Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.33 n.2 Matanzas abr.-jun. 2010
ARTÍCULO DE INVESTIGACIÓN
Diversidad de la flora en fincas ganaderas de la provincia de Matanzas
Flora diversity in livestock production farms of Matanzas province
R. Machado1, Taymer Miranda1 y J.L. Álvarez2
1Estación Experimental de Pastos y Forrajes «Indio Hatuey»
Central España Republicana CP 44280, Matanzas, Cuba.
E-mail: rmachado@indio.atenas.inf.cu
2Universidad de Matanzas «Camilo Cienfuegos», Matanzas, Cuba
RESUMEN
El objetivo de este trabajo fue determinar y cuantificar las especies que forman parte de la diversidad de la flora en ocho fincas ganaderas o integrales, vinculadas al Centro Local de Innovación Agropecuaria de Matanzas. Para ello se utilizó un diseño muestral sistemático; el tamaño de la muestra no fue inferior a 0,5%. En general, existió una alta diversidad para todas las fincas. En tres de las ocho fincas monitoreadas se encontraron 30 o más especies; en otras tres este número fluctuó entre 19 y 26; mientras que en las dos restantes se contabilizaron 13 y 15 especies, respectivamente. Ello se correspondió con los índices de riqueza, que estuvieron entre 2,03 y 5,22. Existió superioridad numérica de las gramíneas y de las leguminosas sobre las especies de otras familias, cuyo número fluctuó entre una y seis especies (cuatro o menos en el 75% de las fincas). Los mayores valores de dispersión se encontraron en las gramíneas, cuyos índices fueron superiores al 60% y de más del 80% para dos de las fincas. Se sugiere hacer un análisis casuístico, en cada finca, para determinar las futuras estrategias, incluyendo la erradicación parcial y mejora de la cubierta vegetal, o su erradicación y retransformación total, variantes que decidirán los productores respaldados por un asesoramiento.
Palabras clave: Biodiversidad, pastizal natural
ABSTRACT
The objective of this work was to determine and quantify the species that are part of the flora diversity in eight livestock production or integral farms, linked to the Local Agricultural Innovation Center of Matanzas. For that purpose, a systematic sampling design was used; the sample size was not lower than 0,5%. In general, there was high diversity for all farms. In three of the eight monitored farms 30 or more species were found; in other three this number fluctuated between 19 and 26; while in the other two 13 and 15 species were counted, respectively. This was in correspondence with the richness values, which were between 2,03 and 5,22. There was numeric superiority of grasses and legumes over the species from other families, which number oscillated between one and six species (four or less in 75% of the farms). The highest dispersion values were found in the grasses, higher than 60% and more than 80% for two farms. The performance of a casuistic analysis, in each farm, is suggested in order to determine the future strategies, including the partial eradication and improvement of the plant cover, or its eradication and total retransformation, variants that will be decided by the farmers supported by advice.
Key words: Biodiversity, natural pastureland
INTRODUCCIÓN
En los pastizales naturales o establecidos, la diversidad de plantas superiores en la cubierta vegetal o la composición botánica, como se le conoce en el ambiente agropecuario, generalmente está representada por especies pertenecientes a una o más familias. Cuando esta diversidad se estima de forma conveniente con el uso de métodos apropiados, se obtienen indicadores de relevante importancia en la valoración de su estructura y naturaleza, toda vez que estos permiten comprobar la cuantía, la especificidad y otros importantes aspectos relacionados con el valor de sus componentes.
Por otra parte, estos indicadores ayudan a interpretar la influencia que ha tenido el ambiente, incluido el manejo, a partir del estatus existente. Con ello se posibilita la toma de decisiones para el diseño de alternativas de retransformación de esas áreas, de forma parcial o total, o bien el mantenimiento de la composición.
El monitoreo de la composición florística ha sido frecuentemente utilizado en experimentos de tipo agronómico o de perfil puramente pecuario, con el fin de comprobar los cambios que ocurren en la cubierta vegetal por efecto de la aplicación de uno o varios tratamientos, y también para el desarrollo de censos poblacionales en entidades productivas, con el objetivo de establecer programas de ajuste en el fomento o recuperación de los pastizales (MINAGRI, 1991; Anon, 2000).
El objetivo de este trabajo fue monitorear la diversidad florística en ocho fincas ganaderas vinculadas al Centro Local de Innovación Agropecuaria (CLIA) de la provincia de Matanzas, con el fin de obtener información acerca de la naturaleza y la cuantía de las especies existentes, previamente a la toma de decisiones de retransformación de estas áreas.
MATERIALES Y MÉTODOS
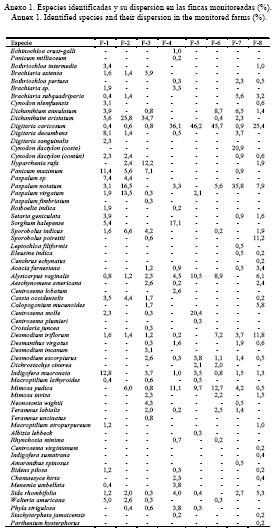
Características de las fincas y los suelos. Las fincas pertenecen a productores líderes integrados a varias cooperativas de créditos y servicios de la provincia Matanzas. Estas se caracterizan por poseer áreas relativamente pequeñas y, según el propósito (integrales o ganaderas), los pastizales se explotan en condiciones de secano y están formados por especies naturalizadas; en algunos casos se encuentran pobladas por especies de gramíneas cultivadas que se establecieron en etapas anteriores. Ninguna de estas áreas se fertiliza; los animales dependen fundamentalmente del pasto para la producción de leche o carne, y la mayoría reciben cantidades limitadas de suplemento (tabla 1).

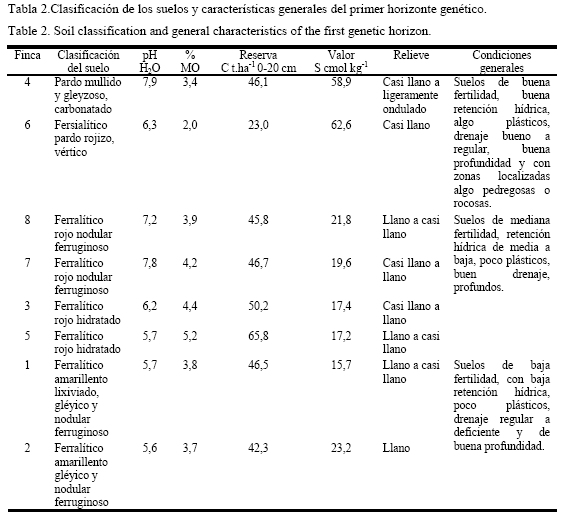
Para determinar los tipos de suelos, en correspondencia con la clasificación genética de uso actual (Hernández et al., 1999), se cavaron calicatas de más de un metro de profundidad en puntos representativos del área, coincidentes aproximadamente con el centro de cada una de las fincas. Además se tomaron entre seis y ocho muestras de suelo alrededor de la calicata y en puntos dispersos a través de transeptos diagonales, a una profundidad de 15 cm. En la determinación del pH y el contenido de MO se utilizaron los métodos potenciométrico y Walkley Black, respectivamente; mientras que el valor de S se calculó sobre la base de la suma de los principales cationes retenidos en el suelo: Ca++, Mg++, K+ y Na+.
Seis de las fincas están ubicadas en suelos del agrupamiento Ferralítico, caracterizado por presentar un pH de moderadamente ácido a neutro, con bajos valores de S, y medianamente abastecido de MO (3,7 - 5,2%). Las dos restantes ocupan suelos de los agrupamientos Pardo y Fersialítico, químicamente más fértiles, pero con algunos problemas asociados al drenaje. Solo en dos de las fincas en suelo Ferralítico, se comprobó la existencia de drenaje deficiente por la presencia de horizontes gléyicos (fincas 1 y 2). De acuerdo con estas características, las fincas se distribuyeron en tres grupos (tabla 2).
PROCEDIMIENTO EXPERIMENTAL
Muestreo. Con el fin de identificar las especies y su cuantía, se llevó a cabo un sistema muestral sistemático a través de transeptos en diagonales y paralelos.
Se eligió un tamaño de muestra nunca inferior a 0,5%, lo que equivale al conteo de las especies en no menos de 200 puntos por hectárea, al considerar que en cada punto se determinaron todas las especies existentes en un área equivalente a 0,25 m2.
Análisis de los resultados
La interpretación de los resultados se hizo a través de un sistema descriptivo para todas las especies y su ubicación taxonómica por familia para las gramíneas y las leguminosas, no así para las restantes que se identificaron, en general, como especies de otras familias.
Además, se determinaron los índices de dispersión y de riqueza. Para el primero se sumaron las observaciones de todas las especies (cifra que se asumió como 100%) y por simple proporción se determinó el porcentaje de cada especie en particular; a continuación se sumaron todos los porcentajes para cada familia. Para el segundo índice también se utilizó la
suma total de observaciones, el número de especies y el número de veces que apareció cada una de estas. Los datos se procesaron a través del programa DIVERSI para ambiente Windows, que permite obtener el índice de riqueza y otros índices de interés.
RESULTADOS Y DISCUSIÓN
La diversidad de especies, su abundancia y la composición de la comunidad en los agroecosistemas, se considera como un producto de las interacciones bióticas interespecíficas (cuya base radica en un fuerte efecto de competencia y/o depredación en la estructura de la comunidad, por ejemplo la competencia que se establece con los herbívoros entre plantas), o como el efecto que producen los eventos de dispersión y las limitaciones de la dispersión local. Incluso, Tilman (1997) plantea que es un producto de ambos efectos.
Teniendo en cuenta este último enfoque como el más apropiado, lo más importante radica en reconsiderar, en el orden práctico, la cuantía y la naturaleza de las especies que se encuentran en esos agroecosistemas y analizar la calidad de la composición del material, lo cual permitirá alcanzar una visión realista para la propuesta de estrategias de cambio, mantenimiento o mejora de esa composición.
Los resultados mostraron que existió una alta diversidad en todas las fincas monitoreadas (tabla 3). De esta forma, los pastizales en tres de las fincas estuvieron conformados por 30 o más especies botánicas. Este número fluctuó entre 19 y 26 en otras tres; mientras que en las dos restantes se contabilizaron 13 y 15 especies, respectivamente. Ello se correspondió con los índices de riqueza.

De acuerdo con lo propuesto por Christensen (1995) esta diversidad responde, en primer lugar, a la madurez que han alcanzado estos agroecosistemas, ya que se trata de pastizales con muchos años de existencia; en segundo lugar, a la presencia de los animales casi de forma permanente (fundamentalmente bovinos), los cuales ejercen un papel importante en la diversidad de las especies, debido a los efectos directos e indirectos que tienen sobre la cubierta vegetal, según la revisión realizada por Machado y Olivera (2003); y, por último, al efecto que puede causar el clima y otros factores en la riqueza de especies (Jones et al., 1995).
Por otra parte, el número de especies, además de ser alto, también fue considerablemente variable. Ello indica que la composición de las especies en términos numéricos puede ser diferente entre los agroecosistemas de pastizales naturales y más aún en los pastizales comerciales. Machado (2002) detectó 39 especies en un pastizal establecido con una variedad cultivada cinco años antes del monitoreo y manejado indistintamente con el aporte de fertilización nitrogenada durante la época de lluvia (tres años) o sin fertilización (dos años); mientras que Jones et al. (1991) encontraron 57 especies diferentes que crecieron, a partir de las reservas de semillas en el suelo, en pastizales asociados o gramíneas fertilizadas en un período experimental de 11 años, lo que reafirma tal criterio.
El número de especies es un indicador importante, al igual que la naturaleza de las especies que componen la cubierta vegetal, lo que obedece a la mayor o menor presencia de plantas con un determinado valor para la producción animal. En este sentido es válido resaltar la menor cuantía en que se encontraron las especies de otras familias, las cuales en sentido general mostraron un menor valor y son menos consumidas que las gramíneas y las leguminosas, con algunas excepciones. Para estas su número fluctuó entre una y seis (cuatro o menos en el 75% de las fincas).
La mayor presencia de las gramíneas y las leguminosas, con un valor más alto, tiene su explicación en varios atributos morfoestructurales y fisiológicos conocidos, tales como: un mayor número de puntos de crecimiento; reservas suficientes para desarrollar su potencialidad de rebrote después de la defoliación o de los daños producidos por estrés; estolones o rizomas (o ambos) que le confieren un alto poder de colonización en la mayoría de los casos y una alta producción de semilla, lo que asegura su rápida propagación en el área; en dichos aspectos superan a las especies de otras familias, que se encontraron siempre en menor número.
Los índices de riqueza se manifestaron indistintamente con valores altos o bajos. Así, en las fincas 4 y 6 (grupo 1) fueron de 3,93 y 2,13 (de medios a bajos); estos son los suelos de mayor fertilidad y retención hídrica, en los que normalmente se adapta una amplia gama de especies vegetales. Los valores de las fincas del grupo 2 ( 3, 5, 7 y 8) fluctuaron entre 4,1 y 5,1 para tres de ellas (aceptables), a pesar de que los suelos poseen fertilidad media, y sólo fueron de 2,03 en la finca 5; mientras que en las fincas del grupo 3, donde predominan los suelos de más baja fertilidad y con condiciones físicas limitantes para el desarrollo de diversas especies vegetales (fincas 1 y 2), fueron de 5,2 (alto) y 2,9 (bajo), respectivamente. Ello indica que el índice de riqueza no parece tener mucha relación, en estos casos, con el tipo, la calidad y la capacidad productiva de sus suelos, sino que puede obedecer a aspectos relacionados con la capacidad de adaptación que poseen las especies para colonizar un agroecosistema determinado, independientemente de la fertilidad, lo que confirma lo planteado por Tilman (1997).
Aunque se encontraron especies de leguminosas en estos agroecosistemas (tablas 2 y 3; anexo 1), independientemente del grado de fertilidad de los suelos y de sus propiedades físicas, el menor número se halló en las fincas ubicadas sobre los suelos del grupo 3, es decir, los de fertilidad más baja, con una baja retención hídrica y un drenaje de regular a deficiente, aunque de buena profundidad. En cuatro de las fincas enclavadas en suelos de los grupos 1 y 2, donde las condiciones edáficas son mejores, el número de leguminosas fue superior al de gramíneas, en otra lo igualó y solo en una de ellas (finca 7) fue inferior. Estas especies, mucho menos eficientes que las gramíneas desde el punto de vista fisiológico y en general más limitadas para adaptarse a ambientes agresivos, tienden a aumentar en número y área cuando las condiciones les son más favorables o propicias para su desarrollo, ya que pueden sobrevivir y lograr una tasa de crecimiento determinado en suelos de mediana a alta fertilidad (Thornley et al., 1995), y además muestran preferencia para proliferar en suelos bien drenados (Álvarez, 2002), sobre todo cuando no se utilizan fertilizantes inorgánicos y otros agrotóxicos (Abdalla et al., 2000).
Los valores más altos en términos de dispersión para todas las fincas, excepto en una, se detectaron en las gramíneas (tabla 4), con índices superiores al 60%, y más del 80% para dos de ellas. En la finca donde las leguminosas superaron en dispersión a las gramíneas (finca 5), el porcentaje fue superior solo en tres unidades, lo que denota más bien un estatus de equilibrio que una superioridad absoluta.
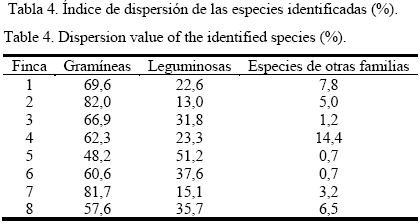
No obstante, los índices de dispersión de las leguminosas se consideran adecuados, particularmente en las fincas 3, 6 y 8, además de la 5, y solo estuvo por debajo del 15% en la finca 2, en correspondencia con sus características (tabla 2). La combinación de las leguminosas con las gramíneas de diversos hábitos en los pastizales, se atribuye al alto grado de compatibilidad que caracteriza a muchas de sus especies, sobre todo cuando encuentran ambientes adecuados para su desarrollo y proliferación. La presencia de estas especies, con un alto grado de dispersión en los sistemas, fue detallada por Jardines (2006) al estudiar la flora preponderante en pastizales naturales sometidos o no a pastoreo.
Es importante señalar que prácticamente todas las gramíneas naturalizadas en estos pastizales, e incluso muchas de las leguminosas, se caracterizan por una marcada agresividad y un amplio margen de adaptación en esos lugares, donde han creado sus bancos de semilla, y a la vez han contribuido al mantenimiento de un cierto nivel de producción en la masa ganadera que en ellos pastorea. Ello alerta sobre la necesidad de llevar a cabo transformaciones en estas fincas que prevean la introducción de especies o variedades comerciales de gramíneas y de leguminosas, incluyendo entre estas últimas tipos herbáceos o arbóreos con posibilidades de adaptación, pero que a la vez posibiliten incrementar el potencial de producción de los animales sobre la base de una mayor calidad y un mayor consumo.
Los índices de dispersión en las especies de otras familias fueron bajos. Estas no constituyen un peligro de convertirse en invasoras de grandes áreas compactas, ya que normalmente se distribuyen de forma individual o crean pequeñas poblaciones de individuos, lo cual se puede observar en la mayoría de los pastizales naturales.
Los resultados se consideran valiosos, ya que permiten obtener una visión real de la composición de la flora en fincas en las que se pretende acometer transformaciones. Ello presupone hacer un análisis casuístico, en cada una, para determinar las futuras estrategias que se aplicarán, las que podrían incluir: erradicarla parcialmente y mejorar esas áreas (en el caso de las fincas 3, 5, 6 y 8) o erradicarla totalmente y transformar las áreas (fincas 1, 2 y 7), en las que predominan gramíneas de muy baja calidad y pocas especies de leguminosas con baja dispersión; incluso en la finca 4, donde hay un mayor número de leguminosas, tampoco alcanzan un aceptable grado de dispersión en relación con las gramíneas. No obstante, se sugiere que cualquiera de las alternativas que se aplique sea una decisión de los productores, respaldada por un asesoramiento técnico.
REFERENCIAS BIBLIOGRÁFICAS
1. Abdalla, M.H. et al. The impact of pesticides on arbuscular mycorrizal and nitrogen fixing symbiosis in legumes. Applied Soil Ecology. 14:191. 2000
2. Álvarez, Orquidia. Diversidad de leguminosas y su potencial productivo sobre suelos Pardos con Carbonatos de la zona centro de Sancti Spiritus. Tesis en opción al título de Maestro en Ciencias en Pastos y Forrajes. Estación Experimental de Pastos y Forrajes «Indio Hatuey». Matanzas, Cuba. 69 p. 2002
3. Anon. Recuperación de pastizales. Vías y estrategias para Cuba. Taller 35 Aniversario del Instituto de Ciencia Animal (ICA)-Departamento de Pastos. La Habana, Cuba. 2000
4. Christensen, V. Ecosystem maturity- towards quantification. Ecological Modeling. 77:3. 1995
5. Hernández, A. et al. Nueva versión de clasificación genética de los suelos de Cuba. Instituto de Suelos. La Habana, Cuba. 64 p. 1999
6. Jardines, Sonia. Caracterización ambiental de los pastizales naturales de Cuba. Trabajo de Investigación. Universitat de Girona. Institut de Medi Ambient. Doctorat de Cooperació «Gestión Ambiental y Desarrollo Sostenible». 204 p. 2006
7. Jones, R.M. et al. Levels of germinable seed in topsoil and cattle faeces in legume-grass and nitrogen-fertilizer pasture in South-East Queensland. Aust. J. Agric. Res. 42:953. 1991
8. Jones, R.M. et al. Some advantages of long-term grazing trials, with particular reference to changes in botanical composition. Aust. of Exp. Agric. 35:9. 1995
9. Machado, R. Variaciones morfoestructurales y poblacionales de Andropogon gayanus Kunth y su relación con la vegetación adventicia bajo pastoreo intensivo. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas. Universidad de Matanzas «Camilo Cienfuegos»-Estación Experimental de Pastos y Forrajes «Indio Hatuey». Matanzas, Cuba. 116 p. 2002
10. Machado, R. & Olivera, Yuseika. Bases conceptuales e implicaciones de las variaciones de la vegetación en los pastizales. Pastos y Forrajes. 26:279. 2003
11. MINAGRI. Resultados del Inventario Nacional de Pastos al final de 1991. La Habana, Cuba. (Mimeo). 1991
12. Thornley, J.H.M. et al. Complex dinamics in a carbon-nitrogen model of a grass-legume pasture. Annals of Botany. 75:79. 1995
13. Tilman, D. Community invasibility, recruitment limitation and grassland biodiversity. Ecology. 78 (1):81. 1997
Recibido el 14 de octubre del 2009
Aceptado el 20 de diciembre del 2009