










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.32 n.3 Matanzas jul.-sep. 2009
ARTÍCULO DE INVESTIGACIÓN
Descomposición de la hojarasca en un sistema silvopastoril de Panicum maximum y Leucaena leucocephala (Lam) de Wit cv. Cunningham. I. Influencia de su composición química
Litter decomposition in a silvopastoral system of Panicum maximum and Leucaena leucocephala (Lam) de Wit cv. Cunningham. I. Influence of their chemical composition
Saray Sánchez1, G. Crespo2 y Marta Hernández1 1Estación Experimental de Pastos y Forrajes "Indio Hatuey". Central España Republicana, CP 44280, Matanzas, Cuba.
E-mail: saray.sanchez@indio.atenas.inf.cu
2Instituto de Ciencia Animal, La Habana, Cuba
RESUMEN
Con el objetivo de determinar la descomposición de la hojarasca y su relación con la composición química de Panicum maximum y Leucaena leucocephala en un sistema silvopastoril, se realizó este experimento en la EEPF "Indio Hatuey". La descomposición de la hojarasca se determinó como la pérdida de biomasa a través del tiempo, con relación al peso inicial. Para el estudio de la dinámica de la descomposición se utilizó el método de bolsas de hojarasca (litter bags). En cada especie se presentó un patrón diferente de descomposición de la hojarasca; la tasa promedio de descomposición de la hojarasca en leucaena fue mayor que en la guinea. En ambas especies se encontró una rápida pérdida de peso durante los primeros 30 días y después el proceso fue más lento. Este comportamiento puede estar relacionado con la composición química de los pastos, pues el porcentaje de biomasa perdida de la hojarasca de L. leucocephala presentó una mayor correlación con las concentraciones del contenido celular, la relación lignina/nitrógeno, la celulosa y el Nt. Se observó una dependencia significativa y negativa de la hojarasca de P. maximum con las concentraciones de la FND y la hemicelulosa; mientras que se relacionó de forma positiva con las de N-FND y la relación lignina/nitrógeno
Palabras clave: Composición química, hojarasca, Leucaena leucocephala, Panicum maximum
ABSTRACT
With the objective of determining the litter decomposition and its relationship to the chemical composition of Panicum maximum and Leucaena leucocephala in a silvopastoral system, this trial was conducted at the EEPF «Indio Hatuey». Litter decomposition was determined as biomass loss through time, with regards to initial weight. For studying the decomposition dynamics, the method of litter bags was used. In each species a different litter decomposition pattern appeared; the average litter decomposition rate was higher in leucaena than in Guinea grass. Rapid weight loss was found in both species during the first 30 days and afterwards the process was slower. This behavior could be related to the chemical composition of pastures, because the percentage of lost biomass in L. leucocephala litter showed a higher correlation to the cell content concentrations, lignin/nitrogen relationship, cellulose and Nt. A significant and negative dependence was observed between the P. maximum litter and the concentrations of NDF and hemicellulose; while it was positively related to the N-NDF concentrations and the lignin/nitrogen relationship.
Key words: Chemical composition, litter, Leucaena leucocephala, Panicum maximum
INTRODUCCIÓN
La descomposición de la hojarasca constituye la vía de entrada principal de los nutrientes en el suelo y es uno de los puntos clave del reciclado de la materia orgánica y los nutrientes (Vitousek et al., 1994; Aerts y De Caluwe, 1997). Varios autores han estudiado con detalle la dinámica de la descomposición de la hojarasca en plantas leñosas, tanto en climas templados como en el mediterráneo (González y Gallardo, 1995; Berg, 2000). Sin embargo, hay pocos estudios sobre dicha dinámica en pastizales (Bardgett y Shine, 1999), a pesar de su importancia en los sistemas silvopastoriles, por ser una alternativa viable para lograr la sostenibilidad ecológica y productiva de los pastizales tropicales.
La hojarasca determina la estabilidad y el funcionamiento del ecosistema, pues constituye la fuente principal de circulación de materia orgánica, energía y nutrientes entre las plantas y el suelo; su contenido depende del balance entre los procesos de acumulación y descomposición, lo cual está determinado por la especie de planta y su composición química, entre otros factores.
Por lo antes expuesto, el presente trabajo tuvo como objetivo determinar la descomposición de la hojarasca y su relación con la composición química de las principales especies vegetales presentes en un sistema silvopastoril de Panicum maximum y Leucaena leucocephala (Lam) de Wit cv. Cunningham.
MATERIALES Y MÉTODOS
La investigación se realizó en la Estación Experimental de Pastos y Forrajes «Indio Hatuey», situada entre los 22°, 48' y 7" de latitud Norte y los 81° y 2' de longitud Oeste, a 19,01 msnm, en el municipio de Perico, provincia de Matanzas, Cuba (Academia de Ciencias de Cuba, 1989).
El suelo donde se llevó a cabo la fase experimental se clasifica como Ferralítico Rojo lixiviado (Hernández et al., 1999).
La investigación se inició en febrero del 2004 y se extendió hasta septiembre de ese año. La descomposición de la hojarasca se determinó como la pérdida de biomasa a través del tiempo, con relación al peso inicial (Liu et al., 2000).
Para el estudio de la dinámica de la descomposición se utilizó el método de bolsas de hojarasca (litter bags), de Caldentey et al. (2001). Las bolsas medían 10 x 10 x 10 cm, con poros de 1 cm de diámetro, lo cual permite el acceso de un amplio rango de la biota edáfica hacia el interior. Se distribuyeron al azar 40 bolsas que contenían hojarasca de P. maximum y 40 de L. leucocephala. En cada bolsa se colocaron 20 g de hojarasca (base seca) previamente colectada. Las bolsas se enumeraron, se situaron de manera que toda su superficie estuviera en contacto con el horizonte orgánico y se fijaron al suelo mediante estacas metálicas.
Se escogieron al azar cuatro bolsas a los 30, 60, 90, 120, 150, 180 y 210 días de situadas en el pastizal. En cada fecha de recolección, la hojarasca remanente de cada bolsa se lavó con agua destilada y desionizada y se puso a secar en estufa a 60°C hasta peso constante. Se calculó la diferencia entre el peso inicial y el peso remanente. La muestra seca se molió a tamaño de partícula menor que 1 mm y se le determinó el N, el P y el Ca en base seca, según las técnicas de la AOAC (1995). Se analizaron las fracciones de la fibra (fibra ácido detergente, fibra neutro detergente, lignina, celulosa, el N unido a la fibra neutro detergente, N-FND y a la fibra ácido detergente, N-FAD) de acuerdo con Van Soest y Wine (1968). La hemicelulosa se calculó mediante la diferencia de la FND-FAD y el contenido celular mediante 100-FND (Herrera, 1981).
Análisis matemático. Se realizó un análisis de varianza según el modelo lineal de clasificación simple. Se aplicó la dócima de Duncan (1955) en los casos necesarios.
Se utilizó el análisis de correlación y regresión lineal y múltiple para conocer la interrelación entre las variables y los modelos de mejor ajuste. Se consideraron como variables independientes los días y como variables dependientes, la composición química de la hojarasca remanente y el porcentaje de biomasa perdida.
Para el procesamiento de la información se utilizó el software estadístico INFOSTAT (2001), versión 1.
RESULTADOS Y DISCUSIÓN
La dinámica de descomposición de la hojarasca de la leucaena y de la guinea en el sistema silvopastoril se muestra en la figura 1. La cantidad de material descompuesto fue mayor y ocurrió más rápido en L. leucocephala, con diferencias altamente significativas entre los días de descomposición. A los 210 días quedó sin descomponer solo el 3,12% de la hojarasca de leucaena; sin embargo, en similar tiempo la hojarasca de la guinea aún representaba el 28,2% del peso inicial.

Jamaludheen y Kumar (1997) describieron el proceso de descomposición de la hojarasca de la leucaena en la India, donde fue totalmente descompuesta a los ocho meses, y lo correlacionaron con su alto contenido de nitrógeno. Por su parte, Vanlauwe et al. (1997) señalaron que L. leucocephala, en condiciones subhúmedas tropicales, logró su total descomposición a los 112 días y este proceso fue más rápido que en la hojarasca de Senna siamea, la cual presentó una menor concentración de N.
Otros investigadores (Mtambanengwe y Kirchmann, 1995; Crespo et al., 2001) informaron patrones semejantes de la descomposición de la hojarasca, tanto de la gramínea como de la leguminosa, y señalaron que las diferencias marcadas en la velocidad de desaparición de la hojarasca entre las especies están estrechamente relacionadas con su composición química, ya que por lo general en las gramíneas las relaciones C/N y lignina/nitrógeno son mayores que en las leguminosas, lo cual hace más lenta la velocidad de descomposición.
La tasa promedio de descomposición de la hojarasca en leucaena fue mayor que en la guinea (tabla 1). En ambas especies se encontró una rápida pérdida de peso durante los primeros 30 días; después de este primer mes el proceso fue más lento. Este comportamiento fue hallado también por otros autores en diversos países (Wieder y Lang, 1982; King y Hutchinson, 1992; Aceñolaza y Gallardo, 1995; Saviozzi et al., 1995), quienes plantearon que puede ser explicado por la descomposición microbiana de sustancias fácilmente degradables y por el lavado de sustancias lábiles en las primeras etapas de descomposición de la hojarasca de numerosas plantas (Martín, 1995).
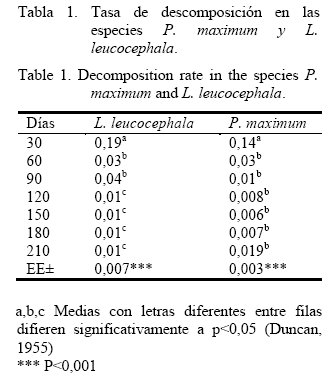
Estas tasas de descomposición de la hojarasca en pastizales son los primeros resultados que se obtienen en el país y pueden constituir el punto de partida para conocer este aspecto en otras especies de pastos y sistemas ganaderos.
El porcentaje de biomasa perdida de la hojarasca de L. leucocephala presentó una mayor correlación con las concentraciones del contenido celular (r= -0,98), la relación lignina/nitrógeno (r= 0,94), la celulosa (r= 0,91) y el Nt (R=-0,84). En cuanto a la hojarasca de P. maximum se observó una dependencia significativa y negativa con las concentraciones de la FND y la hemicelulosa; mientras que se relacionó de forma positiva con las de N-FND y la relación lignina/nitrógeno (tabla 2).
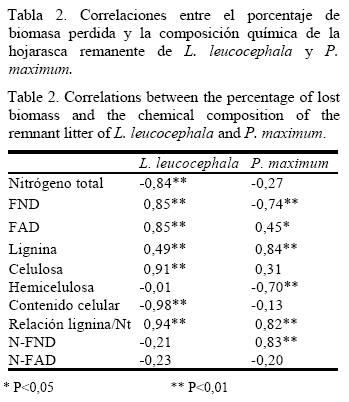
Al tomar en consideración las mayores correlaciones, se determinó la variación que experimentaron estos indicadores durante el proceso de descomposición de la hojarasca.
En la hojarasca de P. maximum (fig. 2) la concentración de lignina y del N-FND mostraron ajustes significativos al modelo lineal, lo que indica que a medida que ocurrió el proceso de descomposición aumentó la concentración de ambos indicadores. Por su parte, la relación lignina/Nt se incrementó también en el material remanente, con mayor ajuste al modelo cuadrático.
Es conocido que durante la descomposición, las concentraciones de lignina, tanino, celulosa, hemicelulosa, nitrógeno y carbono pueden aumentar (Edmon y Thomas, 1995; Badejo et al., 1998). Estos cambios se relacionan con la colonización y actividad de la flora y la fauna descomponedora. En este sentido, Hunter et al. (2003) y Barajas-Guzmán y Álvarez-Sánchez (2003) señalaron que la fauna del suelo y de la hojarasca son los factores biológicos reguladores de la descomposición que causan cambios químicos en los compuestos de esta; sin embargo, los estudios más recientes señalan que esta relación es empírica y no se ha encontrado explicación aún a este fenómeno (Berg y Laskowski, 2005).
En la hojarasca de la leucaena (fig. 3) el contenido celular y el nitrógeno mostraron una tendencia a disminuir; mientras que la concentración de la celulosa y la relación lignina/Nt presentaron un incremento lineal en la medida que transcurrió el proceso de descomposición. Este comportamiento puede atribuirse a lo apetecible que resultan los primeros constituyentes a la fauna descomponedora y, como resultado de ello, existe un aumento en la proporción de los más recalcitrantes.
CONCLUSIONES
De forma general, los resultados demostraron que en cada especie estudiada se presentó un patrón diferente de descomposición de la hojarasca. La tasa promedio de descomposición en la leucaena fue mayor que en la guinea. En ambas especies se encontró una rápida pérdida de peso durante los primeros 30 días; después de este primer mes el proceso fue más lento. Este comportamiento puede estar relacionado con la composición química de las diferentes especies de pastos, ya que el porcentaje de biomasa perdida de la hojarasca de L. leucocephala presentó una mayor correlación con las concentraciones del contenido celular, la relación lignina/nitrógeno, la celulosa y el Nt. En cuanto a la hojarasca de P. maximum se observó una dependencia significativa y negativa con las concentraciones de la FND y la hemicelulosa; mientras que se relacionó de forma positiva con las de N-FND y la relación lignina/nitrógeno.
REFERENCIAS BIBLIOGRÁFICAS
1. Academia de Ciencias de Cuba. 1989. Nuevo Atlas Nacional de Cuba. Instituto Cubano de Geodesia y Cartografía. La Habana, Cuba. p. 41
2. Aceñolaza, P.G. & Gallardo, J.F. 1995. Influencia de la edad del bosque en la evolución de la pérdida de peso seco en hojarasca de Alnus acuminata en alisedas de la provincia de Tucumán (Argentina). Investigación Agraria. Sistemas y Recursos Forestales. 4:177
3. Aerts, R. & De Caluwe, H. 1997. Nutritional and plant mediated controls on leaf litter decomposition of Carex species. Ecology.78:244
4. AOAC. 1995. Official methods of analysis. Ass. Off. Agric. Chem. Washington, D.C.
5. Badejo, M.A. et al. 1998. Abundance of springtails (Collembola) under four agroforestry trees species with contrasting litter quality. Biology and Fertility of Soil. 27:15
6. Barajas-Guzmán, G. & Álvarez-Sánchez, J. 2003. The relationships between litter fauna and rates of litter decomposition in a tropical rain forest. Applied Soil Ecology. 24:91
7. Bardgett, R.D. & Shine, A. 1999. Linkages between plant litter diversity, soil microbial biomass and ecosystem functioning temperate grasslands. Soil Biology & Biochemistry. 31:317
8. Berg, B. 2000. Litter decomposition and organic matter turnover in northern forest soils. Forest Ecology and Management. 13:133
9. Berg, B. & Laskowski, R. 2005. Litter decomposition: A guide to carbon and nutrient turnover. (Eds. B. Berg & R. Laskowski). Academic Press, New York. 448 p.
10. Caldentey, J. et al. 2001. Litter fluxes and decomposition in Nothofagus pumilio stands in the region of Magallanes, Chile. Forest Ecology and Management. 148:145
11. Crespo, G. et al. 2001. Tasas de acumulación, descomposición y NPK liberados por la hojarasca de leguminosas perennes. Rev. cubana Cienc. agríc. 35:39
12. Edmonds, R.L. & Thomas, T.B. 1995. Decomposition and nutrient release from green needles of western hemlock and Pacific silver fir in an old-growth temperate rain forests, Olympic National Park Washington. Can. J. For. Res. 25:1049
13. González, I.M. & Gallardo, J.F. 1995. El efecto de la hojarasca: una revisión. En: Anales de Edafología y Agrobiología. Centro de Edafología y Biología Aplicada. Salamanca, España. p. 86
14. Hernández, A. et al. 1999. Nueva versión de la clasificación genética de los suelos de Cuba. Ministerio de la Agricultura. La Habana, Cuba. p. 26
15. Herrera, R.S. 1981. Influencia del fertilizante nitrogenado y la edad del rebrote en la calidad del pasto Bermuda cruzada (Cynodon dactylon vc Coast cross 1). Tesis de Doctor en Ciencias. Instituto de Ciencia Animal. La Habana, Cuba. p. 145
16. Hunter, M.D. et al. 2003. Relative effects of macroinvertebrates and habitat on the chemistry of litter during decomposition. Pedobiología. 47:101
17. InfoStat. 2001. Software estadístico. Manual de usuario. Versión 1. Córdoba, Argentina.
18. Jamaludhee, V. & Kumar, M.B. 1999. Litter of multipurpose tree in Kerala, India: Variations in the amount, quality, decay rates and the release of nutrients. Forest Ecology and Management. 115:1
19. King, K.L. & Hutchinson, K.J. 1992. Microarthropods and nutrient transfer from pasture litter. Proceedings 6th Australian Society of Agronomy Conference, Armidale. p. 362
20. Liu, W. et al. 2000. Leaf litter decomposition of canopy trees, bamboo and moss in a montane moist evergreen broad-leaved forest on Ailao Mountain, Yunnan, south-west China. Ecol. Res. 15:435
21. Martín, A.E. 1995. Reciclado de bioelementos a través de la hojarasca en ecosistemas forestales de la Sierra de Gata. Sistema Central Español. Universidad de Salamanca. Facultad de Ciencias Químicas, España.
22. Mtambanengwe, F. & Kirchmann, H. 1995. Litter from a tropical savanna woodland (Miombo): Chemical composition and C and N mineralization. Soil Biol. Biochem. 27:1639
23. Saviozzi, A. et al. 1995. Decomposition of crop residues under laboratory conditions. Soil Use and Management. 11:193
24. Vanlauwe, B. et al. 1997. Decomposition of four Leucaena y Senna prunings in alley cropping systems under sub-humid tropical conditions. The process and its modifiers. Soil Biology and Biochemistry. 29:131
25. Van Soest, P.J. & Wine, R.H. 1968. Determination of lignin and cellulose in aciddetergent fibre with permanganate. Journal of the Association of Official Agricultural Chemists. 51:780
26. Vitousek, P.M. et al. 1994. Litter decomposition on the Manua Loa environmental matrix, Hawaii: patterns, mechanisms and models. Ecology. 75:418
27. Wieder, R. & Lang, G. 1982. A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology. 63:1636
Recibido el 4 de mayo del 2009
Aceptado el 10 de junio del 2009

{kind=link}
{kind=link}