










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.33 n.1 Matanzas ene.-mar. 2010
ARTÍCULO DE INVESTIGACIÓN
Caracterización cultural y morfológica e identificación de 12 aislamientos fungosos de semillas de Leucaena leucocephala cv. Perú
Cultural and morphological characterization and identification of 12 fungal isolations in seeds of Leucaena leucocephala cv. Peru
J. C. Lezcano1, B. Martínez2 y O. Alonso1
1Estación Experimental de Pastos y Forrajes"Indio Hatuey"Central España Republicana, CP 44280, Matanzas, Cuba
E-mail: juan.lezcano@indio.atenas.inf.cu
2Centro Nacional de Sanidad Agropecuaria, La Habana, Cuba
RESUMEN
La presente investigación tuvo como objetivo identificar los agentes fungosos asociados a las semillas de Leucaena leucocephala cv. Perú almacenadas al ambiente, a partir de la caracterización cultural y morfológica de 12 aislamientos puros; estos se obtuvieron de la siembra de las estructuras fúngicas (vegetativas y/o reproductivas), en placas Petri (9 cm de diámetro) que contenían Agar Papa Dextrosa (APD) y Agar Malta (AM). Las placas se incubaron durante 10 días a 25ºC, con alternancia de 8 h luz/16 h oscuridad o a oscuridad constante, según los requerimientos de cada organismo. Se identificaron siete agentes fungosos asociados a las semillas, los cuales se clasificaron teniendo en cuenta los caracteres culturales y morfológicos, y se corroboraron con las claves taxonómicas. Ello permitió agrupar: Penicillium expansum Link, Rhizopus stolonifer Ehrenb. ex Fr., Cladosporium sphaerospermum Penz., Chaetomium indicum Corda, Alternaria alternata (Fr) Keissl., Pestalotia sp. y Trichoderma sp. Dichos caracteres constituyen una herramienta importante para la identificación de los hongos hasta el nivel de especie, por lo que se recomienda realizar nuevos estudios con los aislamientos #9 y #11 con vista a completar la identificación hasta la especie en el caso de los géneros Trichoderma y Pestalotia; así como identificar las especies fungosas del resto de los aislamientos.
Palabras clave: Hongos, Leucaena leucocephala cv. Perú, semillas.
ABSTRACT
The objective of this study was to identify the fungal agents associated to seeds of Leucaena leucocephala cv. Peru stored under ambient conditions, from the cultural and morphological characterization of 12 pure isolations; they were obtained from planting the fungal structures (vegetative and/or reproductive), in Petri dishes (9 cm diameter) which contained Potato Dextrose Agar (PDA) and Malt Agar (MA). The dishes were incubated for 10 days at 25ºC, alternating 8 h light/16 h darkness or at constant darkness, according to the requirements of each organism. Seven fungal agents associated to the seeds were identified, which were classified taking into consideration the cultural and morphological characteristics, and they were corroborated with the taxonomic keys. This allowed to group: Penicillium expansum Link, Rhizopus stolonifer Ehrenb. Ex Fr., Cladosporium sphaerospermum Penz., Chaetomium indicum Corda, Alternaria alternata (Fr) Keissl., Pestalotia sp. and Trichoderma sp. Such characteristics constitute an important tool for the identification of fungi to the species level, for which new studies with isolations #9 and #11 are recommended in order to complete the identification to species level in the case of genera Trichoderma and Pestalotia; as well as to identify the fungal species of the other isolations.
Key words: Fungi, Leucaena leucocephala cv. Peru, seeds
C
INTRODUCCIÓN
La sanidad de las semillas es un elemento a tener en cuenta en el proceso productivo de Leucaena leucocephala cv. Perú, pues de ella depende en gran medida el éxito de su propagación, comercialización y perpetuación en la naturaleza.
Sin embargo, en la literatura científica internacional no es común encontrar referencias que aborden el tema con profundidad y la mayoría se restringe a informes de la lista de microorganismos patógenos que afectan dicha planta forrajera en los diferentes países, como es el caso del informe realizado por Lenné y Boa (1994) sobre las enfermedades de los árboles leguminosos, donde resaltan la presencia de Camptomeris leucaenae, Ganoderma licidum y Fusarium oxysporum entre los más importantes presentes en L. leucocephala.
En tal sentido, en Cuba en los últimos años se ha informado la presencia de numerosos microorganismos que no solo afectan la producción de semilla de esta leguminosa en condiciones de campo, sino que también causan considerables pérdidas que aceleran el deterioro de su calidad y pueden ocasionarle hasta su muerte en el almacén donde se depositan para su conservación (Lezcano et al., 2007), lo cual se debe fundamentalmente a la decoloración, la alteración del olor y su aspecto (por pudrición) y la disminución del poder germinativo y su viabilidad, por solo citar algunos ejemplos.
Dentro de los agentes patógenos asociados a las semillas de L. leucocephala cv. Perú, los hongos constituyen el grupo más numeroso y uno de los de mayor importancia. Sin embargo, los estudios relacionados con la identificación de estos agentes en Cuba no han sido completados, pues en la literatura consultada solo se hace referencia a dos investigaciones preliminares realizadas por Delgado et al. (1989) y Alonso et al. (1996), quienes solamente clasificaron, hasta nivel genérico, los 18 hongos que encontraron en las semillas de este cultivar, entre los que se encuentran: Trichoderma sp., Rhizopus sp., Phoma sp., Drechslera sp., Chaetomium sp., Gonatobotrys sp., Alternaria sp., Penicillium sp., Aspergillus sp., Fusarium sp., Cladosporium sp., Botrytis sp., Curvularia sp., Doratomyces sp., Helmisthosporium sp., Macrophomina sp., Nigrospora sp. y Rhizoctonia sp.
Por tal motivo, es necesario la búsqueda de diversos métodos de diagnóstico, como la caracterización cultural y morfológica (Cruz et al., 2004; Bernal et al., 2005), que a través de la descripción in vitro de los caracteres más importantes de los agentes saprofitos o patógenos, posibiliten la identificación hasta el nivel de especie, conjuntamente con el empleo de las claves taxonómicas. Dicho aspecto constituyó el objetivo fundamental del presente trabajo, que por su importancia debe ser un elemento a considerar en los momentos actuales cuando la utilización de L. leucocephala cv. Perú se ha incrementado, máxime si se tiene en cuenta que su semilla botánica constituye una vía económica para la diseminación de esta arbórea en el país (Pérez et al., 2000).
MATERIALES Y MÉTODOS
Material vegetal. Semillas de L. leucocephala cv. Perú con seis y 18 meses de almacenamiento al ambiente, procedentes del almacén de la finca de semillas "La Rioja" perteneciente a la Empresa Pecuaria "José Martí" en la provincia de Matanzas, Cuba.
Lugar donde se realizaron las investigaciones. Laboratorio de Micología Vegetal del Centro Nacional de Sanidad Agropecuaria (CENSA) en La Habana, Cuba.
Aislamiento de los hongos y obtención de cultivos puros. Las diferentes estructuras fúngicas (vegetativas y/o reproductivas) halladas en las semillas cuando se sometieron al test de patología (Blotter Test) recomendado por el ISTA (1999), se observaron con un estereomicroscopio Zeiss SV-6, utilizando lentes oculares desde 10 hasta 100 x. Con el empleo de agujas previamente desinfectadas se sembraron en placas Petri de cristal, estériles, de 9 cm de diámetro, las cuales contenían medio Agar Papa Dextrosa (APD), y se incubaron a 28ºC por 7 días. Al finalizar este período, de los 32 cultivos se procedió al montaje de las preparaciones fijas en portaobjetos para su observación al microscopio óptico (Zeiss Estandar 25), y de acuerdo con las estructuras se agruparon, por género, los 12 aislamientos seleccionados.
Para la obtención de los cultivos puros, de cada placa Petri se extrajo un disco micelial de 7 mm de diámetro de los 12 aislamientos, que se transfirieron sobre cuñas de APD en tubos de cristal de 22 x 165 mm con vista a su conservación. Posteriormente se hizo una nueva siembra de cada aislamiento en placas Petri y a los 10-15 días de edad de los cultivos se extrajo un disco micelial de 7 mm de diámetro, que se colocó en el centro de tres placas (repeticiones) para realizar las mediciones.
Dichas placas se sellaron con papel parafinado (Parafilm) y se incubaron durante 10 días a 25ºC, con alternancia de 8 h luz/16 h oscuridad o a oscuridad constante, según los requerimientos para cada organismo.
Caracterización cultural. Se tuvo en cuenta: el color de la colonia en el anverso y el reverso de la placa, la textura, el crecimiento lineal (expresado en cm/día) y el crecimiento aéreo. En el caso del crecimiento micelial lineal de cada aislamiento, se midió el diámetro de las colonias (dos mediciones diarias, a las 8:00 a.m. y 4:00 p.m. hasta el décimo día); mientras que la coloración de la colonia se determinó por la tabla de colores de Rayner (1970).
Los medios de cultivo (pertenecientes a la firma Merck) responden a los criterios de la literatura científica, reflejados en la primera, tercera, cuarta y séptima de las claves taxonómicas que se relacionan posteriormente, referentes a cada género fungoso, según la agrupación de acuerdo con sus estructuras vegetativas y reproductivas.
Los aislamientos #8, #11, #15, #16, #24 y #32 se cultivaron en Agar Papa Dextrosa (APD); el #1, #13 y #14 en Agar Czapek (AZ) y en Agar Malta el #1, #9, #12, #13, #14 y #23.
Caracterización morfológica. Se tuvo en cuenta el tamaño (en µm), la forma y el color de las estructuras vegetativas y reproductivas en los cultivos de los 12 aislamientos. Por cada estructura se realizaron 50 mediciones, en un microscopio óptico Zeiss Estándar 25 (lentes oculares 10-100 x), con una escala micrométrica, con el objetivo de corroborar la clasificación de la especie fungosa, según las claves taxonómicas.
Claves taxonómicas utilizadas para la identificación de los hongos. Malone y Muskett (1964), Onions (1966), Rifai (1969), Ellis (1971), Lunn (1977), Sutton (1980), Barnett y Hunter (1999), Ho et al. (1999) y López et al. (1999).
RESULTADOS Y DISCUSIÓN
Como resultado de este estudio, a continuación se hace referencia a la caracterización cultural y morfológica de los 12 aislamientos analizados, con vista a la identificación de las especies fungosas.
Como resultado de este estudio, a continuación se hace referencia a la caracterización se hace referencia a la caracterización cultural y morfológica de los 12 aislamientos analizados, con visita a la identificación de las especies fungosas.
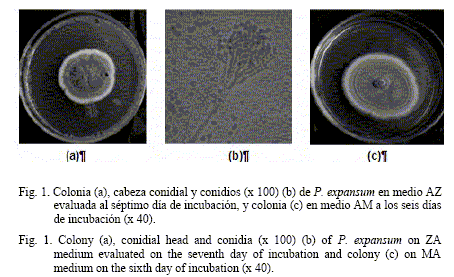
En el caso de los aislamientos #1, #13 y #14, culturalmente manifestaron un comportamiento similar en los medios donde se cultivaron (AZ y AM). En las placas con cultivos sobre AZ incubadas a 25ºC, en el anverso produjeron colonias de crecimiento rápido (16 mm de diámetro al séptimo día de incubación) y de color verde grisáceo, a menudo, con una zona radial con bordes regulares de color blanco, los que llegaron a tornarse verde grisáceo con la producción de los conidios.
En el reverso las colonias generalmente se observaron incoloras, aunque también un aislamiento (el #13) presentó color pardo con tonalidades amarillas.
En medio AM las colonias de estos aislamientos incubadas a 25ºC, en el anverso de las placas se observaron afieltradas y de color verde grisáceo, con igual zona radial blanca, polvosa, de crecimiento aéreo escaso y superficial moderadamente abundante (40 y 50 mm al séptimo y al décimotercer día de incubación, respectivamente), lo cual corrobora lo planteado por Onions (1966) cuando expresó que las colonias de este hongo crecen con mayor rapidez en el medio AM que en el de AZ. Por el reverso las colonias no mostraron una distinción clara y tuvieron tendencia a ser incoloras, al igual que en el medio AZ.
Desde el punto de vista morfológico, las principales características de las estructuras, provenientes de los cultivos sobre AZ, fueron: la producción de cabezas conidiales asimétricas, ramificadas, alargadas y con largas cadenas de conidios. Por otra parte, presentaron conidióforos peniciliados con fialides, lisos, moderadamente largos de 345-371 x 3,02-3,16 µm (largo por ancho); ramas de 13-21 x 2,0-2,9 µm y fialides en grupos de cinco, que pueden llegar hasta nueve, con tamaño variable, de 8-12 x 2-2,5 µm y en ocasiones más largas. Los conidios se observaron lisos y elípticos, con un tamaño de 3,68-4,09 x 2,68-3,04 µm.
Por su parte, las estructuras procedentes de los cultivos en AM fueron similares a las descritas anteriormente, con la única diferencia de que las cabezas conidiales pueden poseer una o dos ramas.
Ambos grupos de características coinciden con lo expuesto por Onion (1966) y CABI (2003) para la especie Penicillium expansum Link; de ahí que exista correspondencia con la encontrada en este estudio (fig. 1).
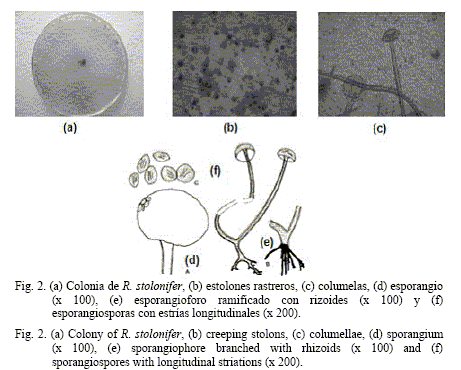
A partir de los aislamientos #8, #15 y #16, que crecieron en APD y fueron incubados a 25ºC, se obtuvieron colonias blancas algodonosas cuando jóvenes, las que se tornaron de color negro parduzco con la madurez, por la presencia de los esporangios. Un aspecto importante en todos los aislamientos fue la rápida diseminación que mostraron las colonias en los cultivos, al cubrir las placas Petri a las 24-48 h. Esta situación se debió a la presencia de estolones rastreros fijados a través de los rizoides en varios puntos del sustrato.
Como caracteres morfológicos de estos aislamientos se observaron esporangioforos de 30-34 µm de ancho por 900-3 100 µm de largo, de paredes lisas, no septados, de color pardo claro, simples, que a veces surgían en grupos de tres a cinco, a partir de estolones y rizoides opuestos. Los esporangios, de globosos a subglobosos, blancos al principio y negros después, medían de 100-350 µm de diámetro. Las columelas eran subglobosas a aplanadas, grises parduzcas, en forma de sombrillas cuando estaban hendidas, con una dimensión de 60-198 x
67-135 µm. Los rizoides y los estolones eran hialinos y en ocasiones aparentaron ser pardo oscuros. Las esporangiosporas presentaron diversas formas: alargadas, ovaladas y redondas, pero casi siempre irregulares y de tamaño variable (13-17 µm).
Lo descrito con anterioridad coincide con lo informado por Malone y Muskett (1964) y Lunn (1977) acerca de la especie Rhizopus stolonifer (Ehremb. Ex Fr) Lind., por lo que también se clasificó así este agente fungoso (fig. 2).
Las colonias de los aislamientos #12 y #23 sobre medio AM e incubadas a 25ºC, desde el punto de vista cultural al séptimo día se caracterizaron por mostrar en el anverso una coloración de olivácea gris pálido a olivácea gris oscura, polvosas, con un crecimiento superficial lento (27 µm); en el reverso presentaron una coloración negro olivácea. Los micelios se observaron hialinos, septados, lisos, de 3-4 µm de ancho; las hifas mostraron igual coloración que los micelios, aunque posteriormente se tornaban oscuras a medida que avanzaba la edad de la colonia. Los conidióforos eran de color oliváceo claro a oliváceo, formados sobre las hifas, semimacronematosos, rectos y no ramificados, lisos y septados, con una dimensión de 135-140 µm de largo, aunque en ocasiones se observaron de menor tamaño, de 40 hasta 55 µm y con un ancho de 2,7 a 3,2 µm.
En cuanto a los conidios, se caracterizaron por ser de color olivo claro, equinulados y globosos en su gran mayoría, los cuales aparecen en cadenas simples o ramificadas y pueden presentar hasta un septo, de tamaño variable (6,1-7,9 x 3,05-6,03 µm); ramoconidios subglobosos, de color olivo claro, septados (de dos a tres septos) y de 11,07-14,16 µm de largo x 5,5-6,3 µm de ancho.
De acuerdo con las características informadas con anterioridad acerca de estos aislamientos, es de destacar que coinciden con las expuestas por Malone y Muskett (1964) y Ho et al. (1999) para la especie Cladosporium sphaerospermum Penz, lo cual se muestra en la figura 3.

Respecto a las colonias del aislamiento #32 cultivadas en APD, se caracterizaron culturalmente por presentar una coloración blanca, con bordes irregulares en el anverso; el crecimiento lineal fue lento (15 y 22 mm al octavo y décimotercer día de incubación a 25ºC, respectivamente) y el crecimiento aéreo, escaso; se observó un micelio vegetativo sumergido, de apariencia granular, dado quizás por la producción de peritecios superficiales, los cuales reunían las siguientes características morfológicas: color negro, globosos, con un diámetro de 91 a 112 µm y poseían pelos carmelitas con tonalidades casi negras; los pelos laterales eran rectos, más bien pardos, de aproximadamente 3,5 mm, de mayor grosor en la base y menor hacia el ápice hialino; mientras que los terminales tenían color pardo oscuro en su gran mayoría, no septados y se ramificaban de forma repetida. Las ascosporas eran hialinas cuando jóvenes y se tornaban carmelita al envejecer, presentaban forma ovalada u ovoide y medían de 4,30-5,73 µm de largo x 4,0-4,78 µm de ancho.
Acorde con las características descritas con anterioridad se pudo corroborar lo planteado por Malone y Muskett (1964) y Rodríguez (2003) para el género Chaetomium y en particular para la especie Chaetomium indicum Corda, y por consiguiente su identificación en las semillas en estudio (fig. 4).

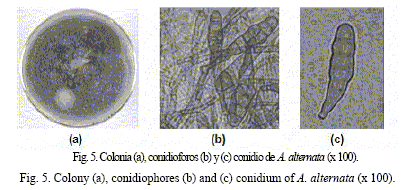
Con relación a la caracterización cultural del aislamiento #24 cultivado en APD, en el anverso de las placas se observaron colonias de color pardo grisáceo a gris, con bordes regulares, crecimiento aéreo escaso y lineal abundante (7 cm al séptimo día de incubación a 25ºC). Entre los caracteres morfológicos se observaron los siguientes: las hifas en algunas ocasiones se encontraron inmersas en el medio y, en otras, superficiales; eran más o menos cilíndricas, tabicadas, ramificadas en ángulos casi rectos y presentaron una coloración con tonalidades carmelitas. Los conidioforos a menudo surgen solitarios o en pequeños grupos, son ramificados, rectos o flexuosos, lisos, de color carmelita, con un tamaño de 3 a 6 µm de ancho y con uno o varios conidios. Dichos conidios se forman en largas cadenas o a menudo en solitario, son obclavados, ovoides o elipsoidales, con un pico cilíndrico o cónico corto (de color pálido, de 2 a 4 µm de ancho), lisos o verruculosos, de color carmelita con aproximadamente hasta ocho septos transversales y usualmente varios longitudinales, de 18-43 x 9-16 µm.
Estos resultados del aislamiento #24 son similares a los obtenidos por Ellis (1971), López et al. (1999) y Cundóm y Cabrera (2002) para la especie Alternaria alternata (Fr) Keissler, lo que indica que es una de las especies presentes en las semillas de L. leucocephala cv. Perú (fig. 5).
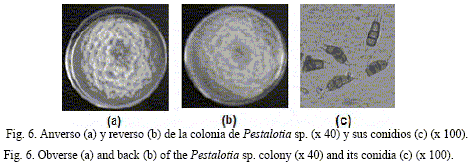
En cuanto al aislamiento #11, su caracterización cultural se realizó en placas Petri en APD; en el anverso de estas se observaron colonias de color blanco cremoso, algodonosas, en forma de rosetas, de crecimiento superficial abundante (85 mm a los siete días de incubación a 25ºC) y aéreo escaso, en cuyo centro se observó la presencia de acérvulos oscuros; mientras que en el reverso las colonias mostraron una coloración blanca con tonalidades amarillas.
Por otra parte, como características morfológicas se observaron: la presencia de conidioforos hialinos, irregularmente ramificados, septados, lisos y cortos; así como de conidios oscuros, rectos o ligeramente curvados, que poseían seis células (con la basal y la terminal hialina, y esta última puntiaguda con dos o más apéndices apicales hialinos, también elipsoidal o fusiforme).
Como resultado de esta caracterización se corroboró lo descrito por Sutton (1980), Barnett y Hunter (1999) y Montiel y Avelar (2001) para el género Pestalotia, por lo que se identificó el agente fungoso correspondiente al aislamiento #11 como Pestalotia sp. (fig. 6).
El aislamiento #9 sembrado en placas Petri con AM, se caracterizó por producir colonias algodonosas de color blanco en el anverso al inicio, las que se tornaron verde rápidamente a medida que transcurría el período de incubación y presentaban bordes regulares de crecimiento radial uniforme (aproximadamente 6 cm al cuarto día de incubación); mientras que el reverso de la colonia permaneció incoloro.
En cuanto a su caracterización morfológica se observó que el micelio estaba compuesto por hifas hialinas, septadas, ramificadas, de paredes lisas y de 4,6-5,13 µm de diámetro. Se observaron clamidosporas intercalares, principalmente globosas, de paredes lisas, incoloras y de 6-12 µm de diámetro. Los conidioforos se encontraron ramificados y no terminaban en punta de micelio, sino en fialides, las cuales tenían forma de botella. Los conidios se corresponden con fialosporas ovoides a subglobosas, de color verde pálido, aparentemente lisas, que se producen solas y sucesivamente se acumulan en el extremo de cada fialide para formar una cabeza conidial globosa; miden de 2,8-3,2 x 2,3-2,8 µm, con un promedio de 2,94 x 2,67 µm.
Teniendo en cuenta las características observadas del aislamiento #9, se puede plantear que se corresponden con las citadas por Rifai (1969), Samuel et al. (1998) y Barnett y Hunter (1999) para las especies del género Trichoderma, por lo que este agente fungoso se clasifica como Trichoderma sp. (fig. 7).

Después de realizar la identificación de las especies fungosas se comprobó que P. expansum, C. sphaerospermum y C. indicum son informados por primera vez para la especie forrajera L. leucocephala; mientras que A. alternata, conjuntamente con las anteriores, también lo son para el cultivar Perú en Cuba, ya que según la revisión bibliográfica de los principales trabajos publicados en el país (Delgado et al.,1989; Pazos, 1989; Alonso et al., 1996; Pupo y Heredia, 1998; González et al., 2006), no se citan con anterioridad.
CONCLUSIÓN
De acuerdo con la caracterización cultural y morfológica de los 12 aislamientos, se identificaron siete especies fungosas presentes en las semillas almacenadas de L. leucocephala cv. Perú: Penicillium expansum Link, Rhizopus stolonifer Ehrenb. ex Fr., Cladosporium sphaerospermum Penz., Chaetomium indicum Corda, Alternaria alternata (Fr) Keissl., Pestalotia sp. y Trichoderma sp., lo que ratifica que dichos caracteres constituyen una herramienta importante para la identificación hasta la categoría de especie cuando se comparan con lo descrito en las claves taxonómicas. Además, es válido destacar que la primera, tercera, cuarta y quinta especies fungosas constituyen nuevos informes para esta leguminosa forrajera en el país.
RECOMENDACIONES
Continuar los estudios de los aislamientos #9 y #11, con vista a completar la identificación hasta la categoría de especie en los géneros Trichoderma y Pestalotia.
Identificar las especies fungosas correspondientes al resto de los aislamientos.
REFERENCIAS BIBLIOGRÁFICAS
1. Alonso, O. et al. Hongos asociados a las semillas de una leguminosa tropical (Leucaena leucocephala cv. Perú). Pastos y Forrajes. 19 (2):161. 1996
2. Barnett, H.L. & Hunter, B.B. Illustrated genera of imperfect fungi (fourth edition). The American Phytopathological Society. Pilot Knob Rodal, St Paul, Minnesota, USA. 216 p. 1999
3. Bernal, A. et al. Caracterización cultural de tres aislamientos de Cladosporium fulvum Cooke. Centro Agrícola. 32 (3):93. 2005
4. CABI. Seed-borne disease. In: Crop protection compendium. (cd-rom). CAB International, UK. 2003
5. Cruz, Mileydi et al. Caracterización de aislados de Pseudocercospora fijiensis Morelet para su utilización en programas de mejoramiento de Musa sp. Memorias. Seminario Científico Internacional de Sanidad Vegetal. 44 Reunión Anual de la Sociedad Americana de Fitopatología. División Caribe. La Habana, Cuba. [cd-rom] . 2004
6. Cundóm, M.A. & Cabrera. M.G. Nota corta: Tizón foliar de Celosia argentea en el nordeste de Argentina. Invest. Agr.: Prod. Prot. Veg. 17 (3):463. 2002
7. Delgado, A. et al. Estudio de la gomosis bacteriana en legumbres de Leucaena leucocephala (Lam) de Wit. Pastos y Forrajes. 12:127. 1989
8. Ellis, M.B. Demateaceous Hyphomycetes. Commowealth Micological Institute. Kew, Surrey, England. 1971
9. González, Gloria et al. Fitopatógenos en los cultivos de pastos y forrajes en Cuba. Fitosanidad. 10 (1):11. 2006
10. Ho, M.H.M. et al. Cladosporium sphaerospermum. In: Cladosporium and Cladophialophora in culture: descriptions and an expanded key. Mycotaxon. 72:115. 1999
11. ISTA. International rules for seed testing. Seed Science and Technology. Supplement. 27. 302 p. 1999
12. Lenné, Jilliam M. & Boa, E.R. Diseases of tree legumes. In: Forage tree legumes in tropical agriculture. (Eds. Gutteridge, R.C. & Shelton, H. M.). CAB International, UK. p. 292. 1994
13. Lezcano, J.C. et al. Determinación de la calidad de las semillas de Leucaena leucocephala cv. Perú almacenadas al ambiente. Pastos y Forrajes. 30 (1):107. 2007
14. López, María O. et al. Manual para la identificación de los hongos fitopatógenos de la caña de azúcar en Cuba. Acta Botánica Cubana. 124:75. 1999
15. Lunn, J. Rhizopus stolonifer. In: Descriptions of pathogenic fungi and bacteria. Commonwealth Mycological Institute (CAB) (Set 53). Kew, Surrey, England. # 524. s/p. 1977
16. Malone, J.P. & Muskett, A.E. Seed-borne fungy description of 77 fungus species. In: Proceedings of the International Seed Testing Association. Edité par l´Association Internationale d´Essais de Semences. Wageningen, Holand. Vol. 29, No. 2, 383 p. 1964
17. Montiel, Rosaura D. & Avelar, J.J. Etiología de la enfermedad "clavo del guayabo". Trabajo presentado en la 5ta Jornada de Investigación, 25 al 29 de junio. Universidad Autónoma de Zacatecas, México. 11 p. 2001
18. Onions, A.H.S. Penicillium expansum. In: Descriptions of pathogenic fungi and bacteria. Commonwealth Mycological Institute (CAB) (Set 10). Kew, Surrey. England. # 97. s/p. 1966
19. Pazos, R. Plagas, enfermedades y malezas en pastos. Documento de campo. Proyecto PNUD/FAO-CUB/86/005. Instituto de Investigaciones de Pastos y Forrajes-MINAG. La Habana,Cuba. 41 .1989
20. Pérez, A. et al. Tecnología para la producción de semillas de gramíneas y leguminosas. En: Tecnología para la producción de semillas de pastos tropicales. Estación Experimental de Pastos y Forrajes "Indio Hatuey" Matanzas, Cuba. p. 17. 2000
21. Pupo, Elsie & Heredia, Irma. 1989. Lista de hongos asociados a las semillas de Leucaena leucocephala. Boletín técnico. No. 1. Instituto de Investigaciones de Sanidad Vegetal, Cuba. p. 26
22. Rayner, R.W. A mycological colour chart. Commonwealth Mycological Institute. Kew, Surrey, England. s/p. 1970
23. Rifai, M.A. A revision of the genus Trichoderma. Micological Paper. No. 116, p. 1. 1969
24. Rodríguez, Kendra. Estudio taxonómico (morfológico y molecular) de especies del género Chaetomium y géneros afines. Tesis presentada en opción al grado de Doctor en Ciencias Agrícolas. Universitat Rovira i Virgili. Facultad de Medicina I Ciencies de la Salut. Departament de Ciencias Médiques. Unitat de Biología. España. 173 p. 2003
25. Samuel, G.J. et al. The Hypocrea schweinitzii complex and Trichoderma sect. Longibrachiatum. Studies in Mycology. 41. Centraalbureau voor schimmelculture, Baarn/delft. The Netherlands Academy of Arts and Science. 53 p. 1998
26. Sutton, B. Pestalotia de Not. In: The coleomycetes fungi imperfecti with picnidia acervuli and stromata. Commonwealth Mycological Institute (CAB) (Set 10). Kew, Surrey, England. p. 263. 1980
Recibido el 8 de junio del 2009
Aceptado el 20 de noviembre del 2009
