










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.31 n.4 Matanzas oct.-dic. 2008
ARTÍCULO DE INVESTIGACIÓN
Selección de rizobios adaptados a ecosistemas ganaderos de Alberta, Canadá, inoculados en maíz (Zea mays, L.). Fase I: invernadero
Selection of rhizobia adapted to livestock production ecosystems from Alberta, Canada, inoculated in corn (Zea mays, L.). Stage I: greenhouse
C.J. Bécquer1, Beatriz Salas1, D. Archambault2, J. Slaski2 y A. Anyia2
1Estación Experimental de Pastos y Forrajes Sancti Spíritus. Apdo. 2228, Zona postal 1, Sancti Spiritus, Cuba
mailto: becquer@pastos.yayabo.inf.cu
2Environmental Technologies Division. Alberta Research Council, Vegreville, Alberta, Canada
RESUMEN
Se efectuó un ensayo en invernadero con el objetivo de seleccionar cepas de rizobio inoculadas en maíz (Zea mays, L.), para lo cual se utilizaron 16 cepas nativas canadienses de Sinorhizobium meliloti, así como 12 cepas comerciales, pertenecientes a diferentes géneros y especies de rizobio. Los inóculos se confeccionaron en medio CLM y fueron agitados hasta alcanzar un título de 106-108 cél./mL. Se evaluaron las variables: peso seco aéreo (PSA), peso seco raíz (PSR) y longitud del tallo (LT). Se constató que en todas las variables existieron tratamientos inoculados que fueron estadísticamente superiores al control absoluto. En el PSA dos tratamientos no difirieron significativamente del control fertilizado; mientras que sólo cuatro tratamientos inoculados en LT y cinco en PSR fueron estadísticamente inferiores al control fertilizado. La agrupación en conglomerados permitió seleccionar las cepas CAC8, CAC9, CAC17 y CAC17a, para su posterior evaluación en condiciones de campo. El 100% de los tratamientos seleccionados en el dendrograma fueron inoculados con cepas nativas. Al evaluar los tratamientos en cuanto al nivel de dependencia del PSA y la LT con respecto al PSR, se observó una regresión fuerte del PSA en los inoculados con cepas nativas, y débil en los inoculados con cepas comerciales, así como en los controles. La regresión fue débil en la LT en todos los tratamientos. Se recomienda la continuación de estas investigaciones en condiciones de campo, además de otros estudios más profundos.
Palabras clave: Inoculación, Rhizobium, Sinorhizobium, Zea mays
ABSTRACT
A greenhouse experiment was conducted in order to select rhizobium strains inoculated on corn (Zea mays, L.), for which 16 Canadian native strains of Sinorhizobium meliloti, as well as 12 commercial strains, belonging to different rhizobium genera and species, were used. The inoculi were prepared in BYM medium and shaken to obtain a titer of 106-108 cells/mL.. The variables evaluated were: aerial dry weight (ADW), root dry weight (RDW) and stem length (SL). In all the variables some inoculated treatments were observed to be statistically higher than the absolute control. In ADW two treatments did not differ significantly from the fertilized control; while only four inoculated treatments in SL and five in RDW were statistically lower than the fertilized control. Cluster analysis allowed to select the strains CAC8, CAC9, CAC17 and CAC17a, for their later evaluation under field conditions. One hundred percent of the treatments selected in the dendrogram were inoculated with native strains. When evaluating the treatments regarding the level of dependence of ADW and SL with regards to RDW, a strong regression of ADW was observed in the ones inoculated with native strains, and a weak regression in the treatments inoculated with commercial strains. The regression was weak in SL for all treatments. To continue this research under field conditions, in addition to further studies, is recommended.
Key words: Inoculation, Rhizobium, Sinorhizobium, Zea mays
INTRODUCCIÓN
El efecto beneficioso de los rizobios como fijadores del nitrógeno atmosférico en las leguminosas a través de la simbiosis es bien conocido, por lo que son utilizados ampliamente en el mundo para el incremento de la producción de grano y forraje, lo cual contribuye al ahorro monetario y al cuidado y preservación del medio ambiente. Estas bacterias pueden colonizar también las raíces de plantas no leguminosas e influir significativamente en su crecimiento, mediante la secreción de sustancias promotoras del crecimiento vegetal (Chabot, Antoun, Kloepper y Beauchamp, 1996). Acorde con lo expresado por Matiru y Dakora (2004), la infección rizobiana en las no leguminosas es más común en la naturaleza que lo que se pensaba anteriormente. A pesar de que en varias instituciones en el extranjero se han realizado estudios con rizobios en diferentes especies, en Cuba solo se conocen los beneficios de la inoculación en plantas de maíz, con cepas nativas de radyrhizobium sp. procedentes de los ecosistemas ganaderos de Sancti Spiritus (Bécquer, Salas, Archambault, Slaski y Anyia, 2008), por lo que se considera que esta investigación es novedosa para el país.
El objetivo de este trabajo fue estudiar el efecto de la inoculación de rizobios nativos canadienses y comerciales en maíz, y así realizar una selección de las cepas más promisorias para su posterior evaluación en condiciones de campo, ya sea en el propio Canadá o en Cuba.
MATERIALES Y MÉTODOS
Identificación de las cepas de rizobio. Se utilizaron 16 cepas que se aislaron e identificaron a partir de nódulos radiculares de leguminosas naturalizadas de Alberta, Canadá (Melilotus officinalis y Medicago sativa) (Bécquer, C.J., inédito); estas fueron confirmadas como pertenecientes al género Sinorhizobium meliloti (de Lajudie et al., 1994). También se usaron 12 cepas comerciales pertenecientes a diferentes géneros y especies de rizobio (tabla 1).

Preparación de los inóculos. Los inóculos se confeccionaron en medio CLM (caldo-levadura-manitol) (Somasegaran y Hoben, 1994), a partir de una asada de cultivos frescos en medio agarizado, y se colocaron en agitación (120 rpm) durante siete días (para las especies de lento crecimiento) y de tres a cuatro días (especies de rápido crecimiento), en una zaranda orbital a 30oC, hasta alcanzar un título UFC (unidad formadora de colonias) de 106-108 cél./mL.
Ensayos de invernadero. Se escogió la variedad de maíz 2338 F15 donada por el Alberta Research Council, Canadá. Las semillas fueron pregerminadas previamente a su siembra en macetas. Cada maceta contenía 1,0 kg de suelo estándar mezclado con bajos niveles de N: 30 ppm (mg/kg). Se raleó a los cinco días de la siembra hasta dejar dos plantas por maceta. Se utilizaron los métodos estándar para la inoculación de los cereales (Sabry, Saleh, Batchelor, Jones, Jotham, Webster, Kothari, Davey y Cocking, 1997; Biswas, Ladha y Dazzo, 2000), con cambios en la frecuencia y la cantidad de inóculo. El inóculo bacteriano en cantidad de 3 mL/planta, que contenía 106-108 cél./mL, se aplicó a los cinco días de la siembra; de forma consecutiva, se realizaron otras tres inoculaciones con el mismo título (6 mL/planta, 12 mL/planta y 12 mL/planta) a los 20, 30 y 40 días de la siembra, respectivamente. Se evaluaron las variables peso seco aéreo (PSA), peso seco raíz (PSR) y longitud del tallo (LT) a los 60 días de la siembra. Se calculó la regresión entre las variables PSA, PSR y LT, en los tratamientos inoculados con cepas nativas y en los inoculados con cepas comerciales, así como en los controles.
Diseño experimental y análisis estadístico. Se empleó un diseño en bloque al azar con 28 tratamientos y cuatro réplicas (Lerch, 1976). Se incluyó un control absoluto y uno con un portador nitrogenado (NH4NO3, 150 ppm N/kg), además de un control al que se le añadió CLM (5 mL/planta), a los 5 días de la siembra; así como 10 mL/planta, 15 mL/planta y 20 mL/planta, a los 20, 30 y 40 días de la siembra, respectivamente. Se realizó un ANOVA mediante el paquete estadístico StatGraphics Plus, versión 5.1, 1994-2001, Statistical Graphics Corporation. Las diferencias entre medias se hallaron mediante la prueba de LSD de Fisher (p<0,05; 95% conf.). Se construyó un dendrograma mediante el método de Ward (distancia métrica City-Block).
RESULTADOS Y DISCUSIÓN
En la tabla 2 se muestran los resultados en cada variable. En el peso seco aéreo se constató que seis tratamientos inoculados con cepas nativas (CAC5, CAC8, CAC9, CAC16, CAC17 y CAC17a) fueron estadísticamente superiores (p<0,05) al control absoluto, lo cual representa un 18,6% de incremento en CAC5; 22,3% en CAC8; 20,0% en CAC9, 19,2% en CAC16; 25,0% en CAC17 y 20,9% en CAC17a. Ello puede deberse a que los rizobios producen fitohormonas que promueven el crecimiento vegetal y posiblemente incrementen los rendimientos (Matiru y Dakora, 2004). La producción de fitohormonas por los rizobios, según Okon y Kapulnik (1986); Zavalin, Kandaurova y Vinogradova (1998) y Saubidet, Fatta y Barneix (2000), favorece una mayor extracción de nutrientes en el suelo.
Similares resultados obtuvieron Bécquer et al. (2008) en maíz, al aplicar cepas de rizobio procedentes de ecosistemas ganaderos de Sancti Spíritus, Cuba. Sin embargo, estos valores fueron superiores a los encontrados por Prévost, Saddiki y Antoun (2000) en un experimento donde se inoculó maíz con cepas comerciales de Bradyrhizobium japonicum, en el cual obtuvieron incrementos en el PSA de la planta desde 6,7% hasta 8,7% con respecto al control absoluto. Yanni et al. (2001) hallaron que la cepa E11 de Rhizobium leguminosarum bv. trifolii fue capaz de estimular el crecimiento en maíz, en condiciones de campo. Los tratamientos inoculados con las cepas nativas CAC8 y CAC17 presentaron superíndices comunes con el control fertilizado. Excepto los tratamientos inoculados con las cepas comerciales NITRAGIN, MSDJ865 y USDA 191, los cuales fueron estadísticamente superiores al control absoluto, el resto presentaron letras comunes con el control que contenía CLM y con el control absoluto.
En la variable peso seco raíz los tratamientos CAC2, CAC4, CAC5, CAC6 C8, CAC9, CAC14, CAC17, MSDJ865 y USDA 191 presentaron diferencias significativas con respecto al control absoluto. En tal sentido, otros autores, como Feng Chi et al. (2005), determinaron que una cepa de rizobio con marcador molecular inoculada en arroz, incrementó significativamente el peso seco aéreo y la masa radicular de la planta. Todos los tratamientos fueron estadísticamente inferiores al control con el medio de cultivo CLM (p<0,05). Excepto CAC1, 25B6, NITRAGIN, ATCC 10317, ATCC 14480, ATCC 9930 y USDA 76, los cuales fueron estadísticamente inferiores, los demás tratamientos no difirieron del control fertilizado. Éste, a su vez, fue estadísticamente inferior al tratamiento con CLM. La razón de estos resultados podría atribuirse a que el CLM en estado puro (sin microorganismos) contiene macro y microelementos esenciales no metabolizados, importantes no sólo para la supervivencia de los rizobios, sino también para la nutrición de las plantas, lo cual lo convierte automáticamente en un segundo control fertilizado.
En cuanto a la longitud del tallo, los tratamientos con las cepas CAC8, CAC9, CAC17, CAC17a y ATCC 10004 fueron estadísticamente superiores (p<0,05) al control absoluto. Con excepción de los inoculados con las cepas CAC4, CAC6, CAC10 y NITRAGIN, los cuales fueron inferiores al tratamiento fertilizado, el resto presentó letras comunes con el control absoluto y el fertilizado. En esta variable, el tratamiento con CLM mostró valores estadísticamente inferiores al resto de los tratamientos.
En el dendrograma (fig. 1) se observan tres grandes conglomerados: el 1, compuesto por ocho tratamientos inoculados con cepas nativas y nueve con cepas comerciales; el 2, compuesto por cuatro tratamientos con cepas nativas y tres cepas comerciales; y el 3, formado por el control fertilizado, el control con CML y los tratamientos inoculados con las cepas nativas CAC8, CAC9, CAC17 y CAC17a; estos mostraron valores estadísticamente superiores en cada una de las variables y no difirieron del control fertilizado, por lo que se considera factible seleccionar estas cepas como altamente promisorias para futuros ensayos de campo.
No se debe descartar otros tratamientos, como los inoculados con las cepas CAC5, USDA 191 y MSDJ865, los cuales no se agruparon con los anteriormente señalados, pero sus valores fueron superiores significativamente al menos en dos variables.
En los gráficos de dispersión que se muestran a continuación, se puede observar que el nivel de dependencia entre el PSA y el PSR fue moderadamente fuerte (r2= 35,01%) para los tratamientos inoculados con las cepas nativas (fig. 2); mientras que en el caso de las cepas comerciales (fig. 3) esta relación fue relativamente débil (r2= 13,7%). Dicho resultado es la primera vez que se obtiene en experimentos de este tipo, ya que en estudios similares realizados por Bécquer, Salas, Archambault, Slaski y Anyia (2006) en plantas de trigo, con rizobios procedentes de Sancti Spíritus (Cuba), y por Bécquer et al. (2008), en plantas de maíz inoculadas con esas mismas cepas cubanas, se obtuvo una regresión débil entre estas variables en los tratamientos con cepas nativas. Los resultados actuales pudieran indicar un efecto inducido en la planta, con la secreción de hormonas de diferente naturaleza química por los rizobios nativos de Alberta, Canadá. El hecho de que las cepas cubanas no indujeran tal efecto pudo deberse quizás a que estas pertenecen al género Bradyrhizobium, el cual es extremadamente lento en su multiplicación; mientras que las cepas canadienses pertenecen a Sinorhizobium, género que se caracteriza por un rápido crecimiento y, por ende, una mayor velocidad en la colonización de las raíces. Se corrobora lo planteado por Lupwayi y Clayton (2004) acerca de que los rizobios pueden actuar como promotores del crecimiento vegetal y cambiar la fisiología y morfología de las raíces inoculadas, lo que resulta en una mayor absorción de N y otros nutrientes. El coeficiente de regresión entre estas dos mismas variables, pero de los tratamientos control (fig. 4), fue también débil (r2= 7,9%), por lo que se infiere que en este caso el incremento del desarrollo vegetal aéreo no dependió estadísticamente del incremento del desarrollo radical.

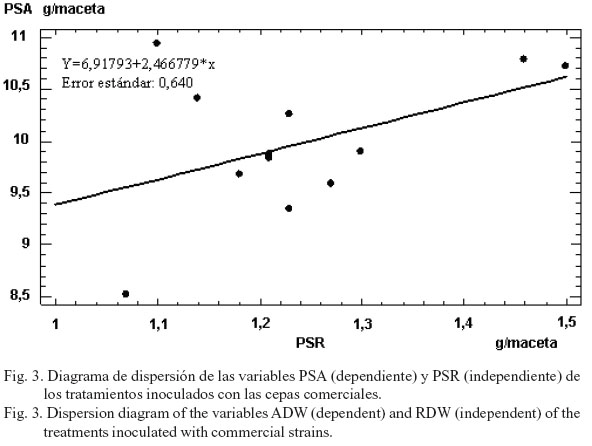
Es posible que al evaluar sólo tres variables de la planta, no se haya podido determinar una relación o dependencia más definitoria de la variable dependiente con otras independientes; aunque este aspecto, de índole puramente estadística, en opinión de los autores no le resta calidad a los resultados del experimento, ya que en los estudios de selección de cepas este número de variables es aceptado, siempre que refleje los indicadores fisiológicos más importantes de la planta para formar un criterio correcto de selección (Somasegaran y Hoben, 1994).
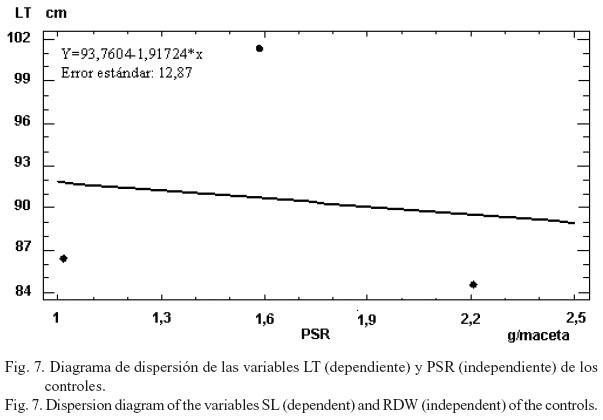
No se observó una regresión fuerte en las variables LT y PSR, tanto en los tratamientos inoculados con cepas nativas, como en los inoculados con cepas comerciales y en los controles (figs. 5, 6 y 7), lo que indica, de forma general, una mayor distribución de las fitohormonas hacia el desarrollo foliar.
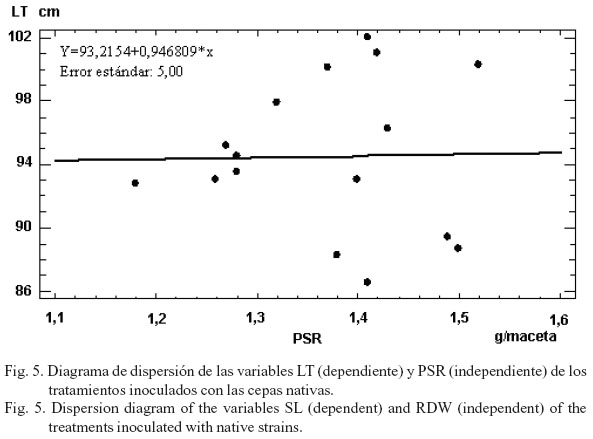

Se concluye que en todas las variables existieron tratamientos inoculados (con cepas nativas y comerciales) estadísticamente superiores al control absoluto. Además, se hallaron dos tratamientos en la variable peso seco aéreo (CAC8 y CAC17), así como 20 en peso seco raíz y 23 en longitud del tallo, los cuales no difirieron del testigo fertilizado. El 100% de los tratamientos seleccionados fueron inoculados con cepas nativas (CAC8, CAC9, CAC17, CAC17a). Existió una mayor correlación y un mayor grado de dependencia estadística entre el PSA y el PSR en el caso de las cepas nativas con relación a las comerciales. Los valores de la longitud del tallo no dependieron estadísticamente del PSR en ningún tratamiento.
Dadas estas observaciones, los autores recomiendan: efectuar ensayos de campo con las cepas seleccionadas, en variedades promisorias de maíz para la alimentación humana y animal, en Sancti Spiritus; realizar ensayos donde se inoculen diferentes genotipos de maíz para seleccionar los tratamientos de mejor interacción genotipo vegetal-cepa de rizobio; y finalmente estudiar las bases bioquímico-fisiológicas del efecto de los rizobios en la planta.
Agradecimientos
Al igual que en experimentos anteriores realizados con trigo, el financiamiento, las instalaciones, los equipos y los materiales utilizados para este trabajo, fueron provistos por Alberta Research Council (Vegreville, Alberta, Canadá), a través de un proyecto de colaboración con la Estación Experimental de Pastos y Forrajes Sancti Spíritus, Cuba. Los autores agradecen profundamente a todos los funcionarios de esta Institución canadiense, especialmente al Dr. Daniel Archambault y al Dr. Jan Slaski, por su gran esfuerzo en hacer realidad este proyecto.
REFERENCIAS BIBLIOGRÁFICAS
1. Bécquer, C.J.; Salas, Beatriz; Archambault, D.; Slaski, J. & Anyia, A. 2006. Inoculación de trigo (Triticum aestivum, L.) con rizobios adaptados a ecosistemas ganaderos de Sancti Spiritus, Cuba. Pastos y Forrajes. 29:255
2. Bécquer, C.J.; Salas, Beatriz; Archambault, D.; Slaski, J. & Anyia, A. 2008. Selección de rizobios adaptados a ecosistemas ganaderos de Sancti Spíritus, Cuba, inoculados en maíz (Zea mays, L.). Fase I: invernadero. Pastos y Forrajes. 31:229
3. Biswas, J.C.; Ladha, J.K. & Dazzo, F.B. 2000. Rhizobia inoculation improves nutrient uptake and growth of lowland rice. Soil Sci. Soc. of America J. 64:1644
4. Chabot, R.; Antoun, H.; Kloepper, J.W. & Beauchamp, Chantal. 1996. Root colonization of maize and lettuce by bioluminoscent Rhizobium leguminosarum biovar phaseoli. Appl. Environ. Microbiol. 62:2767
5. De Lajudie, P. et al. 1994. Polyphasic taxonomy of rhizobia: emendation of the genus Sinorhizobium and description of Sinorhizobium meliloti comb. Nov., Sinorhizobium saheli sp. Nov., and Sinorhizobium teranga sp. Nov. Int. J. Syst. Bacteriol. 44:715
6. Feng, Chi et al. 2005. Ascending migration of endophytic rhizobia, from roots to leaves, inside rice plants and assessment of benefits to rice growth physiology. Appl. Environ. Microbiol. 71:7271
7. Lerch, G. 1976. La experimentación en las ciencias biológicas y agrícolas. Ed. Ciencia y Técnica. La Habana, Cuba. 452 p.
8. Lupwayi, N.Z. & Clayton, G.W. 2004. Endophytic rhizobia and other diazotrophic bacteria in nonlegume crops. Recent Res. Devel. Crop Sci. 1:227
9. Matiru, Viviene & Dakora, F. 2004. Potential use of rhizobial bacteria as promoters of plant growth for increased yield in landraces of African cereal crops. African J. Biotecnol. 3:1
10. Okon, Y. & Kapulnik, Y. 1986. Development and functions of Azospirillum-inoculated roots. Plant Soil. 90:3
11. Prévost, D.; Saddiki, S. & Antoun, H. 2000. Growth and mineral nutrition of corn inoculated with effective strains of Bradyrhizobium japonicum. Proceedings of the 5th International PGPR Workshop. Villa Carlos Paz, Córdoba, Argentina. 7 p.
12. Sabry, S.R.S.; Saleh, S.A.; Batchelor, Caroline; Jones, J.; Jotham, J.; Webster, G.; Kothari, S.L.; Davey, M.R. & Cocking, E.C. 1997. Endophytic establishment of Azorhizobium caulinodans in wheat. Proc. R. Soc. Lond. B. 264:341
13. Saubidet, M.I.; Fatta, N. & Barneix, A.J. 2000. The effects of inoculation with Azospirillum brasilense on growth and nitrogen utilization by wheat plants. Plant Soil. 245:215
14. Somasegaran, P. & Hoben, H.J. 1994. Handbook for rhizobia. Springer-Verlag, New York. 450 p.
15. Yanni, Y.G. et al. 2001. The beneficial plant growth-promoting association of Rhizobium leguminosarum bv. trifolii with rice roots. Australian Journal of Plant Physiology. 28 (9):845
16. Zavalin, A.A.; Kandaurova, T.M. & Vinogradova, L.V. 1998. Influence of nitrogen fixing microorganisms on the nutrition and productivity of spring wheat and on the characteristics of photosynthesis of different varieties of spring wheat. In: Biological nitrogen fixation for the 21st Century. (Eds. Elmerich, C.; Kondorosi, A.; Newton, W.E.). Kluwer Academic Publishers. Dordrecht, The Netherlands. p. 413
Recibido el 8 de enero del 2008
Aceptado el 13 de junio del 2008

{kind=link}
{kind=link}
{kind=link}
{kind=link}