










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.32 n.4 Matanzas oct.-dic. 2009
ARTÍCULO DE INVESTIGACIÓN
Efecto de la inclusión de Leucaena spp. en el comportamiento de la comunidad vegetal
Effect of the inclusion of Leucaena spp. on the performance of the plant community
Hilda B. Wencomo Estación Experimental de Pastos y Forrajes "Indio Hatuey". Central España Republicana. CP 44280. Matanzas, Cuba
E-mail: hilda.wencomo@indio.atenas.inf.cu
RESUMEN
Se realizó un estudio durante dos años en un área de 2 058 m2, en un suelo Ferralítico Rojo de la EEPF «Indio Hatuey», cuyo objetivo fue evaluar el comportamiento de la comunidad vegetal con la inclusión de árboles de leucaena. Se utilizaron parcelas sencillas de 2 x 1 m. Los indicadores estudiados fueron: disponibilidad de materia seca, composición química, composición florística del pastizal, población, altura y diámetro de la leucaena. En la composición florística del pastizal se pudo observar el predominio de P. maximum (P<0,001). Esta asociación permitió un aumento de 6% en la población del pasto mejorado; mientras que leucaena incrementó su grosor y altura. A su vez, la disponibilidad de MS total fue superior a las 8 t/ha/rotación durante el período de evaluación. Por su parte, las gramíneas presentaron niveles de proteína que oscilaron entre 9,7 y 11,7%, para la época poco lluviosa y la lluviosa, respectivamente; mientras que en leucaena fue superior al 20%. No se detectaron diferencias significativas en la composición química entre las épocas del año para ninguna de las especies. Se logró mantener la composición florística por encima del 80% al incrementarse el área cubierta por el pasto mejorado; la arbórea evolucionó de forma positiva en el tiempo, al aumentar el diámetro del tronco.
Palabras clave: Comunidades vegetales, Leucaena spp., Panicum maximum
ABSTRACT
A study was conducted for two years in an area of 2 058 m2, on a Ferralitic Red soil of the EEPF «Indio Hatuey», which objective was to evaluate the performance of the plant community with the inclusion of leucaena trees. Simple 2 x 1 m plots were used. The studied indicators were: dry matter availability, chemical composition, floristic composition of the pastureland, population, height and diameter of leucaena. In the floristic composition of the pastureland, the predominance of P. maximum (P<0,001) could be observed. This association allowed a 6% increase in the population of the improved pasture; while leucaena increased its diameter and height. In turn, total DM availability was higher than 8 t/ha/rotation during the evaluation period. On the other hand, the grasses showed protein levels that oscillated between 9,7 and 11,7%, for the dry and rainy season, respectively; while in leucaena it was higher than 20%. No significant differences were detected in the chemical composition between seasons for any of the species. The floristic composition could be kept over 80% increasing the area covered by the improved pasture; the tree evolved positively in time, increasing the stem diameter.
Key words: Plant communities, Leucaena spp., Panicum maximum
INTRODUCCIÓN
A nivel mundial se promueve actualmente el desarrollo sostenible como la meta hacia la que debe marchar la humanidad en su lucha por la supervivencia de la especie, en armonía con la naturaleza.
La escasez de alimentos durante la época poco lluviosa también ha traído como consecuencia la búsqueda de soluciones para cubrir el déficit de nutrientes que se produce durante ese período (Benavides, 2003). En este sentido, la presencia de árboles forrajeros de la familia de las leguminosas constituye una alternativa de explotación de los recursos naturales (Ibrahim y Mora, 2003).
Son incontables los aportes que hacen los árboles al ecosistema y sus aplicaciones como fuente alternativa para la alimentación animal. El uso de las leguminosas de hábito de crecimiento arbóreo, con alto potencial productivo y elevado valor nutritivo, se presenta como una solución económica viable y socialmente aceptable para incrementar la productividad animal en las regiones tropicales (Hernández-Daumás y Russell, 2001).
En condiciones de investigación, con el empleo de los árboles se han logrado rendimientos de MS similares a los obtenidos cuando se aplican niveles medios de fertilización; además se mantiene una alta persistencia de las leguminosas herbáceas y/o rastreras y de las gramíneas (Murgueitio, 2003).
Los estudios realizados en sistemas sostenibles en el trópico recomiendan el silvopastoreo como una alternativa para los productores que dispongan de pocos recursos. En este sistema se optimiza la producción de la especie leñosa y de las gramíneas; además, se crea una estructura vegetativa similar a la de las sabanas (Shaver, 2002). Las formas más estudiadas son: banco de proteína, asociación de árboles en toda el área de pastoreo y cercas vivas (Pezo e Ibrahim, 1999).
Entre las especies de la familia de las leguminosas de porte arbóreo más empleadas en estos sistemas se destaca Leucaena leucocephala. Esto se debe a su gran versatilidad, control de la erosión, reforestación, producción de madera y sus derivados, árbol de sombra, fertilizante orgánico y alimento para el ganado.
En Cuba se han realizado investigaciones para evaluar la influencia de las asociaciones de plantas de leucaena con gramíneas, en la respuesta productiva de la alimentación del ganado, sobre todo el bovino. El presente trabajo tuvo como objetivo evaluar el comportamiento de la comunidad vegetal con la inclusión de árboles de leucaena.
MATERIALES Y MÉTODOS
Localización. La investigación se realizó en un área de 2 058 m2, en la EEPF «Indio Hatuey», provincia de Matanzas, Cuba. Las coordenadas geográficas del lugar son 20º 50' de latitud norte y 79º 32' de longitud oeste, con una altitud de 19,9 msnm.
Características edafoclimáticas. El suelo se clasifica como Ferralítico Rojo lixiviado (Hernández et al., 2003), de topografía plana, con una pendiente de 1,0%. La temperatura media anual fue de 23,1ºC, con una media de 21 a 26,5ºC en invierno y en verano, respectivamente. La precipitación promedio anual es de 1 200 mm, con una humedad relativa entre 60-70% durante el día y de 80-90% en la noche (Hernández, 2000).
Manejo. En la etapa de evaluación el área experimental fue sometida, en condiciones de pastoreo simulado, a una carga instantánea de 51,9 UGM/ha, de la raza Cebú (machos en crecimiento), con un peso promedio de 240 kg. El tiempo de ocupación del área fue de tres días aproximadamente, con lo que se garantizaron períodos de reposo de dos y cuatro meses en las épocas lluviosa y poco lluviosa, respectivamente.
Diseño experimental. En el área se sembraron 34 accesiones de Leucaena spp. (tabla 1), en parcelas simples de 2,0 x 1,0 m distribuidas al azar. Las parcelas contaron con una densidad inicial de 10 497 plantas/ha.
Mediciones
Disponibilidad y calidad del pasto. Se empleó el método alternativo propuesto por Martínez et al. (1990), que consiste en la estimación de la disponibilidad de pasto utilizando su altura media; los muestreos (80 en cada parcela) se realizaron todos los meses. Se tomaron muestras del pasto (200 g) y se estimó su calidad, simulando con la mano la selección que hace el animal en pastoreo.
Disponibilidad de las plantas de leucaena. La disponibilidad se estimó en 10 de los árboles establecidos, simulando el ramoneo que realizan los animales según su altura de consumo (Lamela, 1998); se aplicó la técnica del ordeño de las partes más tiernas (hojas y tallos finos). Se tomaron muestras para determinar su composición bromatológica.
Altura de las plantas de leucaena. Se determinó la altura con una regla graduada, al inicio, a mediados y al final del experimento, en el 100% de las plantas.
Diámetro del tronco y número de ramas primarias de leucaena. Las variables dasométricas se determinaron al inicio de cada rotación en cada parcela, en 10 plantas seleccionadas. El diámetro del tronco se midió a 1,30 m a la altura del pecho, con un pie de rey, y el número de ramas se contó en las plantas seleccionadas. Posteriormente se efectuó la poda de uniformidad (en noviembre), a 1 m sobre el nivel del suelo, según lo recomendado por Francisco et al. (1998), ya que las plantas sobrepasaban los 2 m de altura.
Análisis de laboratorio. Después de efectuar cada muestreo se enviaron muestras representativas de pasto al laboratorio para determinar la composición bromatológica del alimento. Los indicadores medidos fueron: porcentaje de materia seca, de proteína bruta, de fibra bruta, de calcio y de fósforo. Se emplearon las técnicas descritas por la AOAC (1990). El contenido de P se determinó mediante el método de Amaral (1972).
Composición florística del pastizal. Se estimó por el método de los pasos, descrito por Anon (1980). Cada dos pasos el observador clasificó la especie de pasto que coincidió con la punta de su zapato. Esta medición se realizó al inicio y al final de cada época del año.
Procesamiento de los resultados. Se sometieron a un ANOVA según modelo lineal de clasificación simple, considerando el efecto de la época y de las accesiones, y las medias fueron comparadas mediante la dócima de Duncan para un 5% de significación, después de verificarse que cumplían con el ajuste de distribución normal y de homogeneidad de varianza, a través del programa estadístico SPSS® versión 11,5 para Microsoft® Windows® (Visuata, 1998).
RESULTADOS Y DISCUSIÓN
La composición florística del pastizal se presenta en la figura 1. La especie predominante fue la guinea (Panicum maximum), que mostró una mejor adaptación; mientras que los pastos naturales y las leguminosas rastreras mantuvieron estabilidad; las leguminosas estuvieron representadas por centrosema (Centrosema pubescens) y macroptilium (Macroptilium atropurpureum) y aparecieron de forma espontánea en el sistema, lo cual evidenció la persistencia del pastizal y la adaptación de estas especies a las condiciones de explotación del área.
La guinea tuvo un decrecimiento en el segundo año, aspecto que según lo informado por Hernández (2000), pudo estar influido por la selección que realizan los animales al pastar, ya que se conoce de la preferencia de estos por los pastos que tienen una alta proporción de hoja-tallo, como P. maximum; esta planta está considerada entre los pastos tropicales que poseen una estructura más ventajosa, razón por la que el ganado manifiesta su capacidad selectiva (Hernández et al., 1998).
Por su parte, los pastos naturales durante las dos épocas del año se mantuvieron con una población entre el 20 y 27%, sin diferencia significativa durante el experimento, lo que es un indicador de la estabilidad del sistema en el período evaluado. Similar comportamiento fue reportado por Sánchez et al. (2002) al estudiar el efecto de una asociación de Leucaena con gramíneas mejoradas en la producción de leche.
Por su parte, las malezas tuvieron un valor inicial de 7,5% y su tendencia fue a disminuir. Es necesario destacar que en el área se chapeó en cuatro ocasiones, con el objetivo de controlarlas, lo cual influyó en los resultados.
Las leguminosas rastreras pudieron persistir debido a la inclusión de los árboles, los cuales sirvieron como tutores y evitaron las defoliaciones excesivas provocadas por el diente del animal durante el pastoreo (Simón, 2005). A pesar de ello hubo un decrecimiento de su población, lo que pudo deberse al pisoteo o a la selección que realizan los animales.
La composición química de las plantas de leucaena y de las gramíneas se muestra en la figura 2; no se hallaron diferencias significativas en el contenido de proteína cruda, tanto para el período lluvioso como para el poco lluvioso en ambas especies, ni para el contenido de calcio y de fósforo, cuyos valores se encontraron entre 0,8 y 1,3%, y 0,13 y 0,18%, respectivamente.
Las plantas de leucaena presentaron un contenido de proteína bruta alto, con valores superiores al 20%, que coinciden con los informados con anterioridad por García et al. (2007). Resultados similares también han sido reportados para estas y otras accesiones de arbóreas por Galindo et al. (2005). El contenido de PB permite aseverar que las especies y accesiones de este género presentan una elevada homogeneidad en este indicador.
González y Cáceres (2002), al estudiar el valor nutritivo de diferentes árboles, arbustos y otras plantas para la alimentación de los rumiantes, indicaron que a diferencia de lo que de forma frecuente ocurre con otras forrajeras (como las gramíneas y las leguminosas herbáceas) en similares condiciones agrotécnicas, ninguno de los indicadores sufrió variación en sus valores por el efecto de la época; ello se debió a que las raíces de las accesiones de leucaena penetran en los estratos profundos del suelo y propician la extracción del agua, lo que les permite manifestar un comportamiento más estable a través de todo el año en cuanto a la composición química de su biomasa.
A su vez los pastos mostraron, en sentido general, valores adecuados de PB, similares a los informados por Lamela et al. (1999) cuando se asociaron a la leucaena y equivalentes a los encontrados al emplear niveles de fertilización entre 150 y 300 kg de N/ha/año.
La mejoría en la calidad nutricional de los pastos se debió a la presencia de la arbórea en el pastizal, la cual tiene la capacidad de fijar el nitrógeno atmosférico al suelo a través de la simbiosis con los rizobios, que a su vez es aprovechado por los pastos que se hallan en el sistema; además, la leucaena extrae nutrientes y agua de las capas más profundas, debido a su profuso sistema radical.
La concentración de los componentes estructurales (fibra bruta) para las especies de Leucaena estudiadas estuvo entre 18 y 25%. Con relación a esto, Clavero y Razz (1999) encontraron incrementos del contenido de fibra neutro detergente (FND) y disminución de la proteína bruta a medida que transcurría el ciclo biológico de la planta, debido al aumento de los elementos estructurales y la reducción del número de hojas jóvenes.
En el caso de las gramíneas los valores se pueden considerar aceptables; la fibra bruta es un indicador de la calidad del pasto porque cuando presenta un incremento (aunque sea ligero) se debe a un aumento de las partes menos digeribles de las plantas. Estos valores coinciden con los informados por Sánchez (2007) al estudiar sistemas asociados de guinea y leucaena.
Los contenidos de calcio y fósforo en las accesiones de Leucaena se hallaron dentro del rango de valores para este género (Shelton y Brewbaker, 1994; Anon, 2000); un comportamiento similar se presentó en las gramíneas. De ahí la necesidad de una suplementación fosfórica a los animales que pastoreaban en el sistema, si se tiene en consideración su importancia en la reproducción de las vacas y en el metabolismo. Es significativo señalar que la carencia de P en las dietas constituye un problema para la nutrición adecuada, debido al bajo contenido que presentan los pastos tropicales, lo cual se acentúa en los suelos deficientes en este mineral (Rolo, 1999).
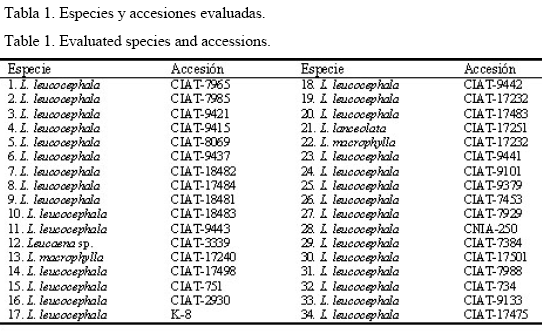
En la figura 3 se muestra el comportamiento de la disponibilidad y la altura del pasto durante la evaluación, en las cuales se encontraron variaciones altamente significativas (P<0,001).
De forma general existió una alta disponibilidad del pasto durante toda la etapa de evaluación; sin embargo, se apreció un efecto de la época del año en este indicador. Similar comportamiento se informó en Cuba por Lamela et al. (1999) para diferentes sistemas silvopastoriles, en los que se obtuvieron las mayores disponibilidades del pasto en el período lluvioso. En esta época del año las precipitaciones son mayores, al igual que la temperatura y la radiación solar, lo cual favorece el crecimiento de los pastos.
Los mayores valores se observaron en la lluvia de 1997 y en la seca de 1998 (11,9 y 9,5 t MS/ha, respectivamente). El rendimiento en la lluvia de 1997 pudo deberse a que existía un gran volumen de biomasa, que se había acumulado durante el tiempo en que el sistema permaneció en reposo mientras se establecía, y la guinea alcanzó más de un metro de altura; además en este período las plantas muestran su mayor vigor juvenil, ya que se produce una mayor acumulación de reservas en los pastos a causa del reposo que transcurre desde la siembra hasta que comienzan las defoliaciones o los cortes, lo que influye de forma positiva en los rendimientos (Gerardo y Thompson, 1985).
Las condiciones ambientales que prevalecieron en la lluvia de 1998 pudieron influir en el aumento de la disponibilidad de la guinea en esta etapa y propiciar el buen desarrollo de los rebrotes, lo que resultó más efectivo en unas accesiones que en otras. Es válido mencionar que los volúmenes de biomasa pudieron deberse, a su vez, a los tiempos de reposo, que fueron mayores que los utilizados comúnmente en los sistemas silvopastoriles (40 días en el período lluvioso y 120 días en el poco lluvioso).
La distribución de las lluvias y las variaciones de la temperatura durante este período influyeron en el crecimiento de esta especie; en tal sentido, Fernández et al. (2002) plantean que este efecto provoca un desbalance estacional de los rendimientos y limitaciones en la disponibilidad de forraje durante la época poco lluviosa.
Posteriormente la disponibilidad de pasto declinó en los muestreos de la seca de 1998 y de la lluvia de ese mismo año (4,5 y 4,7 t de MS/ha, respectivamente). Al final de la evaluación (lluvia-98) la producción de la guinea se incrementó a 9,5 t de MS/ha y alcanzó una altura de 115 cm. Según Padilla et al. (2000) estas variaciones en la disponibilidad de la guinea asociada a leucaena pudieran estar relacionadas con las condiciones ambientales y de manejo en que coincidieron los muestreos; todos estos factores en su conjunto influyen notablemente en el establecimiento, el nivel de producción y la vida útil de los pastos.
Los rendimientos de P. maximum corroboran su versatilidad, tanto en condiciones de arbolado como de sol, por ser una especie heliófila facultativa, con un alto nivel de plasticidad fenotípica que le permite explotar de forma óptima los recursos en diferentes ambientes (Pentón, 2000).
Por su parte, las plantas de leucaena presentaron una mayor disponibilidad de materia seca en la seca de 1998 y en la lluvia de ese año, debido a la altura alcanzada en esta etapa (superior a 300 cm), por lo que fue necesario una poda estratégica para poner el forraje al alcance de los animales (Simón y Francisco, 2003), la cual coincidió con el período de menores precipitaciones, temperatura y humedad.
La disponibilidad de biomasa total de leucaena presentó una disminución altamente significativa (P<0,001) durante la evaluación (fig. 4). Los mayores aportes (1 444,70 y 1 276,76 kg de MS/ha) se observaron en la seca de 1997 y en la lluvia de 1998; en los muestreos posteriores se obtuvo un rendimiento de 977 y 870 kg de MS/ha, respectivamente.
Esta dinámica manifiesta el comportamiento que presentan las arbóreas en asociación sometidas a defoliaciones y pastoreos continuos, durante el primer año de evaluación (Papanastasi et al., 1998), y ha sido observado también en otras especies como Gliricidia sepium (Razz, 1991) y Albizia lebbeck (Francisco, 2003).
En el caso de la biomasa comestible y sus componentes (hojas y tallos tiernos) se manifestó un patrón similar, ya que hubo un decrecimiento altamente significativo (P<0,001) con los muestreos sucesivos. La producción de material comestible al inicio de la evaluación (lluvia del 2000) fue de 954 kg de MS/ha, en la seca de 1998 se incrementó a 1 220 kg de MS/ha y en la lluvia del mismo año mostró una marcada disminución (694 kg de MS/ha).
Ello pudiera estar determinado por su capacidad para la movilización de las reservas, la que facilitó la emisión de rebrotes, como ha ocurrido en la especie G. sepium (García et al., 2001). En este sentido, Berninger et al. (2000) señalaron que las frecuencias de corte imponen cambios en la dinámica de todas las partes de la biomasa y, a su vez, en la movilización de las reservas de los carbohidratos, tan esenciales y determinantes para proporcionar el crecimiento de los rebrotes.
La menor disponibilidad de biomasa comestible a partir del muestreo de la lluvia de 1998 (junio), la cual se mantuvo hasta septiembre, pudo deberse al estrés como producto del pastoreo y la poda; ello pudo disminuir sus reservas y, desde el punto de vista funcional, la planta tuvo una mayor necesidad de hacer uso de los carbohidratos de reserva de la raíz para sustentar el rebrote.
La producción de tallos leñosos mostró una disminución altamente significativa (P<0,001) con los sucesivos muestreos, que fue más marcada en la seca de 1997 (75 kg de MS/ha) y en la lluvia de 1998 (96 kg de MS/ha); esto se relacionó con la mayor fortaleza del rebrote y la cantidad de sustancias de reserva almacenadas en el tallo y en la raíz, que aumentaron de forma progresiva en el tiempo de reposo al que se sometieron las plantas, y coincide con lo referido por García et al. (2001).
Este resultado pudo estar influido también por el efecto acumulado del manejo al que estuvo sometido el pastizal, el cual permitió la rehabilitación, el rejuvenecimiento y el desarrollo posterior de este pasto (Hernández et al., 1999).
Es válido señalar que a pesar de que en la época lluviosa las precipitaciones son mayores, al igual que la temperatura y la radiación solar (lo cual favorece el crecimiento de las especies), en esta evaluación no se apreció efecto de la época del año en la disponibilidad de materia seca de las accesiones de Leucaena. Esto puede atribuirse a que las raíces de las plantas de este género penetran en los estratos profundos del suelo y propician la extracción del agua; además presentan un sendero fotosintético C3 por lo que necesitan menos intensidad luminosa que las gramíneas, que son C4 (Pérez-Infante, 1977; Simón, 1998). Ello beneficia tanto al pasto acompañante como a los microorganismos que habitan en el suelo y es de vital importancia en el sistema, ya que permite su conservación y el incremento de la biodiversidad.
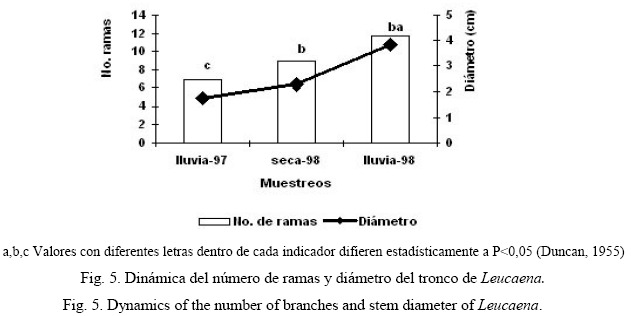
Se pudo observar, además, que durante el período de evaluación hubo un incremento significativo del diámetro del tronco de la arbórea, así como del número de ramas (fig. 5). Al final de la evaluación la arbórea presentó un promedio de 14 ramas y un grosor del fuste de 2,45 cm.
La dinámica significativamente ascendente del diámetro del tronco y el número de ramas evidenció la elevada recuperación que presenta esta planta después de la poda. Según Dávila y Urbano (1996), el incremento de los indicadores dasométricos en las plantas arbóreas denota el desarrollo progresivo de la plantación. El número de ramas no presentó diferencias significativas, lo que pudo estar motivado por las características particulares de su disposición.
En cuanto a la supervivencia de las plantas no hubo variaciones. Según Simón et al. (1994), la estabilidad de la población de los árboles sometidos a los efectos de la poda y el ramoneo determina sus posibilidades para persistir en el sistema, por lo que la resistencia al ramoneo y la capacidad de recuperación después de la poda forman parte de los atributos que debe reunir una arbórea para su inclusión en este tipo de sistema. En este sentido, Simón y Francisco (2003) señalaron una serie de características agronómicas y zootécnicas que pueden determinar en menor o mayor medida la persistencia del árbol dentro del pastizal, y destacan a L. leucocephala entre las especies arbóreas que mejor se adaptan a las condiciones de pastoreo.
De acuerdo con los resultados se concluye que la asociación de pastos con Leucaena presentó una disponibilidad media de MS, lo cual permitió garantizar una oferta de alimentos con un contenido aceptable de proteína bruta (11,7 y 9,7% para las gramíneas y 30 y 27% para la Leucaena). Los pastos se beneficiaron con la presencia de la arbórea, lo cual se observó en la mejoría de su composición química, además de incrementar su población en un 6%; mientras que la arbórea mantuvo una alta densidad en el pastizal.
REFERENCIAS BIBLIOGRÁFICAS
1. Amaral, A. 1972. Técnicas analíticas para evaluar macronutrientes en ceniza de caña de azúcar. Laboratorio de caña. Escuela de Química, Universidad de La Habana, Cuba
2. Anon. 1980. Muestreo de pastos. Taller del IV Seminario Científico de la EEPF «Indio Hatuey». Matanzas, Cuba
3. Anon. 2000. Tablas de valor nutritivo y requerimientos para el ganado bovino. Pastos y Forrajes. 23:105
4. AOAC. 1990. Official methods of analysis. 17th ed. Association of Official Agricultural Chemists, Washington. 1298 p.
5. Benavides, J. 2003. Árboles y arbustos forrajeros: Una alternativa para la sostenibilidad en la ganadería. Memorias. Taller Internacional «Ganadería, desarrollo sostenible y medio ambiente». La Habana, Cuba. 157 p.
6. Berninger, F. et al. 2000. Modelling of reserve carbohydrate dynamics, regrowth and nodulation in a N2-fixing tree managed by periodic prunings. Plant, Cell and Enviroment. 23:1025
7. Clavero, T. & Razz, R. 1999. Valor nutritivo de la Gliricidia sepium en condiciones de bosque seco tropical. Rev. cubana Cienc. agríc. 33:97
8. Dávila, C. & Urbano, D. 1996. Leguminosas arbóreas en la zona sur del Lago de Maracaibo. En: Leguminosas forrajeras arbóreas en la agricultura tropical. (Ed. T. Clavero). Centro de Transferencia de Tecnología en Pastos y Forrajes. Universidad del Zulia, Venezuela. p. 101
9. Fernández, J.L. et al. 2002. Efecto de la edad de rebrote en el rendimiento de Brachiaria purpurascens cv. Aguada en el Valle del Cauto en Cuba. Rev. cubana. Cienc. agríc. 34:267
10. Francisco, Geraldine. 2003. Manejo estratégico de las defoliaciones en especies arbóreas. Pastos y Forrajes. 26:185
11. Francisco, Geraldine et al. 1998. Efecto de tres alturas de corte en el rendimiento de biomasa de Leucaena leucocephala cv. CNIA-250. Pastos y Forrajes. 21:337
12. Galindo, J. et al. 2005. Impacto de los árboles, los arbustos y otras leguminosas en la ecología ruminal de animales que consumen dietas fibrosas. Pastos y Forrajes. 28:59
13. García, D.E. et al. 2007. Variabilidad fitoquímica y repercusión antinutricional potencial en especies del género Albizia. Pastos y Forrajes. 29:153
14. García, H. et al. 2001. Dynamics of non-structural carbohydrates and biomass yield in a fodder legume tree under different harvest intensities. Tree Physiol. 21:523
15. Gerardo, J. & Thompsom, M. 1985. Evaluación zonal de pastos tropicales bajo condiciones de pastoreo. XII. Empresa Pecuaria La Sierrita. Pastos y Forrajes. 8:337
16. González, E. & Cáceres, O. 2002. Valor nutritivo de árboles, arbustos y otras plantas forrajeras para los rumiantes. Pastos y Forrajes. 25:15
17. Hernández, A. et al. 2003. Nuevos aportes a la clasificación genética de suelos en el ámbito nacional e internacional. Instituto de Suelos. Ministerio de la Agricultura. AGRINFOR. La Habana, Cuba. 145 p.
18. Hernández, D. et al. 1998. Explotación de un sistema silvopastoril multiasociado para la producción de leche. Memorias. III Taller Internacional Silvopastoril «Los Árboles y arbustos en la ganadería». EEPF «Indio Hatuey». Matanzas, Cuba. p. 214
19. Hernández, D. et al. 1999. Establecimiento de un sistema silvopastoril multiasociado. Pastos y Forrajes. 22:123
20. Hernández-Daumás, S. & Russell, G. 2001. The tree-grass-soil interaction in silvopastoral systems. In: International Symposium on Silvopastoral Systems. Second Congress of Livestock Production in Latin America. San José, Costa Rica. p. 140
21. Hernández, I. 2000. Utilización de las leguminosas arbóreas L. leucocephala, A. lebbeck y B. purpurea en sistemas silvopastoriles. Tesis presentada en opción al grado de Dr. en Ciencias Agrícolas. Instituto de Ciencia Animal. Estación Experimental de Pastos y Forrajes «Indio Hatuey», Matanzas, Cuba. 118 p.
22. Ibrahim, M. & Mora, J. 2003. Criterios y herramientas para la promoción de una ganadería eco-amigable en el trópico americano. Memorias. Taller Internacional «Ganadería, desarrollo sostenible y medio ambiente». La Habana, Cuba. p. 23
23. Lamela, L. 1998. Métodos de muestreo y mediciones en sistemas silvopastoriles. En: Compendio de Conferencias para el Diplomado en Silvopastoreo. EEPF «Indio Hatuey», Matanzas, Cuba.
24. Lamela, L. et al. 1999. Producción de leche en un sistema con banco de proteína. Pastos y Forrajes. 22:239
25. Martínez, J. et al. 1990. Un método ágil para estimar la disponibilidad de pasto en una vaquería comercial. Pastos y Forrajes. 13:101
26. Murgueitio, E. 2003. Investigación participativa en sistemas silvopastoriles integrados: La experiencia de CIPAV en Colombia. Memorias. Taller Internacional «Ganadería, desarrollo sostenible y medio ambiente». La Habana, Cuba. p. 207
27. Padilla, C. et al. 2000. Efecto del intercalamiento de Vigna unguiculata (INIFAT 93) y Zea maiz (Jívara) en Leucaena leucocephala cv. Perú y Panicum maximum cv. Likoni. Memorias IV Taller Internacional Silvopastoril «Los árboles y arbustos en la ganadería tropical». EEPF «Indio Hatuey». Matanzas, Cuba. p. 33
28. Papanastasi, V.P. et al. 1998. Effects of age and frequency of cutting on productivity of mediterranean deciduous fodder tree and shrubs plantation. Forest Ecology and Management. 110:283
29. Pentón, G. 2000. Nota técnica: Tolerancia del Panicum maximum cv. Likoni a la sombra en condiciones controladas. Pastos y Forrajes. 23:79
30. Pérez-Infante, F. 1977. Posibilidades de los pastos en el trópico. Rev. cubana de Cienc. agríc. 11:119
31. Pezo, D. & Ibrahim, M. 1999. Sistemas silvopastoriles. Colección de Módulos Agroforestales No. 2. CATIE. Turrialba, Costa Rica. p. 15
32. Razz, R. 1991. Mata Ratón (Gliricidia sepium). Cuadernos técnicos. Facultad de Agronomía. Universidad del Zulia, Venezuela. 5 p.
33. Rolo, R. 1999. Relación nutrición-fertilidad en la hembra bovina. Memorias. II Congreso Nacional de Medicina Veterinaria y Zootecnia. Colegio de Médicos Veterinarios de Honduras, Honduras. s/p
34. Sánchez, Tania. 2007. Evaluación productiva de una asociación de gramíneas mejoradas y Leucaena leucocephala cv. Cunningham con vacas Mambí de Cuba en condiciones comerciales. Tesis presentada en opción al Grado Científico de Doctor en Ciencias Veterinarias. Universidad de Camagüey, Cuba.103 p.
35. Sánchez, Tania et al. 2002. Efecto de una asociación de Leucaena con gramíneas mejoradas en la producción de leche. Pastos y Forrajes. 26:137
36. Shaver, P. 2002. Fire and silvopasture. USDA National Agroforestry Center. Inside agroforestry. Fall, p. 1
37. Shelton, H.M. & Brewbaker, J.L. 1994. Leucaena leucocephala-the most widely used forage tree legume. (Eds. R. C. Gutteridge and H. M Shelton). CAB International, UK. p 15
38. Simón, L. 1998. Del monocultivo de pastos al silvopastoreo: La experiencia de la EEPF «Indio Hatuey». En: Los árboles y arbustos en la ganadería. Tomo 1. Silvopastoreo. (Ed. L. Simón). EEPF «Indio Hatuey». Matanzas, Cuba. p. 9
39. Simón, L. 2005. El Silvopastoreo: Un nuevo concepto de pastizal. Editorial Universitaria, Guatemala-EEPF «Indio Hatuey», Matanzas, Cuba. 214 p.
40. Simón, L. et al. 1994. Efecto del pastoreo de Albizia lebbeck (Algarrobo de olor) en el comportamiento de hembras bovinas en crecimiento. Resúmenes. Taller Internacional «Sistemas silvopastoriles en la producción ganadera». EEPF «Indio Hatuey». Matanzas, Cuba. p. 38
41. Simón, L. & Francisco, G. 2003. Atributos deseables de los árboles forrajeros: Leucaena leucocephala, Albizia lebbeck y Gliricidia sepium. Revista Cubana de Producción Animal. 2:11
42. Visuata, B. 1998. Análisis estadístico con SPSS para Windows. Vol. II. Estadística multivariante. (Ed. C. Fernández). Madrid, España. p. 24
Recibido el 16 de marzo del 2009
Aceptado el 27 de agosto del 2009

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}