a ampicilina (14 %) y trimetoprim -sulfametoxazol (16 %);17 mientras que en 2008 se constata un aumento de los valores de resistencia hasta un 30,7 % con respecto a la ampicilina, manteniéndose niveles similares de susceptibilidad frente al trimetoprim-sulfametoxazol y disminuyendo los referentes a la tetraciclina y al cloranfenicol en un 13 % y 11 %, respectivamente.4
No obstante, en el 2016 se publicó un nuevo estudio realizado a 144 aislados de V. cholerae O1, los cuales mostraron valores de resistencia a ampicilina y trimetoprim-sulfametoxazol por encima de 90 %, lo que representa un incremento con respecto a los valores obtenidos en los estudios previos, encontrándose en concordancia con los hallazgos de esta investigación.
En cuanto a doxiciclina, se encontró 94,1 % de sensibilidad, valor que es elevado, pero que pudiera sugerir que circulan aislamientos que han adquirido los plásmidos que portar resistencia para las tetraciclinas, por lo que se puede afirmar la necesidad de vigilancia de la resistencia de V. cholerae ante este medicamento, ya que es la primera opción terapéutica en los casos de cólera, y en la realización de quimioprofilaxis a los contactos. De igual forma los valores obtenidos frente a la azitromicina podría estar relacionado con la mayor actividad demostrada por este fármaco frente a V. cholerae,18 ya que su uso se ha recomendado solo para los pacientes de edad pediátrica o mujeres embarazadas con sospecha de EDA por cólera y como segunda opción en el resto de los casos.
Estudios publicados en Cuba por Fernández y otros, en el 2017, en V. cholerae no O1/no O139 mostraron un 4,8 % con patrones de multirresistencia (AMP-,C-,TE-; AMP-,SXT-,C; AMP-,SXT-,SUL-,AK-; AMP-,CIP-,SXT-,SUL-; AMP-, SXT-,C-,TE-,SUL-,DXT) diferentes a los de la presente investigación.19Ceccarelli y otros, en el 2016, en un estudio realizado en V. cholerae, concluyeron que independientemente del serotipo, este microorganismo posee una elevada plasticidad genómica y una historia de exitosa asociación con plásmidos, tramsposones y elemento integrador conjugativo(ICE) que favorecen el fenotipo multirresistente que hoy caracteriza a este patógeno.20
El número de aislados multirresistentes encontrado en el presente estudio podría estar relacionado con la presencia de integrones en los aislados, ya que los mismos son una de las causas principales de multirresistencia entre los enteropatógenos, tal y como reportan Bakhshi y otros, quienes sugieren que la presencia de un integrón clase 1 está distribuido entre los enteropatógenos, por lo que se hace necesario hacer estudios moleculares.19,21 Por esta razón, monitorear los cambios en los patrones de susceptibilidad e identificar los mecanismos de resistencia, constituye una prioridad para el control de la diseminación de los determinantes genéticos de resistencia ya que la resistencia ha sido reconocida en la actualidad por la OMS como un problema extremadamente serio.19,22
Estudios realizados a nivel internacional en el género Vibrio sobre mecanismos implicados en la patogenicidad y virulencia, como son la producción de citotoxinas, proteasas y enzimas extracelulares; ha demostrado que los mismos están implicados en la aparición de hemorragias, edemas y alteraciones del sistema de defensa en el organismo. Muchos de estos cambios se ha debido a la presión de selección provocada por la introducción de los antimicrobianos, favoreciendo el desarrollo de los procesos infecciosos.19,23,24
En este trabajo se encontró que los factores de virulencia enzimáticos la gelatinasa y la lecitinasa estuvieron presentes en la mayor cantidad de aislados, seguidos de hemolisinas y DNasa. En Cuba, estudios realizados por Cabrera y otros, en el 2008, a 65 cepas procedentes de muestras de heces, se demostró la presencia de al menos un factor de virulencia en todos los aislados. Se evidenció que la hemolisina y gelatinasa (100 %) se encontraron en mayor proporción, seguido de DNasa (78,8 %) y lecitinasa (80 %). Los resultados de dicha investigación son opuestos a los del presente estudio donde se obtuvo en mayor porcentaje o presencia a gelatinasa y lecitinasa.23
Este resultado es similar a los encontrados en Cuba por Pérez y otros, en el 2013, en un estudio realizado a 70 aislamientos procedentes de muestras clínicas; no siendo así para el resto de la presencia de los factores de virulencia .(25) Asimismo, en el 2017 Fernández y otros, en estudios relacionados con la resistencia y la presencia de genes de virulencia en 125 aislamientos de V. cholerae no-O1/no-O139 identificados en muestras de origen clínico, demostraron la presencia de la enzima gelatinasa (100 %), lecitinasa (77,6 %), hemolisina (61,6 %) y para la DNasa (60 %).21
Los resultados de la presente investigación sobre la presencia de factores de virulencia enzimáticos en los aislamientos puede estar relacionado con el hecho de que los microorganismos pueden producir varias proteínas extracelulares e intracelulares para invadir el sistema inmunológico del hospedero y lograr su colonización, ya que V. cholerae tiene varias “herramientas” en su arsenal para lograr completar el proceso infeccioso.24
Los mecanismos bacterianos implicados en la patogenicidad y virulencia son en la actualidad objeto de numerosos estudios en el ámbito de la microbiología médica infecciosa. Sin embargo, estos mecanismos han experimentado un proceso largo evolutivo dependiente de la relación hospedero-patógeno. Muchos de estos cambios se han debido a la presión de selección provocada por la introducción de los antimicrobianos en medicina.25
Conclusiones
El estudio de la susceptibilidad antimicrobiana demuestra la factibilidad de continuar con el esquema de tratamiento antimicrobiano de primera y segunda línea establecidos por el Programa Nacional de Enfermedades de Trasmisión Digestiva para pacientes con cólera en Cuba.
Se evidencia por primera vez en el país aislados de V. cholerae con un espectro de multirresistencia incrementado, lo que constituye una expresión del avance de la resistencia antimicrobiana en la especie.
Se demuestra que todos los aislados cubanos estudiados poseen al menos dos de los factores de virulencia enzimáticos reconocidos en la especie, sugiriendo este hecho un alto potencial de virulencia.
Se evidencia una relación directa y proporcional entre la presencia de los factores de virulencia y resistencia antimicrobiana, sinergismo que surgiere mayor patogenicidad de los aislados estudiados procedentes de brotes epidémicos.

