










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Investigaciones Biomédicas
versión On-line ISSN 1561-3011
Rev Cubana Invest Bioméd v.26 n.4 Ciudad de la Habana oct.-dic. 2007
Determinación de anticuerpos IgG contra el ADN de doble cadena mediante ELISA utilizando como recubrimiento ADN xenógeno y alógeno
Determination of IgG antibodies versus dsDNA by ELISA using xenogenous and halogenous DNA coating
Dra. Gipsis Suárez Román; Dr. Antonio Mario González Griego; Dra. Tammy Fernández Romero; Dra. Victoria Esther González Ramírez
RESUMEN
Se evaluó la utilidad del ADN alógeno como antígeno de recubrimiento para detectar los anticuerpos IgG contra el ADN de doble cadena. El ADN xenógeno empleado como recubrimiento de un enzimoinmunoensayo indirecto, para la determinación de estos anticuerpos, es de difícil obtención y muy costoso. Se empleó como recubrimiento ADN de timo de ternera y ADN de 6 sujetos supuestamente sanos. El análisis de la presencia de los anticuerpos se realizó en suero de 16 individuos con diagnóstico de lupus eritematoso sistémico. Se obtuvo 100 % de coincidencia en los resultados, así como pocas diferencias en la capacidad de discriminación del método con el empleo de los 2 tipos de ADN. Según lo anterior se concluyó que el ADN humano es útil como recubrimiento de este enzimoinmunoensayo. La obtención de este antígeno es menos costosa que la del ADN de timo de ternera.
Palabras clave: Enzimoinmunoensayo, anticuerpos IgG contra el ADN de doble cadena, ADN xenógeno, ADN alógeno.
SUMMARY
The usefulness of allogenous DNA as a coating antigen to detect IgG antibodies versus dsDNA was evaluated. The xenogenous DNA used as a coating of an indirect immunoenzimatic assay for determining these antibodies is difficult to obtain and very expensive. Calf thymus DNA and DNA from 6 apparently sound subjects was utilized for coating. The analysis of the presence of antibodies was made in serum from 16 individuals with diagnosis of systemic lupus erythematosus. 100 % of coincidence was obtained in the results. A few differences in the discrimination capacity of the method with the use of two DNA types were observed. According to the above, it was concluded that human DNA is useful as a coating of this enzymoimmunoassay. The obtention of such antigen cost less than that of the calf thymus.
Key words: Enzymoimmunoassay, IgG antibodies versus dsDNA, xenogenous DNA, halogenous DNA.
INTRODUCCIÓN
Los anticuerpos contra el ADN de doble cadena (anti-ADNdc) fueron descubiertos en 1957. Su presencia constituye uno de los criterios diagnósticos de la Asociación Americana de Reumatología para el lupus eritematoso sistémico (LES).1-3
Numerosas pruebas para la determinación de anti-ADNdc han sido ideadas. Entre ellas se encuentran el radioinmunoanálisis (RIA), la inmunofluorescencia indirecta con Crithidia luciliae (CLIF), el ELISA (enzyme linked immunosorbent assay), el EliA (fluorescence enzyme linked immunoassay) y la citometría de flujo.1-3
La detección de anti-ADNdc mediante ELISA está ampliamente difundida en los laboratorios clínicos por las ventajas que esta técnica posee: la relativa sencillez, la rapidez de ejecución, el bajo costo del equipamiento, la posibilidad de automatización y el procesamiento de gran número de muestras a la vez. Además emplea pequeñas cantidades de muestras y reactivos y presenta elevada precisión, sensibilidad, especificidad y detectabilidad.4-6
Existen juegos diagnósticos comerciales disponibles que utilizan como recubrimiento ADN plasmídico, ADN bacteriano, ADN de timo de ternera y ADN humano recombinante, los cuales son muy costosos. A esto se adiciona que estos antígenos son difíciles de obtener.1,7-9
Para que el diagnóstico del LES se haga en condiciones que se acerquen más al concepto de autoinmunidad y lograr una mayor especificidad, así como disponer de un antígeno obtenido en nuestros propios laboratorios por un método fácil, sencillo y menos costoso, este trabajo tiene el objetivo de evaluar la utilidad del ADN genómico humano (alógeno) como recubrimiento del enzimoinmunoensayo para la determinación de IgG anti-ADNdc.
MÉTODOS
Muestras
Se recogió suero de 16 pacientes con diagnóstico de LES, según criterios de la Asociación Americana de Reumatología modificados en 1982 y revisados en 1997. Se obtuvo sangre total fresca de 6 individuos supuestamente sanos destinados a la obtención del antígeno. Se empleó suero de personas consideradas sanas como control negativo en los ensayos, procedente del banco de sangre ubicado en el municipio Marianao. Se utilizó suero de individuos enfermos con más de 100 DE (desviaciones estándar) o unidades de positividad como control positivo.
Obtención del antígeno
Extracción de ADN genómico humano: 10 mL de sangre fresca se mezclaron con 250 µL de EDTA (56 mg/mL) como anticoagulante en tubo corning de 50 mL. Se añadió solución reguladora fosfato salino (PBS 1x) hasta completar 50 mL, se homogenizó suavemente y se centrifugó a 4 000 rpm durante 15 min a 4 °C, luego se desechó el sobrenadante. Se resuspendió el pellet con solución de lisis celular (TLB) y se colocó durante 30 min a 4 °C. Se centrifugó a 4 000 rpm durante 15 min y se desechó el sobrenadante. Se le añadió al pellet 3 mL de solución de lisis nuclear (NLB), 230 µL de SDS 10 % y 60 µL de proteinasa K y se incubó 1 h a 55 °C y luego a 37 °C durante toda la noche. Se le añadió 1 mL de NaCl 6 M y 5 mL de cloroformo-alcohol isoamilico, se precipitó el ADN con 2-propanol y se lavó con etanol 70 %. El pellet se disolvió en Tris-EDTA.
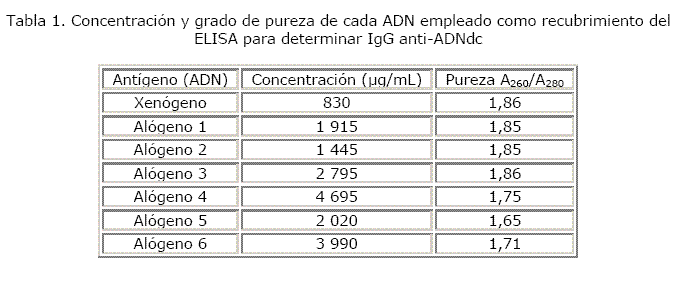
La concentración del ADN extraído fue medida espectrofotométricamente a 260 nm y la pureza fue confirmada mediante la razón A260/A280.
Determinación de IgG anti-ADNdc mediante ELISA indirecto
Se utilizó como antígeno ADN genómico de 6 personas supuestamente sanas (alógeno) y ADN de timo de ternera (xenógeno), obtenido de manera comercial (Merck). Las condiciones empleadas con ambos tipos de antígenos fueron las siguientes: las placas de microtitulación de polivinil cloruro (PVC) fueron irradiadas con luz ultravioleta durante 16 h. Luego se cubrieron con ADN xenógeno y 3 ADN alógenos disueltos en PBS 1x, pH 7,2 a concentración de 10 µg/mL. Después de incubación en cámara húmeda a 4 °C durante toda la noche, las placas fueron lavadas con PBS 1x. Las muestras y los controles diluidos en PBS-Tween + BSA 2 % fueron añadidos a una dilución 1:100 e incubados en cámara húmeda a 37 °C durante 1 h. Se realizó lavado con PBS-Tween 0,05 % y se añadió IgG anti-humana conjugada con peroxidasa de rábano picante diluido 1:4 000 en PBS-Tween + BSA 2 %. Se incubó en cámara húmeda a 37 °C durante 1 h, se efectuó lavado con PBS-Tween 0,05 % y se añadió sustrato (o-fenilendiamina, Merck). Se incubó en la oscuridad a temperatura ambiente durante 30 min, deteniéndose la reacción con H2SO4 3M. Los valores de absorbancia fueron medidos a 492 nm mediante lector de micro ELISA (Organontechnika).
En cada placa se corrió 1 control negativo, 1 control positivo y 4 muestras. Cada placa se consideró un ensayo. Se efectuaron un total de 4 ensayos (1 al 4). En el primer ensayo se utilizaron 3 ADN alógenos diferentes a los empleados en el segundo, tercero y cuarto.
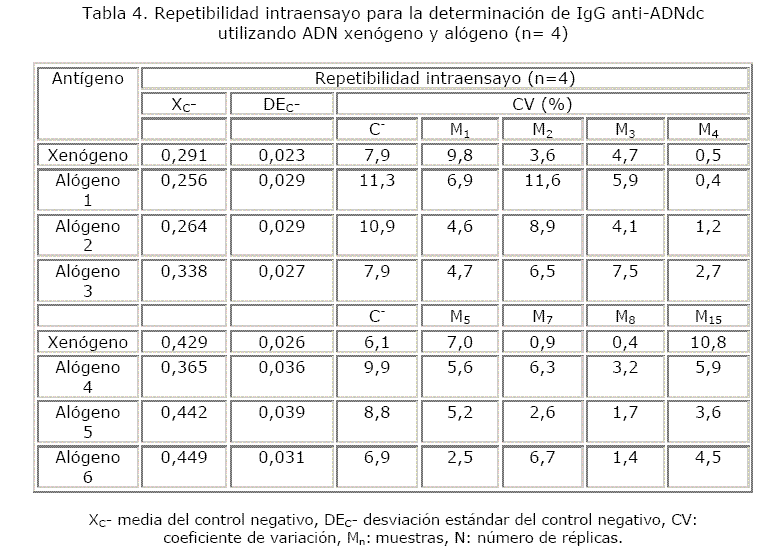
Estudios de precisión: repetibilidad intraensayo e interensayos
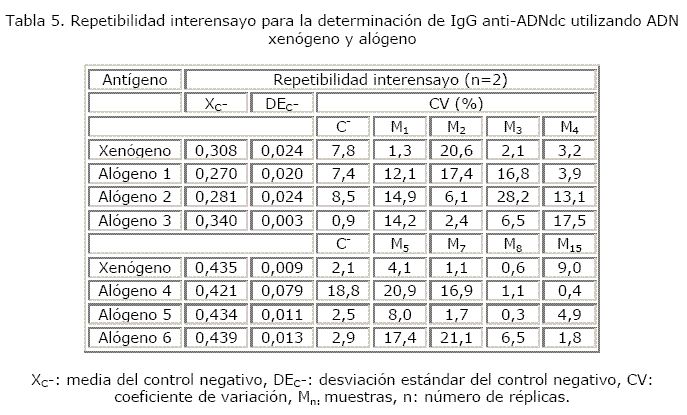
Se montaron 4 réplicas de las muestras y de los controles positivo y negativo para evaluar repetibilidad intraensayo. Cada ensayo se repitió 2 veces, en días diferentes, pero manteniendo las mismas condiciones para evaluar su reproducibilidad.
Análisis de los resultados
Los coeficientes de variación (CV) se determinaron según los métodos estándares. Se calculó la capacidad de discriminación (K) para cada antígeno. Esta es una variable cuantitativa continua, la cual se refiere a la capacidad de diferenciar entre los valores positivos y los negativos. A mayor valor de K mejor es el poder discriminatorio. Es adimensional.
K= XC+ / XC-
XC+ - Densidad óptica media del control positivo.
XC- - Densidad óptica media del control negativo.
La cuantificación de anticuerpos IgG anti-ADNdc (CA) se determinó mediante la aplicación de la fórmula siguiente:
CA=Xm - Xc- / DEc-
Xm - Densidad óptica media de la muestra.
Xc- - Densidad óptica media del control negativo.
DEc- - Desviación estándar del control negativo.
La CA es una variable cuantitativa continua que se expresa en desviaciones estándar (unidades de positividad). Informa la cantidad de veces que está contenida la desviación estándar del control negativo en la muestra. La desviación estándar usualmente es empleada para establecer un valor de corte que permite emitir un criterio cualitativo, o sea, la presencia o no de una sustancia o metabolito en un determinado fluido biológico. Nuestro laboratorio emplea la desviación estándar para dar un criterio cuantitativo, lo cual desde el punto de vista médico tiene gran importancia. Esto posibilita la individualización de la terapéutica de los pacientes lúpicos.
Los criterios con los que se trabajó para 99,9 % de confianza fueron los siguientes: se tomó como valor negativo de referencia cuando CA ≤ 3 DE (unidades de positividad). Se consideró ideal si K ≥ 10, aceptable si 5 ≤ K < 10 y desfavorable si K < 5. La obtención de un CVK ≤ 10 % indicaría la no existencia de diferencias notables en los resultados, un 10 % < CVK ≤ 20 % significaría la existencia de diferencias poco notables en los resultados y si CVK > 20 % implicaría la existencia de diferencias notables. Se tomó un CV intraensayo e interensayo como ideal si CV ≤ 10%, aceptable si 10 % < CV ≤ 20 % y desfavorable si CV > 20 %.
RESULTADOS
El grado de pureza de los 2 tipos de ADN empleados en el estudio se encontró dentro del rango aceptado como ideal (1,6-2,0). Esto indica que el ADN no estaba contaminado con proteínas (tabla 1).
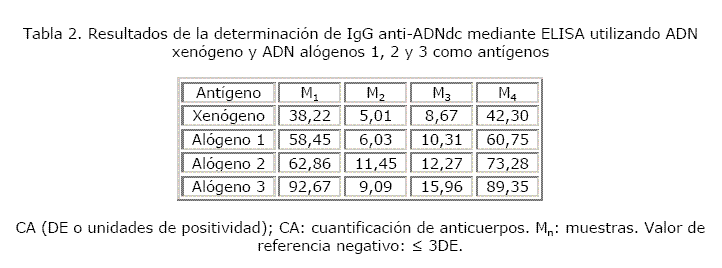
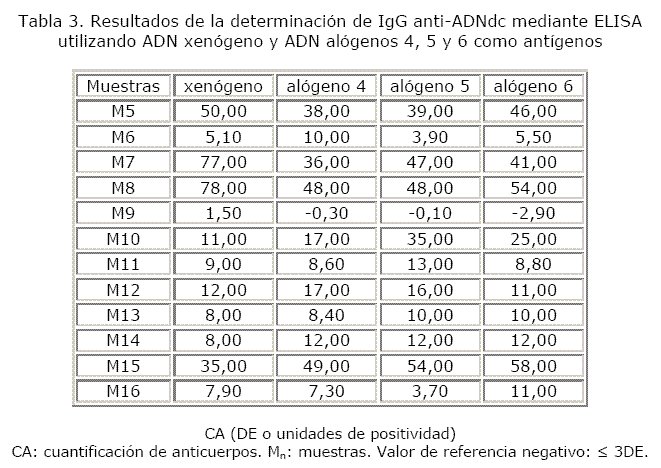
Un total de 16 muestras séricas de individuos con diagnóstico de LES fueron analizadas. Hubo 100 % de concordancia en cuanto a los resultados obtenidos con los diferentes antígenos empleados. En las tablas 2 y 3 se puede observar que 15 de las 16 muestras fueron positivas y solo una negativa con el empleo del ADN xenógeno y de los diferentes ADN alógenos.
En el primer ensayo las capacidades de discriminación (K) del método con el uso de ADN xenógeno y con los ADN alógenos 1, 2 y 3 resultaron aceptables (8,1; 9,3; 8,8 y 7,0, respectivamente). Las K obtenidas en los ensayos 2 y 3 fueron las siguientes: 5,7; 4,9; 5,2 y 4,5 — 6,4; 5,0; 5,4 y 4,4, respectivamente. Con el empleo del ADN alógeno 6 en los ensayos 2 y 3 el método mostró una capacidad de discriminación desfavorable, al igual que con el alógeno 4 en el ensayo 2. En el ensayo 4 se obtuvieron valores de K aceptables tanto con el ADN xenógeno como con los ADN alógenos 4, 5 y 6 (6,9; 5,5; 5,5 y 5,3) (fig).
En los ensayos 2, 3 y 4 la K con el ADN xenógeno fue superior a la obtenida con los ADN alógenos, ocurriendo lo contrario con 2 de los alógenos en el primer ensayo realizado (fig.).
El coeficiente de variación entre las capacidades de discriminación (CVK) con el empleo de cada antígeno se mantuvo por debajo de 10 % en el ensayo 2. En los ensayos 1, 3 y 4 el CVK tomó valores entre 10 y 20 % (fig.).
La mayoría de los coeficientes de variación de los controles negativos y de las muestras estuvieron por debajo de 10 % y una minoría está comprendida en el rango aceptable. Cuando se evaluó la reproducibilidad, algunas muestras (M2 con el ADN xenógeno, M3 con el ADN alógeno 2, M4 con el ADN alógeno 4 y M7 con el ADN alógeno 6) alcanzaron coeficientes de variación desfavorables (tablas 4 y 5).
DISCUSIÓN
La línea que se ha trazado en Cuba, como país en vías de desarrollo, es la de producir, en la medida de las posibilidades, los productos biológicos que se requieren en las técnicas analíticas. Este principio es el que llevó a evaluar la utilidad del empleo del ADN genómico humano (alógeno) como recubrimiento del ELISA para detectar anticuerpos anti-ADNdc en el diagnóstico del LES.
En este estudio solo se determinaron anticuerpos anti-ADNdc de tipo IgG porque son más específicos para el LES que los de tipo IgM.3,8
Se obtuvo 100 % de coincidencia en los resultados con el uso de los 2 tipos de ADN, lo cual indica que cualquiera de ellos puede emplearse como recubrimiento en el método ELISA. En la literatura se reporta que los anticuerpos anti-ADNdc reaccionan contra epítopes conformacionales del eje pentosa-fosfato que es común a todos los ADN.3 Hay autores que plantean que los anti-ADNdc reaccionan específicamente contra secuencias teloméricas del ADN, las cuales son características de cada especie. A pesar de esto las secuencias teloméricas en mamíferos superiores no muestran muchas diferencias.10
La otra variable que se tuvo en cuenta en este estudio fue la capacidad de discriminación del método. En su mayoría, las K obtenidas con cada tipo de antígeno en los 6 ensayos llevados a cabo fueron aceptables. En esto influye si se utilizan sustancias antes del recubrimiento, si los reactivos están en buen estado, la pureza del antígeno y su grado de polimerización, si el antígeno es modificado, la calidad de las soluciones empleadas, etc.1,6,11-13
En el ELISA llevado a cabo no se emplea ninguna sustancia pre-recubrimiento como sulfato de protamina, polihistidina o polilisina, que pueden conllevar a la aparición de falsos positivos.1,11,12 Además, el ADN se utiliza en su forma intacta, o sea, no es fragmentado. Cuando el antígeno es fragmentado se pueden producir reacciones no específicas con esos fragmentos que conducen a resultados falsos positivos.12,13
Las soluciones reguladoras o amortiguadoras usadas fueron preparadas y almacenadas cuidadosamente para evitar su contaminación bacteriana, porque las que contienen fosfato y Tris constituyen un medio excelente para saprófitos, bacterias, etc. No se adicionó a las soluciones amortiguadoras ningún agente antimicrobiano como azida o mercurio pues estos tienen un efecto devastador sobre la actividad enzimática en sistemas, que como este, usan peroxidasa.1,12
Tanto el ADN xenógeno como el ADN genómico alógeno poseían grados de pureza adecuados. A mayor grado de pureza mayor cantidad de ADN se adhiere al pocillo, el riesgo de interferencias disminuye y se produce una mejor señal. Ello contribuye a lograr una buena capacidad de discriminación del método. Precisamente, la utilización de un antígeno muy purificado constituye uno de los requisitos a tener en cuenta a la hora de realizar un enzimoinmunoensayo.1,4,6
El ADN de timo de ternera presentó un grado de pureza (1,86) mayor al de los ADN alógenos 4, 5, 6 (1,75; 1,65 y 1,71, respectivamente). Basado en lo expuesto en el párrafo anterior se explica que en los ensayos 2, 3 y 4 la K del método con el empleo del ADN xenógeno fuera mayor a la K con cada ADN alógeno utilizado. Aunque el ADN xenógeno y los ADN alógenos 1, 2 y 3 utilizados en el ensayo 1 poseían grados de pureza similares, se observa como la K obtenida usando el ADN alógeno 1 y 2 fue superior a la K mostrada con el ADN xenógeno (tabla 1) (fig.).
El grado de polimerización del antígeno influye notablemente sobre la K. A mayor grado de polimerización se incrementa la posibilidad de que no se pierdan los epítopes conformacionales contra los que se supone reaccionan los anti-ADNdc.1,4,6 Se conoce que el ADN de timo de ternera posee un elevado grado de polimerización (según Directorio Linscott de reactivos inmunológicos y biológicos), el cual no fue determinado en los ADN alógenos usados.
A pesar de estas diferencias los coeficientes de variación entre la capacidad de discriminación del método con los antígenos empleados en cada ensayo demostraron que estas no son importantes. Ello se explica por lo expuesto antes al hacer referencia al grado de coincidencia en los resultados.
El costo estimado de una extracción de ADN genómico humano por el método empleado es aproximadamente 3 USD. Este cálculo está basado en el costo de los reactivos, el tiempo de funcionamiento de los equipos, el tiempo empleado por los técnicos para la realización de la técnica, etc. No se incluyen la cristalería, ni el costo de los equipos. Teniendo en cuenta que el volumen mínimo que se puede obtener en una extracción de ADN genómico humano es de 150 µL, se estima que el costo de 1 mL sería de 20 USD.
De manera general se logró una buena precisión de los resultados. Algunas muestras presentaron coeficientes de variación desfavorables, atribuible a su posible contaminación.
Se concluyó que el ADN alógeno es un antígeno tan útil como el ADN xenógeno para la detección de anticuerpos IgG anti-ADNdc mediante ELISA. El ADN alógeno puede ser obtenido en los laboratorios cubanos por un método sencillo y poco costoso.
REFERENCIAS BIBLIOGRÁFICAS
1. Rose NR, Hamilton RG, Detrick B. Manual of Clinical Laboratory Immunology. 6th ed. Washington: Editorial ASM Press; 2002.
2. Oldchowka N. Methods for detection of anti-dsDNA autoantibodies. Elias J 2002;1:7-8.
3. Hahn BH. Antibodies to DNA. N Engl J Med 1998;338(19):1359-68.
4. Ochoa F. Bases metodológicas para la evaluación de anticuerpos en ensayos clínicos de vacunas mediante técnicas inmunoenzimáticas. La Habana: Ediciones Finlay; 2004.
5. Roitt I, Brostoff J, Male D. Inmunología. 5a ed. Madrid: Ed. Harcourt; 2001.
6. Reina M. ELISA. [en línea] 2003 [fecha de acceso enero de 2005]. Disponible en: http://www.ub.es/biocel/wbc/tecnicas/elisa.htm
7. Gonzalez C, Guevara P, Garcia-Berrocal B, Navajo J, Gonzalez-Buitrago J. Clinical evaluation of cobas core anti-dsDNA EIA quant. J Clin Lab Anal 2004;18(3):200-5.
8. Janyapoon K, Jivakanont P, Surbrsing R, Siriprapapan W, Tachawuttiwat T, Korbsrisate S. Detection of anti-dsDNA by ELISA using different sources of antigen. Pathology 2005; 37(1):63-8.
9. Wigand R, Gottschalk R, Falkenbach A, Matthias T, Kaltwasser JP, Hoelzer D. Detection of dsDNA antibodies in diagnosis of systemic lupus erythematosus-comparative studies of diagnostic effectiveness of 3 ELISA methods with different antigens and a Crithidia luciliae immunofluorescence test. Z Rheumatol 1997;56(2):53-62.
10. Wallace DJ, Salonen EM, Avaniss-Aghajani E, Morris R, Metzger AL, Pashinian N. Anti-telomere antibodies in systemic lupus erythematosus: a new ELISA test for anti-DNA with potential pathogenetic implications. Lupus 2000;9(5):328-32.
11. Kroubouzos G, Tosca A, Konstadoulakis M, Varelzidis A. Poly-L-lysine causes false results in ELISA methods detecting anti-dsDNA antibodies. J Immunol Methods 1992;148:261-3.
12. Janyapoon K, Siriprapapun W, Techawuttiwat T, Jivaganon P. Evaluation of optimal conditions for dsDNA coating on microtiter plates for anti-dsDNA detection by ELISA. Bull Health Sci Tech 2003;6:31-8.
13. Pitsetsky D, González T. The influence of DNA size on the binding of antibodies to DNA in the sera of normal human subjets and patient with systemic lupus erythematosus (SLE). Clin Exp Immunol 1999;116:334-9.
Recibido: 14 de junio de 2007.
Aprobado: 30 de junio de 2007.
Dra. Gipsis Suárez Román. Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón" Calle 146 No. 3102 esquina Ave. 31. Cubanacán, municipio Playa. Ciudad de La Habana. Teléf.: 208- 48- 77. Correo electrónico: gipsis@giron.sld.cu
Departamento de Inmunología. Centro Nacional de Genética Médica.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}