










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.33 n.1 Matanzas ene.-mar. 2010
ARTÍCULO DE INVESTIGACIÓN
Descomposición de la hojarasca en un sistema silvopastoril de Panicum maximum y Leucaena leucocephala (Lam) de Wit cv. Cunningham. III. Influencia de la densidad y diversidad de la macrofauna asociada
Litter decomposition in a silvopastoral system of Panicum maximum and Leucaena leucocephala (Lam) de Wit cv. Cunningham. III. Influence of density and diversity of the associated macrofauna
Saray Sánchez1, G.J. Crespo2 y Marta Hernández
1Estación Experimental de Pastos y Forrajes «Indio Hatuey» . Central España Republicana, CP 44280, Matanzas, Cuba
E-mail: saray.sanchez@indio.atenas.inf.cu
2Instituto de Ciencia Animal, La Habana, Cuba
RESUMEN
Con el objetivo de determinar la descomposición de la hojarasca en un sistema silvopastoril de Panicum maximum y Leucaena leucocephala y su relación con la densidad y la diversidad de la macrofauna asociada, se realizó este experimento en la EEPF «Indio Hatuey». Esta se determinó como la pérdida de biomasa a través del tiempo, con relación al peso inicial. Para el estudio de la dinámica de la descomposición se utilizó el método de bolsas de hojarasca (litter bags). Se escogieron al azar cuatro bolsas a los 30, 60, 90, 120, 150, 180 y 210 días de situadas en el pastizal. En cada fecha de recolección, a la hojarasca remanente de cada bolsa se le determinó la población de macrofauna (organismos con diámetro mayor que 2 mm) mediante la separación manual, y se calculó el valor promedio de la densidad (individuos/m2), así como la abundancia proporcional (%) para cada taxón. Se utilizó el análisis de correlación y regresión para conocer la interrelación entre las variables y los modelos de mejor ajuste. De acuerdo con los resultados se concluye que las condiciones de humedad y temperatura que genera el árbol en este sistema, así como la calidad de su hojarasca, posibilitan la presencia de una diversa y estable fauna asociada a las bolsas, la cual influyó en el proceso de descomposición.
Palabras clave: Biodiversidad, hojarasca, Leucaena leucocephala, Panicum maximum
ABSTRACT
The trial was conducted at the EEPF «Indio Hatuey» in order to determine the litter decomposition in a silvopastoral system of Panicum maximum and Leucaena leucocephala and its relation to the density and diversity of the associated macrofauna. It was determined as biomass loss through time, with regards to initial weight. For the study of the decomposition dynamics the litter bag method was used. Four bags were randomly chosen 30, 60, 90, 120, 150, 180 and 210 days after being placed in the pastureland. In each collection date, to the remnant litter of each bag, the macrofauna (organisms with diameter higher than 2 mm) population was determined by means of manual separation, and the average value of density (individuals/m2), as well as the proportional abundance (%) for each taxon, was calculated. The correlation and regression analysis was used to learn the interrelation among variables and the best adjustment models. According to the results, it is concluded that the humidity and temperature conditions generated by the tree in this system, as well as the quality of its litter, allow the presence of a diverse and stable fauna associated to the bags, which influenced the decomposition process.
Key words: Biodiversity, Leucaena leucocephala, litter, Panicum maximum
INTRODUCCIÓN
En la región tropical, los macroinvertebrados del suelo desempeñan un papel clave en el funcionamiento del ecosistema (De Aquino et al., 2008), debido a que contribuyen con diferentes servicios al ecosistema, mediante su acción sobre los procesos del suelo.
En este sentido, el proceso de descomposición de la hojarasca es uno de los más importantes, pues a través de él se libera materia orgánica y nutrientes al suelo. Según Lavelle (2000) entre los factores que regulan este proceso se encuentran: el clima (especialmente la temperatura y la humedad), la composición química de la hojarasca y los organismos del suelo. Estos últimos contribuyen a la descomposición, al fragmentar la hojarasca e ingerir materia orgánica (Brusaard et al., 1997).
Por lo antes expuesto, el presente trabajo tuvo como objetivo determinar la descomposición de la hojarasca de las principales especies presentes en un sistema silvopastoril (Panicum maximum y Leucaena leucocephala (Lam) de Wit cv. Cunningham) y su relación con la densidad y la diversidad de la macrofauna asociada.
MATERIALES Y MÉTODOS
Descripción del área de investigación
La investigación se realizó en la Estación Experimental de Pastos y Forrajes «Indio Hatuey», situada entre los 22° 48' y 7" de latitud Norte y los 81° y 2' de longitud Oeste, a 19,01 msnm, en el municipio de Perico, provincia de Matanzas, Cuba (Academia de Ciencias de Cuba,1989).
El suelo donde se llevó a cabo la fase experimental se clasifica como Ferralítico Rojo lixiviado (Hernández et al., 1999).
Caracterización del sistema. Al comenzar la evaluación en este pastizal (1,3 ha) P. maximum representaba el 79,7% de la composición botánica, seguido por 8,9% de pastos naturales (Sporobolus indicus y Paspalum notatum), 7,6% de Cynodon nlemfuensis y 3,4% de leguminosas herbáceas. La población de L. leucocephala era de 595 plantas ha-1.
Procedimiento experimental. La investigación se inició en febrero del 2004 y se extendió hasta septiembre de ese año. La descomposición de la hojarasca se determinó como la pérdida de biomasa a través del tiempo, con relación al peso inicial (Liu et al., 2000).
Para el estudio de la dinámica de la descomposición se utilizó el método de bolsas de hojarasca (litter bags) de Caldentey et al. (2001). Las bolsas medían 10 x 10 x 10 cm, con poros de 1 cm de diámetro, lo cual permite el acceso de un amplio rango de especies de la biota edáfica hacia el interior. Se distribuyeron al azar 40 bolsas que contenían hojarasca de P. maximum y 40 de L. leucocephala. En cada bolsa se colocaron 20 g de hojarasca (base seca) previamente colectada. Las bolsas se enumeraron y se situaron de manera que toda su superficie estuviera en contacto con el horizonte orgánico, y se fijaron al suelo mediante estacas metálicas.
Se escogieron al azar cuatro bolsas a los 30, 60, 90, 120, 150, 180 y 210 días de situadas en el pastizal. En cada fecha de recolección, a la hojarasca remanente de cada bolsa se le determinó la población de macrofauna (organismos con diámetro mayor que 2 mm) mediante la separación manual, según la metodología del Programa de Investigación Internacional «Biología y Fertilidad del Suelo Tropical» (TSBF), propuesta por Anderson e Ingram (1993) y se calculó el valor promedio de la densidad (individuos), así como la abundancia proporcional (%) para cada taxón. La densidad se determinó a partir del número de individuos y la abundancia relativa mediante la relación entre la cantidad de individuos que pertenecen a un grupo taxonómico y el total de individuos de todos los grupos taxonómicos.
Durante el período de estudio se registró diariamente el comportamiento de la temperatura mínima, máxima y media, la humedad relativa, la evaporación y las precipitaciones, en la estación meteorológica situada a 1 km del área experimental.
Análisis matemático. Se realizó un análisis de varianza según el modelo lineal de clasificación simple y en los casos necesarios se aplicó la dócima de Duncan (1955).
Se comprobaron los supuestos de normalidad de los errores por la prueba modificada de Shapiro Wilk (Royston, 1982), así como la homogeneidad de la varianza según la prueba de Bartlett (1937); en los casos en que no se cumplió, se transformó la variable según Öx.
Se utilizó el análisis de correlación y regresión lineal y múltiple para conocer la interrelación entre las variables y los modelos de mejor ajuste. Se consideró como variable independiente la frecuencia acumulada de la macrofauna, y como variable dependiente el peso de la hojarasca residual.
Para el procesamiento de la información se utilizó el software estadístico INFOSTAT (2001), versión 1.
RESULTADOS Y DISCUSIÓN
La dinámica de descomposición de la hojarasca de la leucaena y de la guinea en el sistema silvopastoril se muestra en la figura 1. La cantidad de material descompuesto fue mayor y el proceso fue más rápido en L. leucocephala, con diferencias altamente significativas entre los días de descomposición. A los 210 días quedó sin descomponer solo el 3,1% de la hojarasca de leucaena; sin embargo, en similar tiempo la hojarasca de la guinea aún representaba el 28,2% del peso inicial.

Existen numerosos factores bióticos y abióticos que influyen en la descomposición de la hojarasca; sin embargo, la macrofauna edáfica desempeña un papel muy importante en los procesos biogeoquímicos de los nutrientes, al realizar un conjunto de funciones esenciales que incluyen: la descomposición, el reciclaje de nutrientes, la síntesis y la mineralización de la materia orgánica, entre otros.
En este sentido, la composición taxonómica de la macrofauna asociada al proceso de descomposición de la hojarasca en este sistema silvopastoril, estuvo constituida por dos phylum, cinco clases, nueve órdenes, nueve géneros y nueve especies (tabla 1). En el phyllum Arthropoda tres clases fueron las más representativas, dentro de las cuales Insecta fue la más destacada. La comunidad estuvo constituida por el 56% de organismos detritívoros, 35% de herbívoros y 9% de depredadores.

El efecto de los invertebrados edáficos en la descomposición de la materia orgánica es esencial, ya que mediante su alimentación hacen el material más asequible a la acción de los microorganismos descomponedores, además de contribuir a la diseminación de hongos y bacterias y al transporte vertical de la materia orgánica desde la superficie hacia las capas más profundas del suelo, lo cual aumenta la velocidad de descomposición (Prieto y Rodríguez, 2001; Cotrufo et al., 2005; Isaac y Nair, 2005).
La selección de alimento depende mucho de la categoría ecológica del invertebrado. Los invertebrados epígeos, que viven y se alimentan de la hojarasca superficial (Cabrera, 2003), producen in situ modificaciones importantes de la hojarasca y de la madera en descomposición. Los artrópodos epígeos poseen una importancia adicional, ya que participan en infinidad de procesos que ocurren en el suelo, como la reducción de los fragmentos vegetales y el reciclado de nutrientes (Rivera y Carrasco, 1991; Torres et al., 2005).
El comportamiento de la densidad de invertebrados durante el proceso de descomposición de la hojarasca se muestra en la tabla 2. La mayor cantidad de individuos en la hojarasca de la guinea se alcanzó entre los 120-180 días y en la leucaena entre los 90-120 días.

Las condiciones de humedad y temperatura que genera la presencia del árbol en este sistema parecen haber incidido en este comportamiento, pues la hojarasca de ambas especies se mantuvo colonizada desde las primeras etapas y no se incrementó con los factores climáticos. La mayor colonización en la leucaena pudiera estar relacionada con lo apetecible que resulta la hojarasca de esta leguminosa para la fauna del suelo, debido fundamentalmente a su alto contenido de nitrógeno (Mwiinga et al., 1994).
La presencia de los grupos de la macrofauna asociados a la descomposición de la hojarasca de P. maximum en el sistema, se muestra en la figura 2. Se observaron diversos órdenes durante todo el proceso y un incremento a partir de los 90 días; a los 210 días estaban representados siete órdenes. En las bolsas que contenían la hojarasca de leucaena se encontró una mayor frecuencia de estos desde el primer mes de colocadas en el pastizal y, por lo general, hubo riqueza de órdenes (entre seis y siete organismos) excepto a los 60, 180 y 210 días. Este comportamiento pudo deberse a la disminución del alimento disponible en las bolsas en la última etapa, pues transcurrido este tiempo existía muy poca hojarasca remanente (fig. 3).
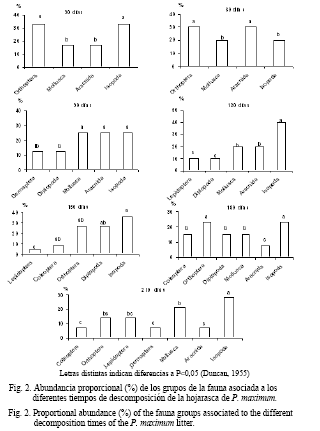
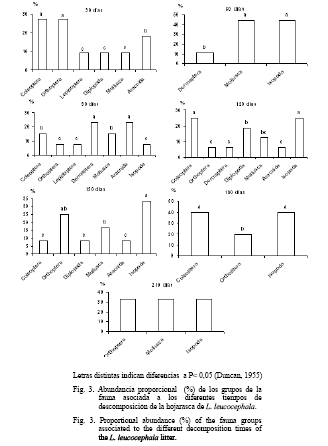
Según Maity y Joy (1999), Tian et al. (2000) y Zimmer y Topp (2002) los invertebrados del suelo difieren entre los tipos de hojarasca, lo cual también pudo estar relacionado con la abundancia de los invertebrados en cada etapa de descomposición. Por otra parte, los organismos de la macrofauna prefieren los restos vegetales algo descompuestos, con una relación carbono: nitrógeno relativamente baja; esto hace que presenten una fuerte selectividad con respecto a la vegetación que existe sobre el suelo (Bastardie y Capowiez, 2004). Los restos de especies con contenidos en nitrógeno superiores a 1,4% son ingeridos con facilidad; mientras que aquellos cuyo contenido es inferior a 1%, les resultan menos atractivos.
Numerosos investigadores determinaron el efecto de la diversidad de la macrofauna en la velocidad de descomposición, mediante la relación entre la frecuencia acumulada de la fauna y la hojarasca remanente en las bolsas (Attignon et al., 2004; Gartner y Cardon, 2004; Shadler y Brandl, 2005). En la figura 4 se muestra la relación entre la frecuencia acumulada y el peso residual de la hojarasca en P. maximum y L. leucocephala en la presente investigación; dicho efecto fue mayor en la leguminosa arbórea.

Estos resultados confirman que la frecuencia acumulada de la fauna durante el tiempo de descomposición de la hojarasca en ambas especies se relaciona con el papel que desempeña en este proceso. En este sentido, Franklin et al. (2005) señalaron que esta alteración puede ser de forma directa a través de la fragmentación de la hojarasca, e indirecta mediante la alteración de la comunidad microbiana y la excreción de nutrientes.
De forma general, los resultados de este estudio permiten concluir que las condiciones de humedad y temperatura que generó el árbol, así como la calidad de su hojarasca, posibilitaron la presencia de una diversa y estable fauna asociada a las bolsas que influyó en el proceso de descomposición. Por ello, el empleo de sistemas que contribuyan a aumentar la diversidad vegetal y la colonización en el suelo de una población biológicamente diversa de organismos, garantiza la mayor sostenibilidad.
REFERENCIAS BIBLIOGRÁFICAS
1. Academia de Ciencias de Cuba. Nuevo Atlas Nacional de Cuba. Instituto Cubano de Geodesia y Cartografía. La Habana, Cuba. p. 41. 1989
2. Anderson, J.M. & Ingram, J. (Eds). Tropical soil biology and fertility. A handbook of methods. 2nd edition. CAB International. Wallingford, UK. 221 p. 1993
3. Attignon, S.E. et al. Leaf litter breakdown in natural and plantation forest of the Lama forest reserve in Benin. Applied Soil Ecology. 27:109. 2004
4. Bartlett, M.S. Properties of sufficiency and statistical tests. Proceedings of the Royal Statistical Society Series A. 160:268. 1937
5. Bastardie, F. & Capowiez, Y. Analysis of earthworm behavior: putting ecological types to test. XIV International Colloquium on Soil Zoology and Ecology. Functional groups and valuation as indicators of soil fauna. Abstracts. p. 236. 2004
6. Brusaard, L. et al. Biodiversity and ecosystem functioning in soil. Ambio. 26:563 1997
7. Cabrera, G. de la C. Caracterización de la macrofauna del suelo en áreas con manejo agroecológico de Cangrejeras, La Habana, Cuba. Tesis en opción al título de Master en Ecología y Sistemática Aplicada. Instituto de Ecología y Sistemática-Ministerio de Ciencia, Tecnología y Medio Ambiente. La Habana, Cuba. 36 p. 2003
8. Caldentey, J. et al. Litter fluxes and decomposition in Nothofagus pumilio stands in the region of Magallanes, Chile. Forest Ecology and Management. 148:145. 2001
9. Cotrufo, M.F. et al. Palatability trials on hardwood leaf litter grown under elevated CO2: a stable carbon isotope study. Soil Biology & Biochemistry. 37:1105. 2005
10. De Aquino, A.M. et al. Diversidade da macroafauna edafica no Brasil. En: Biodiversidade do solo en ecossistemas brasileiros. (Eds. Fátima, M.S. et al. ). Ed. UFLA, Lavras. p. 143. 2008
11. Franklin, E. et al. Relative effects of biotic and abiotic factors on the composition of soil invertebrate communities in an Amazonian savanna. Applied Soil Ecology. 29:259. 2005
12. Gartner, T.B. & Cardon, Z.G. Decomposition dynamics in mixed-species leaf litter. Oikos. 104:230. 2004
13. Hernández, A. et al. Nueva versión de la clasificación genética de los suelos de Cuba. Ministerio de la Agricultura. La Habana, Cuba. p. 26. 1999
14. InfoStat. Software estadístico. Manual de usuario. Versión 1. Córdoba, Argentina. 2001
15. Isaac, S.R. & Naird, M.A. Biodegradation of leaf litter in the warm humid tropics of Kerala, India. Soil Biology & Biochemistry. 37:1656. 2005
16. Lavelle, P. Ecological challenges for soil science. Soil Science. 165:73. 2000
17. Liu, W. et al. Leaf litter decomposition of canopy trees, bamboo and moss in a montane moist evergreen broad-leaved forest on Ailao Mountain, Yunnan, south-west China. Ecol. Res. 15:435. 2000
18. Maity, S.K. & Joy, V.C. Impact nutritional chemical compounds of leaf litter on detritivore soil arthropod fauna. Journal of Ecobiology. 11:193. 1999
19. Mwiinga, R.D. et al. Decomposition of leaves of six multipurpuse tree species in Chipata, Zambia. Forest Ecology and Management. 64:209. 1994
20. Prieto, D. & Rodríguez, C. Fauna edáfica asociada a la hojarasca de la caña de azúcar en la etapa inicial de su descomposición. XV Congreso Latinoamericano y V Cubano de la Ciencia del Suelo. Centro de Convenciones Plaza América, Varadero, Cuba. Boletín 4, p. 68. 2001
21. Rivera, E. & Carrasco, G.V. Estructura trófica de una comunidad de artrópodos epígeos, en un magueyal del Bolsón de Mapimí, Dgo., México (Desierto Chihuahuense). Acta Zool. Mexicana. Nueva Serie. 48:1. 1991
22. Royston, P. An Extension of Shapiro and wilk's test for normality to large samples. Journal of the Royal Statistical Society: Series C (Applied Statistics). 31:115. 1982
23. Shadler, M. & Brandl, R. Do invertebrate affect the disappearance rate of litter mixtures?. Soil Biology and Biochemistry. 37:329. 2005
24. Tian, G. et al. Regeneration of earthworm populations in a degraded soil by natural and planted fallows under humid tropical conditions. Soil Science Society of America Journal. 64:222. 2000
25. Torres, P.A. et al. Microbial succession in litter decomposition in the semi-arid Chaco woodland. Soil Biology & Biochemistry. 37:49. 2005
26. Zimmer, M. & Topp, W. Species-specific utilization of food sources by sympatric woodlice (Isopoda: Oniscidea). Journal of Animal Ecology. 69:1071. 2002
Recibido el 8 de octubre del 2009
Aceptado el 18 de diciembre del 2010
