










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.30 n.4 supl.20074 Matanzas oct.-nov. 2007
ARTÍCULO DE INVESTIGACIÓN
Estudio de la descomposición de la hojarasca en un pastizal de Panicum maximum Jacq cv. Likoni
Study of litter decomposition in a pastureland of Panicum maximum Jacq cv. Likon
Saray SánchezI, G. CrespoII y Marta Hernández I
I Estación Experimental de Pastos y Forrajes ¨Indio Hatuey¨. Central España Republicana, CP 44280, Matanzas, Cuba
E-mail: saray.sanchez@indio.atenas.inf.cu
II Instituto de Ciencia Animal, La Habana, Cuba
RESUMEN
Se realizó un estudio en la Estación Experimental de Pastos y Forrajes ¨Indio Hatuey¨, Matanzas, con el objetivo de determinar la descomposición de la hojarasca de Panicum maximum y su relación con algunos de los factores bióticos y abióticos. La descomposición de la hojarasca se determinó como la pérdida de biomasa a través del tiempo, con relación al peso inicial. Para el estudio de la dinámica de la descomposición se utilizó el método de bolsas de hojarasca (litter bags). Las bolsas medían 10 x 10 x 10 cm, con poros de 1 cm de diámetro, lo cual permite el acceso al interior de un amplio rango de la biota edáfica. Se distribuyeron al azar 40 bolsas que contenían hojarasca de P. maximum. En cada bolsa se colocaron 20 g de hojarasca (base seca) previamente colectada. Los resultados demostraron que el proceso de descomposición de la hojarasca de P. maximum fue lento, ya que a los 210 días la hojarasca aún presentaba el 45,3% de su peso inicial, con una mayor tasa de descomposición en los primeros 30 días. En el material remanente del proceso de descomposición de la hojarasca, la concentración del N-FND se incrementó. La composición taxonómica de la fauna asociada al proceso de descomposición estuvo constituida por tres phylum, cinco clases, ocho órdenes, siete géneros y siete especies determinadas. La presencia de una diversa y estable fauna asociada a las bolsas influyó en el proceso de descomposición de la hojarasca. Las especies Venezillo sp., Nagarus sp. y Pycnocelus surinamensis fueron las de mayor presencia durante todo el proceso. El modelo matemático que contempla el efecto de la temperatura, la humedad relativa y la precipitación, mostró el mejor ajuste en el proceso de descomposición de la hojarasca en este pastizal.
Palabras clave: Factores climáticos, fauna del suelo, hojarasca, P. maximum
ABSTRACT
A study was carried out at the Experimental Station of Pastures and Forages “Indio Hatuey”, Matanzas, with the objective of determining the litter decomposition of Panicum maximum and its relationship to some of the biotic and abiotic factors. The litter decomposition was determined as the loss of biomass in time, with regards to the initial weight. For the study of the decomposition dynamics the method of litter bags was used. The bags measured 10 x 10 x 10 cm, with pores of 1 cm diameter, which allows access inside of a wide range of the edaphic biota. Forty bags that contained litter of P. maximum were distributed randomly. In each bag 20 g of previously collected litter (dry base) were put. The results showed that the decomposition process of P. maximum litter was slow, because 210 days after being deposited the litter still showed 45,3% of its initial weight, with a higher decomposition rate in the first 30 days. In the remnant material of the process of litter decomposition, the N-NDF concentration increased. The taxonomic composition of the fauna associated to the decomposition process was constituted by three phyla, five classes, eight orders, seven genera and seven determined species. The presence of a diverse and stable fauna associated to the bags influenced the process of litter decomposition. The species Venezillo sp., Nagarus sp. and Pycnocelus surinamensis were the ones with higher presence
during the whole process. The mathematical model that shows the effect of temperature, relative humidity and rainfall, showed the best adjustment in the process of litter decomposition in this pastureland.
Key words: Climatic factors, litter, Panicum maximum, soil fauna
INTRODUCCIÓN
La descomposición de la hojarasca constituye la vía de entrada principal de los nutrientes en el suelo y es uno de los puntos clave del reciclado de la materia orgánica y los nutrientes (Vitousek, Turner, Parton y Souford, 1994; Aerts y De Caluwe, 1997). Varios autores han estudiado con detalle la dinámica de la descomposición de la hojarasca de plantas leñosas, tanto en climas templados como en el mediterráneo (González y Gallardo, 1995; Álvarez-Sánchez y Becerra, 1996; Berg, 2000). Sin embargo, hay pocos estudios sobre la dinámica de la descomposición de la hojarasca en pastizales (Bardgett y Shine, 1999), a pesar de su importancia en la producción primaria y secundaria, sobre todo en sistemas donde los nutrientes disponibles para la vegetación escasean, como ocurre en los ecosistemas de pastizales.
El presente trabajo tuvo como objetivo determinar la descomposición de la hojarasca y su relación con algunos de los factores bióticos y abióticos presentes en un pastizal de Panicum maximum.
MATERIALES Y MÉTODOS
Las investigaciones se realizaron en la Estación Experimental de Pastos y Forrajes ¨Indio Hatuey¨, situada entre los 22°, 48' y 7" de latitud Norte y los 81° y 2' de longitud Oeste, a 19,01 msnm, en el municipio de Perico, provincia de Matanzas, Cuba (Academia de Ciencias de Cuba, 1989).
El suelo donde se llevó a cabo la fase experimental se clasifica como Ferralítico Rojo lixiviado (Hernández et al., 1999).
La investigación se inició en febrero del 2004 y se extendió hasta septiembre de ese año. La descomposición de la hojarasca se determinó como la pérdida de biomasa a través del tiempo, con relación al peso inicial (Liu, Fox y Xu, 2000).
Para el estudio de la dinámica de la descomposición se utilizó el método de bolsas de hojarasca (litter bags) de Caldentey, Ibarra y Hernández (2001). Las bolsas medían 10 x 10 x 10 cm, con poros de 1 cm de diámetro, lo cual permite el acceso, al interior, de un amplio rango de la biota edáfica. Se distribuyeron al azar 40 bolsas que contenían hojarasca de P. maximum. En cada bolsa se colocaron 20 g de hojarasca (base seca) previamente colectada. Las bolsas se enumeraron y se colocaron de manera que toda su superficie estuviera en contacto con el horizonte orgánico y se fijaron al suelo mediante estacas metálicas.
Se escogieron al azar cuatro bolsas a los 30, 60, 90, 120, 150, 180 y 210 días de situadas en el pastizal. En cada fecha de recolección, a la hojarasca remanente de cada bolsa se le determinó la población de macrofauna (organismos con diámetro mayor que 2 mm) mediante la separación manual, según la metodología del Programa de Investigación Internacional ¨Biología y Fertilidad del Suelo Tropical¨(TSBF), propuesta por Anderson e Ingram (1993), y se calculó el valor promedio de la densidad (individuos por metro cuadrado), así como la abundancia proporcional (%) para cada taxón. La densidad se determinó a partir del número de individuos y la abundancia relativa mediante la relación entre la cantidad de individuos que pertenecen a un grupo taxonómico y el total de individuos de todos los grupos taxonómicos.
Posteriormente, cada muestra se lavó con agua destilada y desionizada y se puso a secar en estufa a 60°C hasta peso constante. Se calculó la diferencia entre el peso inicial y el peso remanente. La muestra seca se molió a tamaño de partícula menor que 1 mm y se le determinó N, P y Ca en base seca, según las técnicas de la AOAC (1995). Se analizaron las fracciones de la fibra (fibra ácido detergente, fibra neutro detergente, lignina, celulosa, el N unido a la fibra neutro detergente, N-FND y a la fibra ácido detergente, N-FAD) de acuerdo con Van Soest y Wine (1968). La hemicelulosa se calculó mediante la diferencia de la FND-FAD y el contenido celular mediante 100-FND (Herrera, 1981).
Durante el proceso de descomposición se registró diariamente el comportamiento de la temperatura, la humedad relativa, las precipitaciones y la distribución de los días con lluvias.
Análisis matemático. Se realizó análisis de varianza según modelo lineal de clasificación simple. Se aplicó la dócima de Duncan (1955) en los casos necesarios.
Se comprobaron los supuestos de normalidad de los errores por la prueba modificada de Shapiro Wilk (Royston, 1982), así como la homogeneidad de la varianza según la prueba de Bartlex (1937) para verificar la normalidad de los datos y para la uniformidad de la varianza; en los casos de no cumplirse, se transformó la variable según «x.
Se utilizó el análisis de correlación y regresión lineal y múltiple para conocer la interrelación entre las variables y los modelos de mejor ajuste. Se consideró como variables independientes: los días, los factores climáticos y la frecuencia acumulada de la macrofauna; y como variables dependientes la composición química de la hojarasca remanente, el porcentaje de biomasa perdida y el peso de la hojarasca residual.
Para el procesamiento de la información se utilizó el software estadístico INFOSTAT 2001, versión 1.
RESULTADOS Y DISCUSIÓN
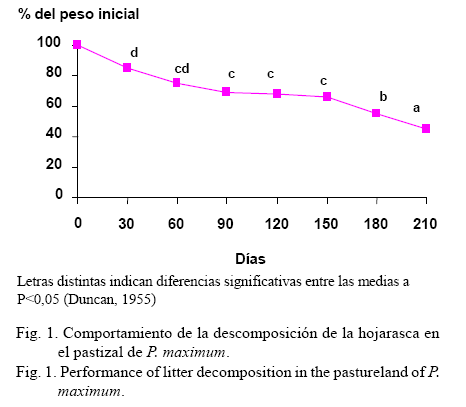
La disminución del peso de la hojarasca en este pastizal fue significativa entre las diferentes fechas de descomposición, aunque a los 210 días de colecta la hojarasca aún presentaba el 45,3% de su peso inicial (fig. 1) y la mayor tasa de descomposición ocurrió en los primeros 30 días (tabla 1).
Esta mayor tasa de descomposición en las primeras etapas coincide con lo informado por Martín (1995), quien señaló que el ciclo de descomposición de la hojarasca consta de tres etapas: 1) biodegradación rápida de la mayoría de los compuestos hidrosolubles y polisacáridos, debido a la acción microbiana y a los pluviolavados que ocurren en los primeros 20 ó 30 días; b) disminución lenta de los compuestos hidrosolubles fenólicos y las hemicelulosas por la fragmentación, el transporte, la mezcla y la biodegradación de la hojarasca, debido al ataque microbiano y faunístico; y c) aumento en el contenido de ligninas y proteínas, por las transformaciones húmica y mineral con el lavado de los hidrosolubles neoformados. Por ello la velocidad de descomposición decrece en el tiempo, debido a que las sustancias más fáciles de descomponer se agotan primero y queda posteriormente un sustrato lígnico más biorresistente.
La tasa media de descomposición de la hojarasca de esta gramínea (0,026 g-1día-1) fue cercana a lo encontrado por otros autores en la hojarasca de otras gramíneas.
Así Sandoval (2006), en pastizales de Brachiaria brizantha, obtuvo una tasa de 0,02 g g-1 día-1 durante el período evaluado (nueve meses); mientras que Boddey, Macedo, Tarré, Ferreira, Oliveira, Rezende, Cantauritti, Pereira, Alves y Urquiaga (2004) encontraron una tasa de 0,030 g g-1 día-1 en el estudio de reciclaje de N en pastizales de Brachiaria humidicola en Brasil, con un clima tropical húmedo.
La correlación entre el porcentaje de biomasa perdida y la composición química de la hojarasca remanente (tabla 2) mostró un mayor valor con el N-FND y la lignina. Precisamente estos dos indicadores se señalan por Melillo, Aber y Muratore (1982) y Tian (1992) como materiales de difícil descomposición, por lo que en la medida que disminuyen los compuestos más lábiles, se produce un aumento en la concentración de los más recalcitrantes.
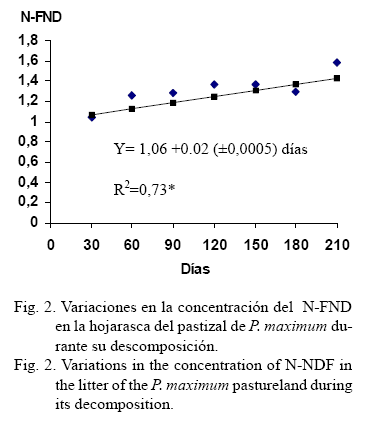
Al tomar en consideración las mayores correlaciones encontradas con las concentraciones de lignina y de N-FND, se determinó la variación que experimentaron estos indicadores durante el proceso de descomposición de la hojarasca. En este sentido, solo se encontró que la concentración de N-FND mostró ajuste significativo (R2 =0,73, P< 0,005) al modelo lineal (fig.2) e indica que a medida que transcurrió el proceso de descomposición se incrementó la concentración de este indicador.
Estos cambios pudieran ser la respuesta al efecto que produce la fauna del suelo en la hojarasca durante su proceso de descomposición,manifestado en: la degradación bioquímica producida por los microorganismos, las estrategias alimentarias que se establecen entre algunos invertebrados y microorganismos, y la acción de los excrementos ricos en minerales de la mayoría de los invertebrados; aunque también se ha encontrado que en la medida que transcurre el proceso de descomposición, se agotan las sustancias de más fácil ataque microbiano y quedan en la hojarasca remanente aquellas sustancias de difícil descomposición. No obstante, será necesario profundizar en el conocimiento de las causas del aumento de la concentración de determinados compuestos químicos en la hojarasca, en la medida que se produce su proceso de descomposición.
En la literatura se citan aumentos en la concentración de diversos indicadores de la composición química presentes en la hojarasca durante el proceso de descomposición. Por ejemplo, Edmonds y Thomas (1995) y Badejo, Nathaniel y Tian (1998) encontraron aumentos de la concentración de lignina, taninos, celulosa, hemicelulosa, nitrógeno y carbono. Estos cambios se relacionan con la colonización y la actividad de la fauna descomponedora (Hunter, Adl, Pringle y Coleman, 2003; Barajas-Guzmán y Álvarez-Sánchez, 2003).
Para determinar el efecto de los factores climáticos en el proceso de descomposición se estudiaron diferentes modelos. En este sentido, los que explicaron con mayor bondad de ajuste esta relación (tabla 3) presentaron elevados coeficientes de determinación (R2); sin embargo, el modelo que contempla la distribución de las lluvias (días con lluvias) como variable independiente mostró el mayor valor. Esta acción marcada de las lluvias en el proceso de descomposición puede deberse tanto a su acción directa en la fragmentación de la hojarasca, como en la provisión de humedad adecuada del sustrato, que unido a la acción de la temperatura puede ofrecer condiciones más favorables para la actividad de la biota responsable de la descomposición (Smith y Bradford, 2003).
Vanlauwe, Sanginga y Merckx (1997) encontraron que la pérdida de biomasa tuvo mejor correlación con el número de días con lluvias que con la cantidad total de lluvias caídas, aunque estos resultados se basaron en regímenes de lluvias aplicados artificialmente y no se tomó en consideración el efecto combinado de otras variables climáticas.
En la presente investigación resulta interesante destacar que cuando se consideraron los factores climáticos combinados, como la temperatura, la humedad relativa y la precipitación (tabla 4), el modelo mostró el mejor ajuste, de modo que su uso pudiera contribuir a predecir, con determinada exactitud, la hojarasca que se pierde en el pastizal en cierto período de tiempo. Por ello es preciso comprobar este modelo en pastizales en otras condiciones ambientales.
La composición taxonómica de la macrofauna asociada a la descomposición de la hojarasca constituye otro aspecto de vital importancia para la comprensión de este proceso en los pastizales. La caracterización de la macrofauna en el pastizal de P. maximum aparece en la tabla 5 y estuvo constituida por dos phylum, cinco clases, nueve órdenes, siete géneros y siete especies. El phylum Arthropoda resultó el más representado, con cuatro clases, y dentro de ellas la clase Insecta fue la más representada. La comunidad estuvo constituida por el 77% de organismos detritívoros y 11,11% de herbívoros y depredadores.
La presencia de organismos detritívoros resulta de gran importancia, pues su función principal, según Stork y Egglenton (1992), es el reciclaje de los nutrimentos, debido a que son consumidores que se alimentan del detritus y de las poblaciones de microorganismos asociados a él.
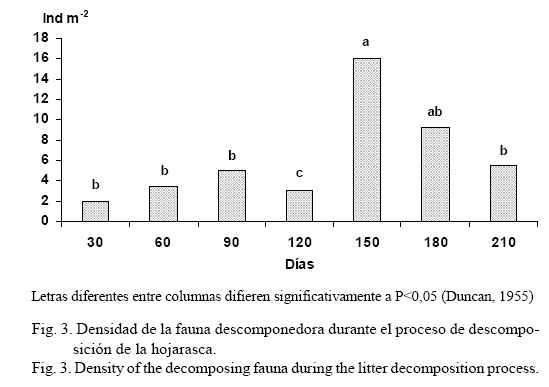
La mayor densidad de invertebrados se encontró en la hojarasca remanente a los 150 días (fig. 3), lo cual debe estar asociado con las complejas interacciones de los factores que influyen durante el proceso de descomposición, entre ellos los elevados valores de la temperatura media y la mayor cantidad de días con lluvias que ocurrieron en dicho período de tiempo (tabla 6). No obstante, pueden existir otros factores no estudiados en el presente trabajo que pudieran haber incidido también en este comportamiento, tales como, la variación en las concentraciones de las sustancias en la hojarasca remanente y las interacciones de diferente índole que se presentan en la cadena trófica entre las comunidades de la biota. De ahí la necesidad de profundizar en la investigación sobre la caracterización del resto de los organismos que participan en la descomposición del detritus vegetal.
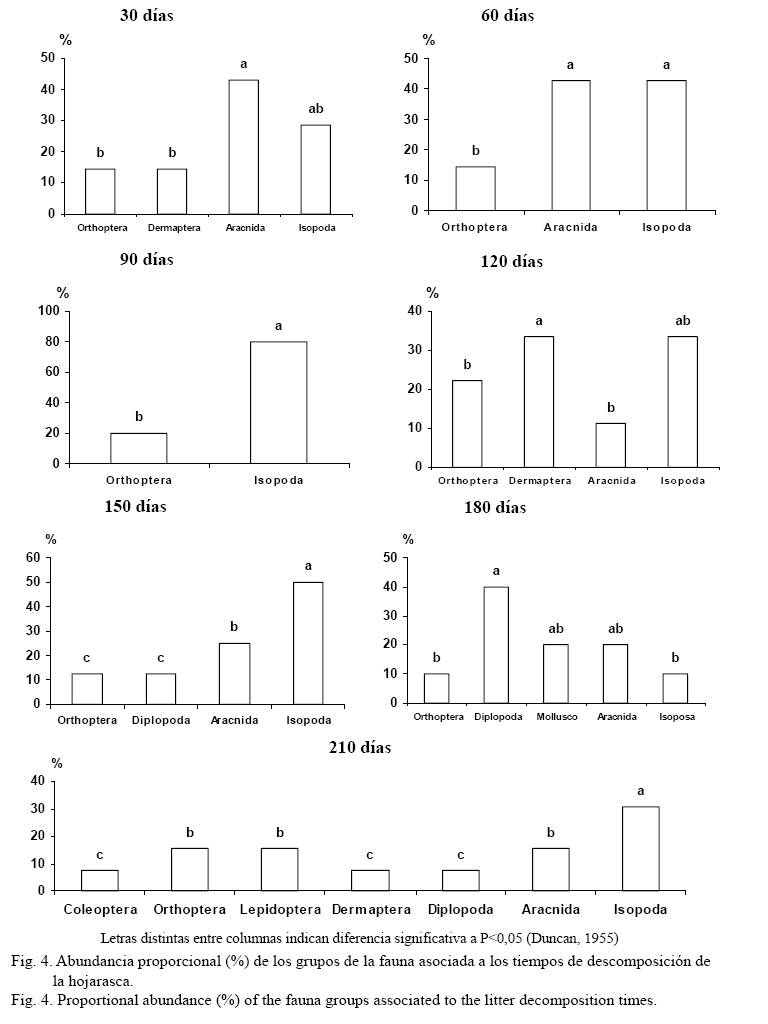
Los organismos de la macrofauna que participaron en el proceso de descomposición de la hojarasca en el pastizal de P. maximum se muestran en la figura 4. La presencia de los órdenes varió con los días de descomposición. La preferencia de los grupos faunísticos en cada etapa mostró un proceso dinámico, donde se manifestó la sucesión de organismos como respuesta a los requerimientos propios de cada uno de ellos. En este sentido, existen poblaciones que solo acuden a la hojarasca como nido o refugio cuando las condiciones de humedad y temperaturas son idóneas, y otras por la preferencia de algún tipo específico de alimento de origen vegetal, animal o fúngico (Ayres, Dromph y Bardgett, 2006).
Es importante señalar que los órdenes Isopoda y Ortoptera, representados principalmente por las especies Venezillo sp., Nagarus sp. y Pycnocelus surinamensis, fueron los de mayor presencia durante todo el proceso. Se considera que ambos grupos son dependientes de la hojarasca, pues constituye su principal fuente de alimento, lo que sugiere que la presencia de ellos durante todo el proceso influye en la descomposición de la materia orgánica en el sistema (Cabrera, 2003). Otros individuos, como los arácnidos y los dermápteros, se presentaron indistintamente en cualquier etapa del proceso, lo cual requiere investigaciones futuras.
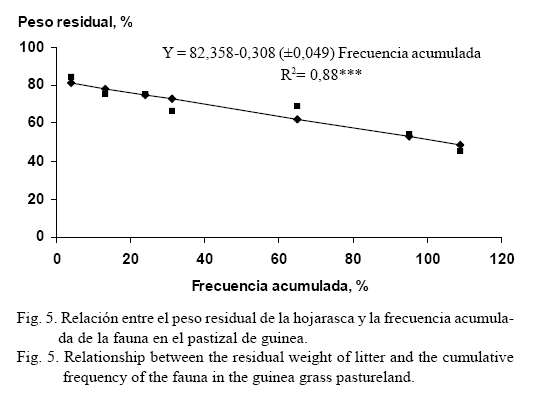
La figura 5 muestra la regresión lineal (R2= 0,88, P<0,01) entre el incremento de la frecuencia acumulada de la macrofauna y el peso remanente de la hojarasca durante el proceso de descomposición, e indica que en la medida que fue mayor la frecuencia de aparición de estos individuos resultó menor el peso de la hojarasca residual. Este efecto puede estar relacionado con el desmenuzamiento mecánico que produce esta fauna en la hojarasca, lo que también debe facilitar la acción de otros organismos que participan en el proceso de descomposición. En investigaciones realizadas por Attignon, Weibel, Lachat, Sensin, Nagel y Peveling (2004) en otras condiciones climáticas y especies de plantas diferentes, se encontró una tendencia similar; mientras que Ouédraogo, Mando y Brussaard (2004) informaron altas correlaciones entre la frecuencia acumulada de organismos (principalmente las termitas) y la disminución del peso remanente de la hojarasca en sistemas agrícolas de bajos insumos en África.
De forma general, los resultados demostraron que el proceso de descomposición de la hojarasca de P. maximum fue lento, ya que a los 210 días la hojarasca aún presentaba el 45,3% de su peso inicial, con una mayor tasa de descomposición en los primeros 30 días En el material remanente la concentración del N-FND se incrementó. La composición taxonómica de la fauna asociada a dicho proceso estuvo constituida por dos phylum, cinco clases, nueve órdenes, siete géneros y siete especies determinadas. La presencia de una diversa y estable fauna asociada a las bolsas influyó en el proceso de descomposición de la hojarasca. Las especies Venezillo sp., Nagarus sp. y Pycnocelus surinamensis fueron las de mayor presencia durante todo el proceso. El modelo matemático que contempla el efecto de la temperatura, la humedad relativa y la precipitación, mostró el mejor ajuste en el proceso de descomposición de la hojarasca en este pastizal.
REFERENCIAS BIBLIOGRÁFICAS
1. Academia de Ciencias de Cuba. 1989. Nuevo Atlas Nacional de Cuba. Instituto Cubano de Geodesia y Cartografía. La Habana, Cuba. p. 41
2. Aerts, R. & De Caluwe, H. 1997. Nutritional and plant mediated controls on leaf litter decomposition of Carex species. Ecology. 78:244
3. Álvarez-Sánchez, J. & Becerra, R. 1996. Leaf decomposition in a Mexican tropical rain forest. Biotropica. 28:657
4. Anderson, J.M. & Ingram, J. (Eds). 1993. Tropical soil biology and fertility. A handbook of methods. 2nd edition. CAB International. Wallingford, UK. 221 p.
5. AOAC. 1995. Official methods of analysis. Ass. Off. Agric. Chem. Washington, D.C.
6. Attignon, S.E.; Weibel, D.; Lachat, T.; Sensin, B.; Nagel, P. & Peveling, R. 2004. Leaf litter breakdown in natural and plantation forest of the Lama forest reserve in Benin. Applied Soil Ecology. 27:109
7. Ayres, E.; Dromph, K.M. & Bardgett, R.D. 2006. Do plant species encourage soil biota that specialise in the rapid decomposition of their litter?. En: http:// www.elsevier.com/locate/soilbio. [Fecha de consulta: 21 de junio 2007]
8. Badejo, M.A.; Nathaniel, T.H. & Tian, G. 1998. Abundance of springtails (Collembola) under four agroforestry trees species with contrasting litter quality. Biology and Fertility of Soil. 27:15
9. Barajas-Guzmán, G. & Álvarez-Sánchez, J. 2003. The relationships between litter fauna and rates of litter decomposition in a tropical rain forest. Applied Soil Ecology. 24:91
10. Bardgett, R.D. & Shine, A. 1999. Linkages between plant litter diversity, soil microbial biomass and ecosystem functioning temperate grasslands. Soil Biology & Biochemistry. 31:317
11. Bartlex, M.S. 1937. Properties of sufficiency and statistical test. Proceedings of the Royal Society of London, Sr. A. 160:268
12. Berg, B. 2000. Litter decomposition and organic matter turnover in northern forest soils. Forest Ecology and Management. 13:133
13. Boddey, R.M.; Macedo, R.; Tarré, R.M.; Ferreira, E.; Oliveira, O.C.; Rezende, C.; Cantauritti, R.B.; Pereira, J.M.; Alves, B.J. & Urquiaga, S. 2004. Nitrogen cycling in Brachiaria pastures: the key to understanding the process of pasture decline. Agriculture, Ecosytems and Environment. 103:389
14. Cabrera, G. de la C. 2003. Caracterización de la macrofauna del suelo en áreas con manejo agroecológico de Cangrejeras, La Habana, Cuba. Tesis en opción al Titulo Académico de Master en Ecología y Sistemática Aplicada. Instituto de Ecología y Sistemática, Ministerio de Ciencia, Tecnología y Medio Ambiente, Ciudad de La Habana, Cuba. 36 p.
15. Caldentey, J.; Ibarra, M. & Hernández, J. 2001. Litter fluxes and decomposition in Nothofagus pumilio stands in the region of Magallanes, Chile. Forest Ecology and Management. 148:145
16. Edmonds, R.L. & Thomas, T.B. 1995. Decomposition and nutrient release from green needles of western hemlock and Pacific silver fir in an old-growth temperate rain forests, Olympic National Park Washington. Can. J. For. Res. 25:1049
17. González, I.M. & Gallardo, J.F. 1995. El efecto de la hojarasca: una revisión. En: Anales de Edafología y Agrobiología. Centro de Edafología y Biología Aplicada. Salamanca, España. p. 86
18. Hernández, A.; Pérez, J.M.; Bosch, D.; Rivero, L.; Camacho, E.; Ruíz, J.; Salgado, E.J.; Marsán, R.; Obregón, A.; Torres, J.M.; González de la Torre, J.E.; Orellana, R.; Paneque, J.; Nápoles, P.; Fuentes, E.; Durán, J.L.; Peña, J.; Cid, G.; Ponce de León, D.; Hernández, M.; Frómeta , E.; Fernández, L.; Carcés, N.; Morales, M.; Suárez, E.; Martínez, E. & Ruíz de León, J.M. 1999. Nueva versión de la clasificación genética de los suelos de Cuba. Ministerio de la Agricultura. La Habana, Cuba. p. 26
19. Herrera, R.S. 1981. Influencia del fertilizante nitrogenado y la edad del rebrote en la calidad del pasto Bermuda cruzada (Cynodon dactylon vc Coast cross 1). Tesis de Doctor en Ciencias. Instituto de Ciencia Animal. La Habana, Cuba. p. 145
20. Hunter, M.D.; Adl, S.; Pringle, C.M. & Coleman, D.C. 2003. Relative effects of macroinvertebrates and habitat on the chemistry of litter during decomposition. Pedobiología. 47:101
21. InfoStat. 2001. Software estadístico. Manual de usuario. Versión 1. Córdoba, Argentina
22. Liu, W.; Fox, J.E.D. & Xu, Z. 2000. Leaf litter decomposition of canopy trees, bamboo and moss in a montane moist evergreen broad-leaved forest on Ailao Mountain, Yunnan, south-west China. Ecol. Res. 15:435
23. Martín, A.E. 1995. Reciclado de bioelementos a través de la hojarasca en ecosistemas forestales de la Sierra de Gata. Sistema Central Español. Universidad de Salamanca. Facultad de Ciencias Químicas, España.
24. Melillo, J.M.; Aber, J.D. & Muratore, J.F. 1982. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology. 63:621
25. Ouédraogo, E.; Mando, A. & Brussaard, L. 2004. Soil macrofaunal-mediated organic resource disappearance in semi-arid West Africa. Applied Soil Ecology. 27:259
26. Royston, P. 1982. An Extension of Shapiro and wilk's test for normality to large samples. Journal of the Royal Statistical Society: Series C (Applied Statistics). 31:115
27. Sandoval, I.E. 2006. Producción de hojarasca y reciclaje de nutrientes de dos especies arbóreas y dos gramíneas en pasturas de Muy, Muy, Nicaragua. Tesis para optar por el grado de Magister Scientiae en Agroforestería Tropical. CATIE. Turrialba, Costa Rica. p. 160
28. Smith, V.C. & Bradford, M.A. 2003. Litter quality impacts on grassland litter decomposition are differently dependent on soil fauna across time. Applied Soil Ecology. 24:197
29. Stork, N.E. & Egglenton, P. 1992. Invertebrates as determints and indicator of soil quality. Am. J. Altern. Agric. 7:38
30. Tian, G. 1992. Biological effects of plant residues with contrasting chemical compositions on plant soil under humic tropical conditions. PhD Thesis. Wageningen Agriculture University, The Netherlands
31. Vanlauwe, B.; Sanginga, N. & Merckx, R. 1997. Decomposition of four Leucaena y Senna prunings in alley cropping systems under sub-humid tropical conditions. The process and its modifiers. Soil Biology and Biochemistry. 29:131
32. Van Soest, P.J. & Wine, R.H. 1968. Determination of lignin and cellulose in aciddetergent ûbre with permanganate. Journal of the Association of Ofûcial Agricultural Chemists. 51:780
33. Vitousek, P.M.; Turner, D.R.; Parton, W.J. & Souford, R.L. 1994. Litter decomposition on the Manua Loa environmental matrix, Hawaii: patterns, mechanisms and models. Ecology. 75:418
Recibido el 15 de septiembre del 2007.
Aceptado el 3 de octubre del 2007.
Saray SánchezI, G. CrespoII y Marta Hernández I.
I Estación Experimental de Pastos y Forrajes ¨Indio Hatuey¨ Central España Republicana, CP 44280, Matanzas, Cuba
E-mail:saray.sanchez@indio.atenas.inf.cu
IIInstituto de Ciencia Animal, La Habana, Cuba

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}