










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.33 n.1 Matanzas ene.-mar. 2010
ARTÍCULO DE INVESTIGACIÓN
Dinámica de las larvas infestantes de estrongílidos gastrointestinales en ovinos en pastoreo
Dynamics of gastrointestinal strongyle infesting larvae in grazing sheep
J. Arece1 y J.G. Rodríguez2 1Estación Experimental de Pastos y Forrajes «Indio Hatuey» . Central España Republicana CP 44280, Matanzas, Cuba
E-mail: javier.arece@indio.atenas.inf.cu
2Centro Nacional de Sanidad Agropecuria (CENSA), La Habana, Cuba
RESUMEN
Se desarrolló un estudio durante dos años, con el objetivo de caracterizar la dinámica de las poblaciones larvarias en «refugio» de nemátodos gastrointestinales, así como su interrelación con las variables meteorológicas, en tres unidades de producción ovina de la provincia de Matanzas (Dos Mercedes, LABIOFAM y la EEPF «Indio Hatuey»). Con una frecuencia mensual se recolectaron e identificaron las larvas de estrongílidos gastrointestinales, mediante una técnica de lavado del pasto proveniente de cuartones centinelas previamente seleccionados. La dinámica de las poblaciones larvarias de tercer estadio tuvo un comportamiento similar en las unidades en estudio; sin embargo, hubo diferencias en las cantidades de L3 por kg de MS de pasto. Se registró un efecto de los meses y la época del año en la disponibilidad de larvas. En el período lluvioso (PLL) se recolectó una mayor cantidad de L3 como producto de los efectos favorables de las precipitaciones y la temperatura. En el pasto se encontró un predominio de L3 de Haemonchus spp. y, en menor cuantía, Trichostrongylus colubriformis y Oesophagostomum columbianum. Las poblaciones larvarias aparecieron en mayor cuantía durante el PLL y su dinámica se relacionó con las variables ambientales y con los factores vinculados al sistema de producción en cada unidad.
Palabras clave: Haemonchus spp., larvas, ovinos
ABSTRACT
A study was conducted for two years, with the objective of characterizing the dynamics of the larval populations in «refuge» of gastrointestinal nematodes, as well as its interrelation with the meteorological variables in three sheep production farms in Matanzas province, Cuba (Dos Mercedes, LABIOFAM and the EEPF «Indio Hatuey»). Larvae of gastrointestinal strongyles were monthly collected from pasture and identified by means of a pasture washing technique from previously selected sentinel paddocks. The third stage larval population's dynamics in pasture showed a similar trend in the three farms; however, there were differences in the amounts of L3, per kg DM of pasture. A significant effect of month of the year and season on larvae availability was recorded. In the rainy season (RS) a higher quantity of L3 was collected due to the favorable effects of rainfall and temperature. Haemonchus spp. was the prevailing species, followed by Trichostrongylus colubriformis and Oesophagostomum columbianum. The larval populations showed higher quantity during the RS and their dynamics was related to the environmental variables and the factors related to the production system in each unit.
Key words: Haemonchus, larvae, sheep
INTRODUCCIÓN
En Cuba los sistemas de explotación de ovinos basan su alimentación en el aprovechamiento de los pastos. Como resultado del proceso de pastoreo existe un enorme riesgo de adquisición de parásitos del sistema digestivo, ya que es allí donde completan parte de su ciclo biológico (Cuéllar, 2002).
En la actualidad una de las premisas fundamentales para el incremento de la productividad y rentabilidad de las explotaciones ovinas es la disminución de los costos asociados con las endoparasitosis. En este contexto, la búsqueda de alternativas de control parasitario constituye un elemento clave (González, 2004).
El conocimiento de las fluctuaciones estacionales de los estadios larvarios constituye un elemento principal al diseñar una estrategia de control parasitario, ya que se puede fijar un modelo de contaminación de los pastizales y de este modo trazar las posteriores medidas preventivas y curativas (García-Romero et al., 1995). Por tanto conocer la población larvaria en «refugio1» es importante para el mantenimiento de la eficacia de los medicamentos antiparasitarios (Sissay et al., 2006).
Las investigaciones referentes a las poblaciones de larvas del tercer estadio de los nemátodos gastrointestinales de ovinos realizadas en la región occidental de Cuba hasta la actualidad se refieren, fundamentalmente, a los estudios de contaminación puntual y de desarrollo y sobrevivencia de estas en el medio ambiente (Delgado, 1983; García et al., 1994); sin embargo, se han realizado escasos trabajos con el objetivo de caracterizar la dinámica poblacional de las L3 en el pasto, así como su interrelación con las variables meteorológicas en condiciones de producción dentro de un estudio integral del parasitismo interno en ovinos.
MATERIALES Y MÉTODOS
Características de las unidades
Se seleccionaron tres unidades de producción de la provincia Matanzas, las cuales son representativas de los principales sistemas de producción ovina.Finca Dos Mercedes
Localización. Empresa Pecuaria Genética de Matanzas, ubicada en el Consejo Popular Manuelito del municipio Pedro Betancourt. Se dedica exclusivamente a la explotación de ovinos y es considerado un centro multiplicador del ovino Pelibuey, de referencia en la provincia.
Alimentación. Se basa, fundamentalmente, en la utilización de los pastos naturales (Dichanthium-Bothriochloa-Paspalum-Sporobolus ). Como estrategia de desarrollo en el período poco lluvioso, los animales reciben aproximadamente 5 kg/animal/día de king grass (Pennisetum purpureum), dependiendo de la disponibilidad en las áreas forrajeras.
Animales y manejo. El rebaño está constituido por 145 ovinos agrupados en diferentes categorías (reproductoras, crías, animales en desarrollo-crecimiento y sementales). El área de pastoreo es de 9 ha divididas en grandes cuartones, en los cuales los animales se llevan a pastorear en grupos homogéneos por categoría. Los cuartones se intercambian entre los grupos existentes, excepto los sementales que pastan en la misma área durante todo el año. Las crías permanecen estabuladas durante el período de amamantamiento y se llevan una vez por semana junto con las reproductoras al pastoreo. Los animales pastorean de siete a ocho horas diarias y en las noches se trasladan a la nave de tenencia, donde se mantienen los grupos previamente formados. Esta instalación posee piso de hormigón que permite su limpieza mecánica diaria.
Desparasitaciones. Se realizan de forma arbitraria, sin la existencia de un plan de control parasitario, y en ocasiones se hacen cada 21 días. No existe una rotación de los medicamentos antiparasitarios, por lo que en los tratamientos antihelmínticos se utiliza el producto que exista en un momento dado.LABIOFAM
Localización. Pertenece a la Granja Agropecuaria de los Laboratorios Biológicos Farmacéuticos y está situada en el Valle de Yumurí. Es una granja integral que no se dedica exclusivamente a la explotación ovina, aunque tiene bien limitadas las áreas para cada una de las producciones agropecuarias que en ella se desarrollan.
Alimentación. Se basa en el empleo de pastos naturales como fuente de alimento. Los animales reciben suplementación con forraje de king grass en el período poco lluvioso, en dependencia de la disponibilidad de suplemento. Las instalaciones son de piso de tierra y se barren con determinada frecuencia para impedir la acumulación excesiva de las heces.
Animales y manejo. El rebaño está constituido por 150 ovinos divididos en diferentes categorías (reproductoras, crías, desarrollo-ceba y sementales). Posee un área de aproximadamente 20 ha sin divisiones internas. El pastoreo es libre y los animales se llevan a grandes áreas en las que sólo existe cerca perimetral. El rebaño pastorea bajo el concepto de rebaño único, ya que todas las categorías pastan de forma conjunta. No existe una estrategia concertada de mejora genética y la reproducción es mediante monta libre no controlada, lo que ha conllevado un alto grado de consanguinidad visible en los animales.
Desparasitaciones. Se realizan de forma arbitraria, sin la existencia de un plan de control parasitario, y los productos se aplican en función del fármaco disponible.EEPF «Indio Hatuey»
Localización. Pertenece al área de producción animal de la Estación Experimental de Pastos y Forrajes «Indio Hatuey», ubicada en el municipio Perico. Es una finca diversificada que posee bien delimitadas las áreas e instalaciones dedicadas a la explotación ovina.
Alimentación. Se basa en el aprovechamiento de los pastos naturales. En el período poco lluvioso se emplean diferentes fuentes suplementarias de alimentos, como pulpa de cítrico y follaje de morera, entre otras. Las instalaciones poseen piso de concreto, lo cual posibilita su limpieza mecánica diaria.
Animales y manejo. El rebaño está constituido por 120 animales separados en diferentes categorías (reproductoras, crías, desarrollo-ceba y sementales), las cuales pastan en diferentes áreas (4,5 ha) en dependencia de la categoría. Los machos se castran y se incorporan al pastoreo con las hembras en desarrollo. La reproducción se organiza por campañas (junio-julio/octubre-noviembre), lo que posibilita la concentración de los partos y facilita el manejo de los animales lactantes y recentinas. Las crías se mantienen estabuladas durante el período de amamantamiento y sólo salen a pastoreo una vez por semana con las madres.
Desparasitaciones. Se aplican sin un plan de control parasitario, por no existir un criterio para su realización. No se rotan los antiparasitarios, sino que su aplicación depende de la disponibilidad en el momento que se decida el tratamiento.
Determinación del nivel de infestación de las L3 en el pasto
El nivel de contaminación de los pastizales se estimó con una frecuencia mensual, durante dos años, mediante la recolección y el conteo de las L3 en el pasto. Para ello se modificó la técnica de lavado del pasto descrita por García-Romero et al. (2000). Las modificaciones se realizaron en la cantidad de material vegetal para el análisis (100 g), el volumen de agua (2 000 mL) y el del alícuota (10 mL). Los muestreos se hicieron en el horario de 8:00-9:00 a.m. Las L3 se identificaron según la clave propuesta por Valle (1978) y Gruner y Raynaud (1980). Se determinó el contenido de MS del pasto y las larvas se expresaron en L3/kg MS.
Información meteorológica
La información meteorológica se registró con una frecuencia mensual durante dos años (2000-2002) y procedía de centros meteorológicos próximos a las entidades donde se realizó el estudio. Se compilaron las siguientes variables: el acumulado de las precipitaciones (mm), los promedios de la humedad relativa (%), la temperatura media (ºC) y la radiación solar global (MJ/m2), con una frecuencia mensual. Por otra parte, se analizó la información de todas las variables obtenida en los últimos 15 años, excepto la radiación solar global.
Análisis y procedimientos estadísticos
Los factores considerados para el análisis fueron la unidad y la época del año (período lluvioso, PLL y período poco lluvioso, PPLL). La cantidad de las L3 en el pasto se transformó [Log (X+1)] para lograr homogeneidad de las varianzas y una distribución normal de los datos, la cual fue evaluada mediante la prueba Bartlett y la no paramétrica Kolmogorov-Smirnov (P<0,05). La retransformación para obtener la media geométrica se hizo como sigue: media geométrica = (10x)-1) (Smothers et al., 1999).
El análisis se realizó con el paquete estadístico SPSSâ versión 10.0.1 para Windows. Se empleó el procedimiento GLM para la disponibilidad de L3 mediante el siguiente modelo matemático:
Yijkl=µ+Ui+Ej + Sk+Tl(Ui x Ej)+eijkl
Donde:
Yijkl= Observación de la disponibilidad de las L3 en el pasto
µ = Media de la población
Ui= Efecto de la iésima unidad (i=3; 1= Dos Mercedes; 2= LABIOFAM; 3= EEPF)
Ej= Efecto de la jésima época (j=2; 1= seca; 2= Lluvia)
Sk=Efecto de la késima especie de estrongílido (k=3; 1= Haem.; 2= Trich.; 3= Oesoph.)
Tl(UixEj)=Efecto de la interacción de la iésima unidad con la jésima época
eijkl= Efecto aleatorio del error asociado con cada observación.
Las diferencias entre las medias se establecieron mediante la dócima de comparación de rangos múltiples de Duncan (1955) y se consideró significativo cualquier valor de P<0,05. Para el estudio de la relación entre las variables meteorológicas y la disponibilidad de las larvas infestantes en el pasto, se realizó un análisis de correlación no lineal de Spearman.
RESULTADOS
Disponibilidad de las L3 en el pasto
Las especies recuperadas en el pasto fueron Haemonchus spp., Trichostrongylus colubriformis y Oesophagostomum columbianum. En la figura 1 se muestra la dinámica de las L3 (L3/kg MS) en las tres unidades en estudio. En ella se puede apreciar la existencia de patrones similares en su comportamiento en las tres localidades, en los cuales los picos de mayor infestación correspondieron a los meses del PLL, cuyos valores indican un ascenso evidente en el inicio de dicho período (mayo-junio). La época del año (tabla 1) mostró diferencias significativas (P<0,001 y P<0,01) en cuanto a la cantidad de las L3 en Dos Mercedes y la EEPF «Indio Hatuey»; mientras que en LABIOFAM apareció una interacción de primer orden entre la época del año y las especies de estrongílidos gastrointestinales (fig. 2).Por otra parte, la mayor disponibilidad total de las L3 se halló en el PLL (P<0,001) para Dos Mercedes y LABIOFAM; también se encontraron diferencias significativas (P<0,001 y P<0,01) en la cantidad del larvas de tercer estadio de Haemonchus spp. a favor del PLL, mientras que para T. colubriformis y O. columbianum no se apreciaron diferencias.
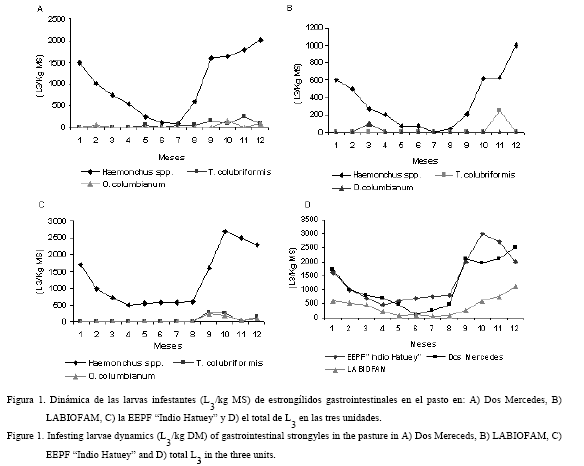

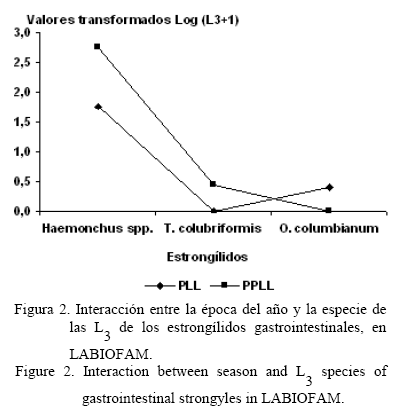
En el caso particular de LABIOFAM se apreció una interacción de primer orden entre la época del año y las especies de estrongílidos gastrointestinales. Este fenómeno fue el resultado de un incremento en la cantidad de las L3 de O. columbianum en el período de mínimas precipitaciones, que superó las encontradas en el PLL.
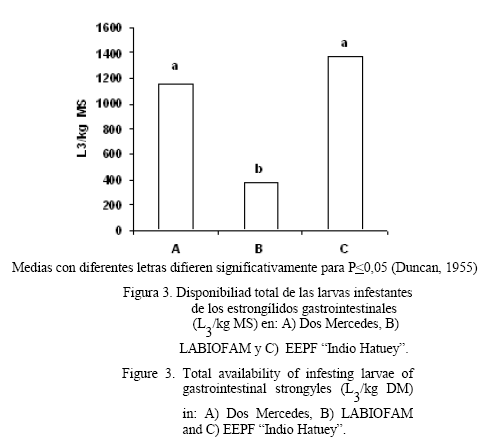
La cantidad de larvas recolectadas en el pasto mostró diferencias entre las especies (tabla 2), con un predominio significativo (P<0,001) de Haemonchus spp. sobre el resto de las larvas infestantes. También se apreciaron diferencias significativas (P<0,01) en la cantidad de L3 en el pasto en las tres unidades (fig. 3), con los mayores niveles de contaminación en la EEPF «Indio Hatuey» y Dos Mercedes.

Relación entre las L3 en el pasto y las variables meteorológicas
En la tabla 3 se muestra la matriz de correlación entre las variables meteorológicas y la cantidad total de las L3 recuperadas en el pasto, en las que se incluye el género Haemonchus y las especies T. colubriformis y O. columbianum. Como se observa, se obtuvo una fuerte relación entre las L3 de Haemonchus spp. y el total de las L3 y las precipitaciones (P<0,01); también se relacionaron directamente con O. columbianum en Dos Mercedes y la EEPF «Indio Hatuey».
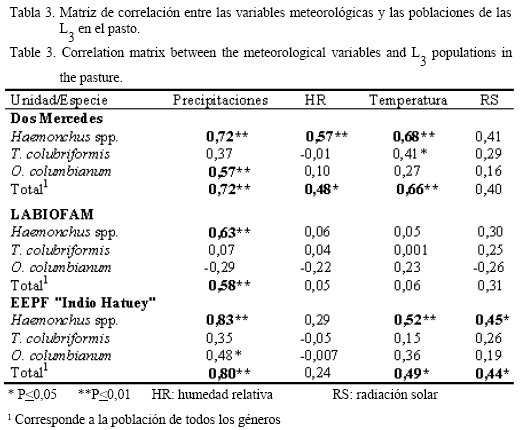
La humedad relativa sólo se relacionó (P<0,05) con las larvas de Haemonchus spp. y el total de las L3 en la finca Dos Mercedes. Por su parte, la temperatura media desempeñó un papel importante en el desarrollo y la sobrevivencia de las larvas de tercer estadio en Dos Mercedes y la EEPF «Indio Hatuey»; mientras que la radiación solar mostró efectos positivos sobre las L3 de Haemonchus spp. y el total en la EEPF «Indio Hatuey».
DISCUSIÓN
Los estrongílidos gastrointestinales encontrados en el lavado del pasto demuestran que los rebaños ovinos en la provincia de Matanzas son afectados por tres especies fundamentales: Haemonchus spp., T. colubriformis y O. columbianum, con predominio del primero. Esto corrobora los resultados de Arece y Rodríguez (2003), quienes consideran que la haemoncosis es el principal problema parasitario de los ovinos en la región.
Los valores promedio de las L3 en las tres unidades fueron moderados, al compararlos con los hallados en estudios similares realizados en Guadalupe (5 450 y 3 520 L3/kg MS para el PLL y el PPLL, respectivamente) por Aumont et al. (1996). Ello indica que en las condiciones de este estudio los animales estuvieron continuamente expuestos a fuentes considerables de infestación durante todo el año, con los mayores riesgos en el PLL.
Se pudo apreciar que la cantidad de las L3 mostró diferencias entre las unidades en estudio, con los mayores conteos en Dos Mercedes y la EEPF «Indio Hatuey». Este comportamiento se puede atribuir a las diferencias en la carga animal en las tres unidades, la cual resultó más baja en LABIOFAM. Por otro lado, es importante señalar que hubo diferencias en el manejo de los tres rebaños, ya que en esta unidad los animales se mantenían en grandes extensiones con pastoreo libre, mientras que en las otras existía acuartonamiento y restricción del pastoreo. En este sentido, Thamsborg et al. (1996) señalaron la existencia de una relación positiva entre la carga animal y la cantidad de las L3 en el pasto, en la cual la mayor carga animal se correspondió con el mayor nivel de contaminación del pasto.
En el presente estudio resultó interesante que los picos de las L3 en el pasto (en las tres unidades) correspondieran a los meses del PLL, mientras que en ese mismo período se encontró la menor cantidad de huevos en las heces (Arece- García et al., 2007). Sin embargo, en el PPLL las poblaciones larvarias fueron bajas (a la altura que pastaban los animales) y los animales mostraron los mayores niveles de infestación parasitaria. Este fenómeno pudiera estar asociado con la instauración de una inmunidad sólida, debido al incremento de la disponibilidad y la calidad del forraje. A su vez, en el PPLL los animales se ven forzados a ingerir el pasto a niveles más bajos, como producto de la disminución en la disponibilidad de alimento, y en este lugar las poblaciones de larvas son más densas (Gruner et al., 1984). Estos resultados coinciden con los de García-Romero et al. (1995), quienes obtuvieron altos CFH con bajas densidades de L3 en el pasto como producto de la posible reinfestación de los animales en lugares puntuales en las áreas de explotación, como son los abrevaderos y las áreas a la entrada de los cuartones.
La dinámica de las L3 en el pasto coincide, en parte, con la obtenida por Altaif y Jacobs (1987), Aumont et al. (1996) y Fonseca et al. (2004) en Iraq, Guadalupe y Cuba, respectivamente. Estos autores hallaron los mayores niveles de infestación de los pastos en el PLL; dicho comportamiento fue atribuido a los efectos positivos de las precipitaciones, la temperatura y la radiación solar en el proceso de desarrollo y sobrevivencia de las larvas en el pasto. Por otra parte, se ha demostrado que las L3 necesitan de una pequeña película de agua para trasladarse por el pasto; en el PPLL esta es mínima y depende, en gran medida, de la humedad condensada en la noche (Cordero et al., 1998). Estos resultados difieren de los obtenidos por Amarante y Barbosa (1995) en Brasil, quienes en condiciones climáticas similares encontraron en el período de máximas precipitaciones una menor cantidad de larvas infestantes; ellos consideran que este comportamiento se debe al «efecto de lavado» de las precipitaciones y al «efecto dilución» de las poblaciones larvarias debido al crecimiento del pasto, situación que no se constató en el estudio aquí realizado.
La relación positiva de las precipitaciones con el desarrollo y la sobrevivencia de las poblaciones de las L3 en el pasto en el presente estudio, coincide con los trabajos realizados por Aumont y Gruner (1989), Rossanigo y Gruner (1994) y García-Romero et al. (1997); sin embargo, también se han reportado relaciones no bien definidas entre estas dos variables (Simón et al., 1996). Los resultados demostraron que las precipitaciones fueron la principal variable meteorológica que influyó en la dinámica de las poblaciones larvarias, en las condiciones del estudio.
La temperatura media influyó de forma positiva en la cantidad de larvas recuperadas en Dos Mercedes y la EEPF «Indio Hatuey»; sin embargo, este comportamiento no quedó bien definido en LABIOFAM. En esta última finca tampoco se encontraron relaciones definidas entre las L3 y la radiación solar global y la humedad relativa. Ello pudiera explicarse por las características de las áreas de pastoreo, las que poseen pastos naturales de baja talla que sólo alcanzan una altura de 3-8 cm aproximadamente y apenas les brindan protección a esos estadios infestivos; esto pudiera limitar la migración de las larvas, las cuales tienden a evitar las temperaturas elevadas y la radiación solar, cuando no encuentran la protección necesaria en la vegetación del entorno.
Los resultados hallados en Dos Mercedes y en la EEPF «Indio Hatuey» acerca de la relación entre las temperaturas y las poblaciones larvarias coinciden con los informados por Rossanigo y Gruner (1994), quienes apreciaron en un análisis de componentes principales que la temperatura media y la mínima, así como el contenido de agua de las heces, explicaban el 54% de la varianza total para el género Haemonchus; sin embargo, difieren de los reportados por García-Romero et al. (1997), quienes encontraron relaciones negativas entre las dos variables analizadas.
La radiación solar global presentó coeficientes medios en la EEPF «Indio Hatuey» (r=0,45 para la cantidad de larvas de Haemonchus spp. y r=0,44 para el total de larvas), mientras que en las otras dos unidades no se apreció un comportamiento definido. Estos resultados difieren de los obtenidos por Simón et al. (1996), quienes encontraron coeficientes de correlación entre -0,21 y -0,40 de acuerdo con las especies y el período de muestreo; ello se atribuyó al tipo de pasto y al horario de muestreo, que generalmente se desarrolló en horas de la mañana (8:00-9:00 a.m.), momento del día en que las radiaciones son débiles y se favorece la migración de las L3 hacia las capas superiores del pasto como producto del fototropismo positivo a la luz tenue (Cuéllar, 2002).
En un estudio epizootiológico resulta de gran interés conocer el origen del aumento de los niveles de las L3 en el pasto, como elemento para la concepción de las estrategias destinadas a minimizar el enfrentamiento entre los animales susceptibles y las poblaciones de las larvas infestantes. En las tres unidades dicho aumento fue estacional, lo cual se debió al rápido desarrollo de los huevos recién expulsados al pasto, que al encontrar condiciones ambientales favorables en el PLL devinieron en larvas infestantes. Las implicaciones epizootiológicas de estos picos adquieren una trascendencia considerable, ya que Haemonchus predomina en los rebaños ovinos (Barger, 1999).
Estos resultados revisten una gran importancia, ya que la tasa de infestación del pasto o la cantidad de larvas ingeridas cada día es la variable epizootiológica de mayor influencia en la carga parasitaria de los animales en pastoreo. En este sentido, el conocimiento de los niveles de contaminación de los pastos por las larvas infestantes de los estrongílidos resulta de interés para los planes estratégicos de control parasitario. Por otra parte, es importante señalar que las tendencias actuales no solo tienen en cuenta los niveles de infestación parasitaria del rebaño, sino también la población larvaria en el pasto, denominada «población en refugio» (Van Wyk, 2001), con el objetivo de trazar estrategias de aplicación de los antihelmínticos y así minimizar la presión de selección de individuos resistentes, para el establecimiento y desarrollo de planes de lucha mediante el uso de antiparasitarios de manera efectiva.
REFERENCIAS BIBLIOGRÁFICAS
1. Altaif, K.I. & Jacobs, A.Y. Development and survival of Haemonchus contortus larvae on pasture in Iraq. Tropical Animal Production. 19:88. 1987
2. Amarante, A.F.T. & Barbosa, Maria A. Seasonal variations in populations of infective larvae on pasture and nematode faecal output in sheep. Veterinária e Zootecnia. 7:127. 1995
3. Arece-García, J. et al. The epizootiology of ovine gastrointestinal strongyles in the province of Matanzas, Cuba. Small Ruminant Research. 72:119. 2007
4. Arece, J. & Rodríguez, J. El parasitismo gastrointestinal del ganado ovino en Cuba. Revista ACPA. 4:50. 2003
5. Aumont, G. et al. The epidemiology of intestinal parasitism in creole goats in Guadeloupe (French West Indies). Annals of the New York Academy of Sciences. 791:738. 1996
6. Aumont, G. & Gruner, L. Population evolution of the free-living stage of goat gastrointestinal nematodes on herbage under tropical conditions in Guadeloupe (French West Indies). International Journal for Parasitology. 19:539. 1989
7. Barger, I. The role of epidemiological knowledge and grazing management for helminth control in small ruminants. International Journal for Parasitology. 29:41. 1999
8. Cordero, M. et al. Parasitología veterinaria. Mc Graw Hill Interamericana, México. 934 p. 1998
9. Cuéllar, J.A. Agentes etiológicos de la nematodiasis gastrointestinal en los diversos ecosistemas. En: Epidemiología y control integral de nemátodos gastrointestinales de importancia económica en pequeños rumiantes. (Eds. J.F. Torres-Acosta y A.J. Aguilar). Segundo curso internacional. FMVZ-Universidad Autónoma de Yucatán. México. p. 1. 2002
10. Delgado, A. Supervivencia de las larvas de Haemonchus spp. en el medio subtropical de Cuba. Revista Cubana de Ciencias Veterinarias. 14 (1):49. 1983
11. Fonseca, N. et al. Alternativas alimentarias para ovinos y caprinos más usadas en las zonas orientales de Cuba. [cd-rom]. Memorias del Congreso Internacional de Agricultura en Ecosistemas Frágiles y Degradados. IIA «Jorge Dimitrov». Granma, Cuba. 2004
12. García, Amelia et al. Supervivencia larvaria de strongylata de ovinos en pasto natural. Revista de Producción Animal. 8 (2):154. 1994
13. García-Romero, C. et al. Influence of climate on pasture infectivity of ovine trichostrongyles in dry pastures. Journal of Veterinary Medicine. 44:437. 1997
14. García-Romero, C. et al. Diagnóstico antermortem: análisis coprológico, de la hierba y hemático. Ovies. 70:23. 2000
15. González, R. Alternativas de control de nemátodos gastrointestinales en ovinos. Conferencia. EEPF «Indio Hatuey». Matanzas, Cuba. 2004
16. Gruner, L. et al. Dynamique des populations des parasites internes dans un élevage semi-intensif de chèvres créoles en Guadeloupe. Les Colloques de l'INRA. 28:695. 1984
17. Gruner, L. & Raynaud, J.P. Technique allégée de prélèvements d'herbe et de numération pour juger de l'infestation des pâturages de bovins par les larves de nématodes parasites. Revue de Médicine Vétérinaire. 131:521. 1980
18. Rossanigo, C.E. & Gruner, L. Relative effect of temperature and moisture on the development of strongyle eggs to infective larvae in bovine pats in Argentina. Veterinary Parasitology. 55:317. 1994
19. Simón, R. et al. Herbage density of third-stage larvae of goat strongyles during the dry season in Guadeloupe. Annals of the New York Academy of Sciences. 791:412. 1996
20. Sissay, M.M. et al. Anthelmintic resistance of nematode parasites of small ruminants in eastern Ethiopia: Exploitation of refugia to restore anthelmintic efficacy. Veterinary Parasitology. 135:337. 2006
21. Smothers, C.D. et al. Comparison of arithmetic and geometric means as measures of central tendency in cattle nematode population. Veterinary Parasitology. 81:211. 1999
22. Thamsborg, S.M. et al. The influence of stocking rate on gastrointestinal nematode infections of sheep over a 2-year grazing period. Veterinary Parasitology. 67:207. 1996
23. Valle, María T. Contribución al estudio de los nemátodos gastrointestinales del ganado bovino. Tesis presentada en opción al grado de Doctor en Ciencias Biológicas. Universidad de La Habana. La Habana, Cuba. 150 p. 1978
24. Van Wyk, J.A. Refugia-overlooked as perhaps the most potent factor concerning the development of anthelmintic resistance. Onderstepoort Journal of Veterinary Research. 66:55. 2001
Recibido el 28 de octubre de 2009
Aceptado el 4 de diciembre de 2009
