










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Investigaciones Biomédicas
versión On-line ISSN 1561-3011
Rev Cubana Invest Bioméd v.21 n.2 Ciudad de la Habana abr.-jun. 2002
Temas de actualización
Instituto de Medicina Tropical “Pedro Kourí”
Estrés oxidativo en la infección por el virus linfotrópico de células T humano de tipo I (vlth-I)
Lic. Lizette Gil, Dr. Alejandro Álvarez, Dr. Héctor Díaz y Dr. Jorge Pérez
Resumen
Se reportó lo más actualizado en relación con el virus linfotrópico de células T humano de tipo I (VLTH-I), que es un retrovirus asociado entre otras enfermedades a la leucemia de células T del adulto y la paraparesia espástica tropical. La patología es el resultado de la actividad de la proteína viral Tax que participa en el incremento de la producción de especies reactivas del oxígeno requeridas para la activación de NF-kB, y que transactiva la expresión de genes virales y celulares que resulta en la transformación celular, la inmortalización y la enfermedad. La caracterización de estos mecanismos contribuye a una mejor comprensión de la fisiopatología de la infección y en la perspectiva de intervención terapéutica preventiva.
DeCS: VLTH-I/inmunología; VIRUS LINFOTROPICO DE CELULAS T HUMANO//fisiopatología; ESTRES OXIDATIVO.
El virus linfotrópico de células T en humanos tipo I (VLTH-I) se reconoce como el agente etiológico de la leucemia-linfoma de células T del adulto (LTA) y la paraparesia espástica tropical o mielopatía asociada al VLTH-I (PET/MAV). LTA fue reconocida como una nueva enfermedad en 1977, reportada por Junji Yodoi en 1974 como el primer cáncer humano causado por un virus. A juzgar por observaciones recientes, el espectro de enfermedades posiblemente asociadas con este virus parece ser más amplio y pudiera incluir otras afecciones que hasta el presente se han considerado de causas autoinmunes o desconocidas. La infección por VLTH-I es endémica en el suroeste de Japón, centro y sur de África, América Central, suroeste de EE.UU. y en el área del Caribe; aunque se reportan casos en otras áreas geográficas. Se estiman entre diez y veinte millones de personas portadoras en el mundo.1,2
Teniendo en cuenta que la infección por VLTH-I es endémica en el área del Caribe, en Cuba se han realizado varios estudios de vigilancia seroepidemiológica. Desde 1978 hasta 1991 se estudiaron 3 031 personas y 2 resultaron seropositivas, lo que se correspondió con un índice de seropositividad de 0,065 %. Esto motivó a estandarizar un ELISA para la detección de anticuerpos VLTH-I/II y de un western blot para la confirmación. Hasta el 31 de diciembre de 1997 se habían estudiado 29 423 personas y 13 de ellas se diagnosticaron como seropositivas al VLTH-I, para un índice general de seropositividad de 0,044 %, que representa un bajo índice con respecto al área.3,4
El VLTH-I es un retrovirus de reconocido carácter inmunoproliferativo, pero los mecanismos por los cuales la infección desarrolla cualquiera de estas patologías no está del todo esclarecido y se han mantenido resistentes a la mayoría de las terapias convencionales. Numerosas evidencias experimentales señalan que el estado de oxidación/reducción celular es esencial en el desarrollo de los organismos vivos, en la fisiología normal, la activación e inactivación de biomoléculas, la transducción de señales, la obtención de energía metabólica, y el recambio y la activación celular, así como en la defensa antimicrobiana y antitumoral.
Algunas proteínas (tioredoxina [TRX], superóxido dismutasa, glutatión peroxidasa, catalasa y bcl-2) relacionadas con el estado redox intervienen en los procesos de muerte y proliferación celular. El estrés oxidativo, desequilibrio redox, caracterizado por un exceso en la producción de especies reactivas del oxígeno (ERO), aparejado o no a un debilitamiento de los sistemas antioxidantes, ha sido asociado a la inducción de apoptosis en numerosas infecciones virales, entre ellas la infección por VIH-I y la infección por VLTH-I.5-8
En la infección por VLTH-I se ha demostrado la desregulación del receptor de IL-2 (IL-2Ra/CD25), lo que llevó a la identificación de la TRX como un inductor del CD25 producido por las células LTA y al reconocimiento del papel de la regulación redox en la fisiopatología de la infección.9 Este artículo es una revisión de los progresos recientes en el campo del estrés oxidativo y la fisiopatología de la infección por VLTH-I.
VLTH-I, IL-2Ra y TRX
El HTLV-I, descubierto por Poiesz y otros en EE.UU. en 1980, en pacientes diagnosticados con LTA, se ha relacionado con un aumento en la expresión del receptor de IL-2 (IL-2Ra/CD25). El ARNm de IL-2Ra/CD25 está aumentado constitutivamente en las células de pacientes LTA y puede ser liberado de manera soluble. Esta evidencia sugiere el posible papel del IL-2 en la fisiopatología de la infección, pero algunas células infectadas no requieren de IL-2 para su crecimiento in vitro y la mayoría de las líneas celulares LTA no proliferan en respuesta a IL-2. Por esto se considera la sobreexpresión de IL-2Ra como un indicador del estado de actividad de las células infectadas, pero el mecanismo fisiopatológico queda por esclarecer.10,11
La TRX, reconocida inicialmente como factor derivado de células LTA (ADF), fue definida como un factor inductor de IL-2Ra producido por las células infectadas. Esta proteína está presente en muchas especies desde plantas hasta mamíferos, es pequeña (12 kDa) y tiene un grupo redox activo ditiol/disulfuro en el sitio activo conservado: Cis-Gli-Pro-Cis. Su expresión es inducida por varios tipos de estrés: infección viral, mitógenos, rayos X, rayos UV, peróxido de hidrógeno y reperfusión posisquémica. El gen que codifica para la TRX humana fue reportado recientemente en el cromosoma 9. El promotor del gen contiene elementos que responden a la acción de citosinas y elementos que responden a estrés oxidativo. Numerosos resultados de investigaciones realizadas en diferentes enfermedades apuntan a la asociación de la disminución de la TRX con disfunciones del sistema inmune.11,12
El estado redox intracelular es esencial para la viabilidad celular, la activación y la proliferación. La TRX junto con NADPH y TRX reductasa opera como un sistema proteico de reducción de disulfuros, de vital importancia en el mantenimiento del estado redox celular. Su función es tanto intracelular como extracelular. Aunque el glutatión es el antioxidante intracelular de mayor cuantía, la TRX desempeña también un papel de antioxidante endógeno.
La TRX participa de la regulación de las interacciones proteína-nucleótidos. Una de las funciones más importante de TRX es la activación de factores de transcripción, entre ellos NF-kB13 y AP-114 (fig.). La unión de NF-kB al sitio kB es regulada por factores redox. Después de la disociación de I-kB, es necesaria la reducción de NF-kB para su unión al sitio activo. El residuo cisteína 62 es el sitio blanco reducido por TRX para su interacción con el ADN.12
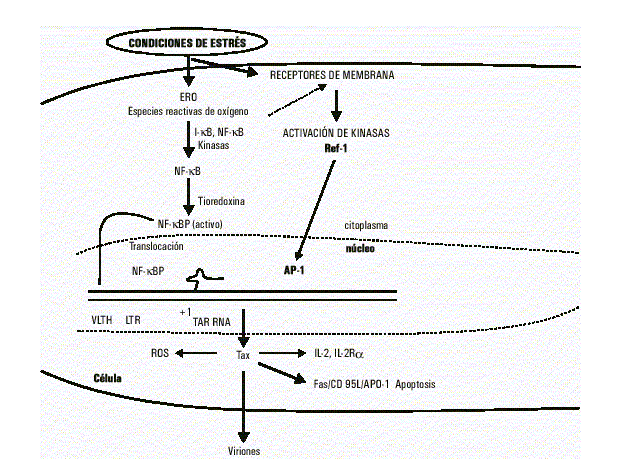
Fig. Diversas biomoléculas de los sistemas oxidante y antioxidante están involucradas en la regulación de la transcripción del VLTH-I mediada por la activación de Tax, NF-kB, AP-1. La generación del estrés oxidativo producto de estímulos externos contribuye al aumento de la replicación viral, al daño de las biomoléculas estructurales y a la activación de las señales celulares de apoptosis (CD 95L), todo esto influye en el desarrollo de estados patológicos relacionados con la progresión de la infección al estado LTA, HEM.
La unión de AP-1 al elemento de respuesta TRE está influenciado por condiciones de reducción como la presencia de TRX. En las células, el factor nuclear redox Ref-1 está involucrado en la regulación del residuo cisteína de AP-1. Nuevos reportes aseveran la interacción directa de TRX y Ref-1; estos estudios reportan la existencia de una cascada redox involucrada entre la activación de TRX y la liberación de AP-114 (fig.). La unión de otros factores de transcripción, incluidos c-Myb y p53, a sus elementos cis, está modulada por el estado redox.
En los límites de la membrana celular, la TRX también desempeña un papel citoprotector frente al estrés oxidativo y la transducción de señales. La TRX promueve el consumo de cisteína en las células y regula los niveles intracelulares de glutatión. Como los residuos cisteínas son importantes para la síntesis del glutatión, el efecto citoprotector es en parte dependiente de los niveles intracelulares de glutatión. La TRX junto con la glutaredoxina constituyen donadores de electrones a la glutatión peroxidasa, que ejerce una importante función antioxidante en el plasma.
La TRX es secretada o liberada por los linfocitos y otros tipos de células como hepatocitos y fibroblastos y su aumento es indicativo de respuesta inflamatoria frente al estado de estrés oxidativo.12, 13
Estrés oxidativo e infección por VLTH-I
El VLTH-I a partir del gen Tax codifica para una proteína de 40 kD localizada primariamente en el núcleo de las células infectadas, en la región conocida como matriz nuclear. La proteína Tax es un transactivador de la transcripción del promotor en el LTR 5’ del genoma del provirus. Ahora se conoce que la proteína Tax es muy promiscua y capaz de transactivar promotores heterólogos, además de su propio LTR. El mecanismo de acción de esta proteína es indirecto sobre otros promotores y existen reportes de la inducción in vitro de la transformación maligna a través de la represión de genes básicos reguladores celulares, mediada por proteínas bHLH en presencia de Tax15-17 (fig.).
Evidencias clínicas de la existencia de estrés oxidativo en pacientes asintomáticos y enfermos de LTA, y de PET aseveran su persistencia desde el estado inicial de la infección, y relacionan esta con el proceso de apoptosis de linfocitos T, hospederos preferenciales del virus.18,19
Investigaciones recientes demuestran la activación hormonal de Tax. Se ha demostrado que Tax estimula un estado prooxidante in vitro en células Jurkat infectadas. Este fue medido como cambios en los niveles intracelulares de glutatión y peróxido de hidrógeno libre, los cuales fueron determinados y relacionados con la muerte celular por apoptosis. El estrés oxidativo generado en células Jurkat T transfectadas establemente con HTLV-I está involucrado en la activación de NF-kB, transactivador celular nuclear que es blanco de las especies reactivas del oxígeno (ERO) y es fundamental en la expresión de genes como: el receptor de IL-2 (CD-25) y el marcador de apoptosis (Fas/CD95/APO-1). De lo que se puede sugerir que una de las funciones de Tax es estimular la producción de ERO que son necesarias para la activación de NF-kB20-23 ( fig.).
Este hecho ha sido corroborado con la adición micromolar del antioxidante ditiocarbamato pirrolidina a células Jurkat T donde se suprimió la activación de NF-kB y fue afectada también la transactivación del virus por Tax.24
La activación sostenida de la replicación viral por cambios hormonales resulta en una selección de clones infectados que pierden la actividad Tax, lo que ocasiona la proliferación celular y la inhibición del proceso de apoptosis. Esto pudiera corresponder con la fisiopatología de la enfermedad de LTA.24
Experimentos in vitro reportan que una elevada concentración de 7,8-dihidroneopterina amplifica la actividad de Tax, lo que puede influir en una estimulación de la apoptosis y en la alteración del estado redox.25
Consideraciones finales
Esta serie de experimentos apuntan actualmente al análisis de la relación entre el estado redox y las funciones y el desarrollo celular. Estas evidencias sugieren que el estado redox junto con la estimulación antigénica y de citosinas contribuyen a una vía de señalización intracelular y en algunos casos las ERO constituyen moléculas señalizadoras. El estado de equilibrio entre oxidantes y antioxidantes en el ciclo de vida de células infectadas con VLTH-I, puede modular el daño a las diferentes biomoléculas estructurales y el proceso de apoptosis que ocurre, esto señala que el estado redox está muy relacionado con la patogénesis de enfermedades como la LTA y PET. Lo anterior constituye un potencial terapéutico en los desórdenes asociados a infecciones por VLTH-I.
Summary
The most updated information is given about the human T cell lymphotropic virus type 1 (HTLV-1), which is a retrovirus associated with T-cell leucemia of the adult and with tropical spastic paraparesis, among other diseases. The pathology is the result of the activity of Tax viral protein that takes part in the increase of the production of reactive oxygen species required for the activation of NF-kB, and that transactivates the expression of viral and cellular genes resulting in the cellular transformation, immortalization and disease. The characaterization of these mechanisms contributes to a better understanding of the physiopathology of the infection and to the perspective of preventive therapeutic intervention.
Subject headings: HTLV-I/immunology; HUMAN LYMPHOTROPIC VIRUS TYPE I/physiopathology; OXIDATIVE STRESS.
Referencias bibliográficas
- Yodoi J, Takatsuki K, Masuda T. Letter: two cases of T-cell chronic lymphocytic leukemia in Japan. N Engl J Med 1974;290:572-3.
- Uchiyama T, Yodoi J, Sagawa K, Takatsuki K, Uchino H. Adult T cell leukemia: clinical and hematologic features of 16 cases. Blood 1977;50:481-92.
- Rolo F, Armas MB de, Mato J, Lubian AL, Diaz H. Confirmation of the presence in Cuba of human T-cell lymphotropic virus type I using the polymerase chain reaction. Rev Cubana Med Trop 1997;47(3):204-8.
- Lubian AL, Diaz HM, Silva E, Perez MT, Cruz O, Fuente JL de la, et al. Seroprevalence of the HTLV-I infection in different risk groups studied in Cuba. Rev Cubana Med 1998;37(4):199-204.
- Eisenlohr LC, Yewdell JW, Bennink JR. Pathogenesis of chronic progressive myelopathy associated with human T cell lymphotropic virus type I. J Exp Med 1992;175:481-7.
- Nakamura H, Nakamura K, Yodoi J. Redox regulation of cellular activation. Annu Rev Immunol 1997;15:351-69.
- Schwarz KB. Oxidative stress during viral infection- a review. Free Radic Biol Med 1996;21:64411-9.
- Buttke TM, Sandstrom PA. Oxidative stress as a mediator of apoptosis. Immunol Today 1994;15:7-10.
- Crenon I, Beraud C, Simard P, Montagne J, Veschambre P, Jalinot P. The transcriptionally active factor mediating the effect of the HTLV-I tax transactivator on the IL-2 R alpha kappa B enhancer included the product of the c-rel protooncogene. Oncogen 1993;8:867-75.
- Yodoi J, Uchiyama T. Human T cell leukemia virus type-I (HTLV-I) associated diseases; virus, Il-2 receptor dysregulation, and redox regulation. Immunol Today 1992;13:405-10.
- Hayashi T, Ueno Y, Okamoto T. Oxidoreductive regulation of nuclear factor kappa-B. Involvement of a cellular reducing catalyst thioredoxin. J Biol Chem 268:11380-8.
- Okamoto T, Sakurada S, Yang J, Merin J. Regulation of NF-kB and diseases control: Identification of a novel serine kinase and thioredoxin as effectors for signal transduction pathway for NF-kB activation. Current Top Cell Reg 1997;35:149-61.
- Hirota K, Matsui M, Iwara S, Nishiyama A, Mori K, Yodoi J. AP-1 transcriptional activity is regulated by a direct association between thioredoxin and Ref-1. Proc Natl Acad Sci 1996 (en prensa).
- Okamoto T, Ogiwara H, Hayashi T, Mitsui A, Kawabe T, Yodoi J. Human thioredoxin/adult T cell leukemia derived factor activates the enhancer binding protein of human immunodeficiency virus type 1 by thiol redox control mechanism. Int Immunol 1992;4:811-9.
- Mattews MA, Markowits RB, Dynan WS. In vitro activation of transcripcion by the Human T-cell leukemia virus type I Tax protein. Mol Cell Biol 1992;12(5):1986-96.
- Yoshida M. HTLV-I Tax: regulation of gene expression and disease. Trends Microbiol 1993;1(4):131-5.
- Uittenbogaard MN, Armstrong AP, Chiaramello A, Nyborg JK. Human T-cell leukemia virus type I Tax protein represses gene expression through the basic helix-loop-helix family of transcripcion factors. J Biol Chem 1994;269(36):22466-9.
- Furuke K, Sasada T, Euda-Taniguchi Y, Yamauchi A, Inamoto T, Yamaoka Y, et al. Role of intracellular redox status in apoptosis induction of Human T cell leukemia virus type I infected lymphocytes by 13-cis retinoic acid. Cancer Res 1997;57:4916-23.
- Los M, Khazaie K, Schulze-Osthoff K, Baeuerle PA, Schirrmarcher V, Chlichlia K. Human T-cell leukemia virus-I (HTLV-I) Tax-mediated apoptosis in activated T cells requires an enhanced intracellular prooxidant state. J Immunol 1998;161(6):3050-5.
- Kohno T, Yamada Y, Hata T, Mori H, Yamamura M, Tomonoga M, et al. Relation of oxidative stress and gluthatione synthesis to CD95 (Fas/APO-1) mediated apoptosis of adult T cell leukemia cells. J Immunol 1996;56:4722-8.
- Abate C, Patel L, Rauscher FJ III, Curan T. Redox regulation of Fos and Jun DNA binding activity in vitro. Science 1990;249: 1157-61.
- Curtis VE, Smilde R, McGuire KL. Requirements for interleukin 2 promoter transactivation by the Tax protein of human T-cell leukemia virus type I. Mol Cell Biol 1996;16(7):3567-75.
- Li Xh, Gaynor RB. Regulation of NF- kB by the HTLV-I Tax protein. Gene Exp 1999;7(4-6):223-45.
- Schreck R, Grassmann R, Fleckenstein B, Baeuerle PA. Antioxidants selectively suppress activation of NF-kappaB by human T-cell leukemia virus type I Tax protein. J Virol 1992;66(11):6288-93.
- Baier-Bitterlich G, Baier G, Fuchs D, Bock G, Hausen A, Utermann G, et al. Role of 7,8-dihydroneopterin in T-cell apoptosis and HTLV-I transcription in vitro. Oncogene 1996;13(10):2281-5.
Recibido: 11 de octubre de 2001. Aprobado: 11 de enero de 2002.
Lic. Lizette Gil. Instituto de Medicina Tropical “Pedro Kourí”. Apartado 601, Marianao 13, Ciudad de La Habana, Cuba. Correo electrónico: lgil@ipk.sld.cu

