










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.34 n.1 Matanzas ene.-mar. 2011
ARTÍCULO DE INVESTIGACIÓN
Capacidad de recuperación de 23 accesiones de Leucaena spp. después de la poda
Recovery capacity of 23 Leucaena spp. accessions after pruning
Hilda B. Wencomo1 y R. Ortiz2
1Estación Experimental de Pastos y Forrajes «Indio Hatuey»Central España Republicana, CP 44280, Matanzas, Cuba.
E-mail: hilda.wecomo@indio.atenas.inf.cu
2Instituto Nacional de Ciencias Agrícolas, La Habana, Cuba
RESUMEN
Se evaluaron 23 accesiones de Leucaena spp. durante dos años, con el objetivo de conocer la variabilidad existente en la población en cuanto a la capacidad de recuperación después de la poda. Cuando las plantas sobrepasaban los 2 m, se realizó la poda de uniformidad a un 1 m de altura sobre el nivel del suelo. Las mediciones fueron: el grosor del tallo, el número de rebrotes y su longitud con una frecuencia semanal, a partir de lo cual se calculó la velocidad de crecimiento de cada accesión; además se determinó el rendimiento. El análisis de componentes principales, el de conglomerados y el de regresión lineal simple se realizaron a través del programa estadístico SPSS versión 11.5 para Microsoft Windows®. Se observó variabilidad en los indicadores evaluados (86,11% acumulado en la componente) y todos contribuyeron a su explicación. A su vez, el análisis de conglomerados permitió la formación de tres grupos. Se concluye que todas las accesiones presentaron capacidad de recuperación después de la poda, con diferencias entre ellas. Las accesiones Leucaena leucocephala cv. Cunningham, cv. Perú, CIAT-9119, CIAT-9438, CIAT-751, CIAT-7988, CIAT-7384, CIAT-7929, CIAT-17480, cv. Ipil-Ipil y cv. CNIA-250; L. lanceolata CIAT-17255 y CIAT-17501 y L. diversifolia CIAT-17270 fueron las de mayor capacidad de recuperación. Por ello se recomienda, en futuras investigaciones relacionadas con la caracterización, evaluación y selección de estas especies, realizar estudios que incluyan su persistencia en el tiempo bajo condiciones de poda, así como su empleo en el fomento de nuevos sistemas silvopastoriles.
Palabras clave: Comportamiento, Leucaena spp., poda.
ABSTRACT
Twenty three Leucaena spp. accessions were evaluated for two years, in order to know the variability existing in the population regarding the recovery capacity after pruning. When the plants were higher than 2 m, the uniformity pruning was made at 1 m above the soil level. The measurements were: stem diameter, number of regrowths and their length with a weekly frequency, from which the growth rate of each accession was calculated; in addition, yield was determined. The main component, cluster and simple lineal regression analyses were made through the statistical program SPSS version 11.5 for Microsoft Windows®. Variability was observed in the evaluated indicators (86,11% accumulated in the component) and they all contributed to its explanation. In turn, the cluster analysis allowed the formation of three groups. All the accessions were concluded to show recovery capacity after pruning, with differences among them. The accessions Leucaena leucocephala cv. Cunningham, cv. Peru, CIAT-9119, CIAT-9438, CIAT-751, CIAT-7988, CIAT-7384, CIAT-7929, CIAT-17480, cv. Ipil-Ipil and cv. CNIA-250; L. lanceolata CIAT-17255 and CIAT-17501 and L. diversifolia CIAT-17270 were the ones with higher recovery capacity. For such reason it is recommended, in future studies related to the characterization, evaluation and selection of these species, to make studies including their persistence in time under pruning conditions, as well as their use in the development of new silvopastoral systems.
Key words: Performance, Leucaena spp., pruning.
INTRODUCCIÓN
La fisiología de la defoliación y el rebrote constituyen asuntos medulares de la biología de las plantas, ya que son elementos básicos del manejo de los pastizales. En el orden práctico se busca respuesta a la influencia de la altura y la frecuencia de defoliación (corte o pastoreo) en la dinámica de crecimiento y en el rendimiento durante el período de rebrote. La biomasa aérea de los árboles y los arbustos presentes en los sistemas silvopastoriles proporciona una parte de los alimentos a los animales que allí se desarrollan, por lo que el conocimiento adecuado de su manejo es fundamental para lograr un mayor nivel y estabilidad en la producción de biomasa a través de todo el año.
La especie Leucaena leucocephala es una de las más estudiadas y utilizadas en los sistemas silvopastoriles (Hernández et al., 2000), lo cual está avalado por su alto valor proteínico, entre otros aspectos; además se utiliza como fuente de sombra (Molina et al., 2001), para conservar y mejorar el suelo, producir grandes cantidades de biomasa y reciclar los nutrientes (Sánchez, 2007); tiene la capacidad de rebrotar después de ramoneada o cosechada de forma mecánica y de restablecerse rápidamente del estrés biótico o abiótico (Ruiz y Febles, 2001), aspecto muy importante a tener en cuenta cuando se emplea en dichos sistemas. A pesar de ello, el comportamiento entre las accesiones es diferente.
Atendiendo a las características mencionadas se inició un estudio de evaluación de 23 accesiones pertenecientes a una colección del género Leucaena, con el objetivo de conocer la variabilidad existente en la población en cuanto al comportamiento de la capacidad de recuperación después de la poda.
MATERIALES Y MÉTODOS
Ubicación del área experimental. El estudio se realizó en las áreas de la Estación Experimental de Pastos y Forrajes Indio Hatuey , la cual se encuentra ubicada en los 22º 48' y 7'' de latitud Norte y los 79º 32' y 2'' de longitud Oeste, a una altitud de 19,9 msnm, en el municipio de Perico, provincia de Matanzas, Cuba (Academia de Ciencias de Cuba, 1989).
Características del clima. La suma promedio de la precipitación de los dos años fue de 2 462,3 mm. La lluvia caída durante la estación lluviosa (mayo-octubre) representó, como promedio, el 79,8% del volumen total anual. La evaporación en la zona aumentó a partir de enero, con valores máximos en abril (220 mm). La humedad relativa promedio anual fue de 82,6%, con el mayor valor en julio (89,0%) y el menor en abril (75,5%).
Características del suelo. El suelo es de topografía llana, con pendiente de 0,5 a 1,0%, y está clasificado por Hernández et al. (2003) como Ferralítico Rojo lixiviado, húmico nodular ferruginoso hidratado, de rápida desecación, arcilloso y profundo sobre calizas. Este tipo es equivalente al grupo de los Ferrosoles, en el sistema de clasificación de la FAO-UNESCO (Alonso, 2003).
Procedimiento experimental
Material vegetal. De las 180 accesiones establecidas en la colección de Leucaena spp. que se conservan en el banco de germoplasma de la Estación, se tomaron 23 (tabla 1) representativas de la población (de cada una de ellas se evaluaron cuatro plantas durante dos años).
La poda se efectúo en el período poco lluvioso, específicamente en el mes de noviembre de cada año según lo propuesto por Hernández (2000). Cuando las plantas sobrepasaban los 2 m de altura (entre 3 y 4 m) se realizó la poda a un 1 m sobre el nivel del suelo, de acuerdo con lo recomendado por Francisco et al. (1998).
Las mediciones relacionadas con la capacidad de recuperación de las accesiones fueron: el número de ramas, el grosor del tallo (en la base, con un pie de rey) y el número de rebrotes. Para esta última se realizó el conteo de los rebrotes emitidos por las plantas y se midió su longitud (con una regla graduada), con una frecuencia semanal.
Este procedimiento se hizo en los cinco rebrotes más desarrollados (Toral et al., 2006) hasta que las plantas podadas alcanzaron los 2 m o más de longitud y se determinó además el rendimiento.
Procesamiento estadístico. Los datos se procesaron mediante el análisis de componentes principales (ACP) (Morrison, 1967), en el cual se tomó como criterio de análisis aquellas componentes principales que presentaron valores propios superiores a uno y factores de suma o de preponderancia mayor que 0,70.
Posteriormente se aplicó el análisis de conglomerados para la agrupación y selección de las accesiones, utilizando como criterio la distancia euclidiana, a partir de lo obtenido en el ACP (Torres et al., 2006), y se determinaron los estadígrafos media y desviación estándar para las variables analizadas.
Se utilizó además el análisis de regresión lineal para conocer los modelos de mejor ajuste y la relación funcional entre las variables número de rebrotes y su longitud. Este análisis identifica el modelo o función que une las variables, estima sus parámetros y, eventualmente, prueba la hipótesis acerca de ellos. Una vez estimado el modelo es posible predecir el valor de la variable dependiente en función de la (s) otra (s) variable (s) independiente (s), y dar una medida de la precisión de esa estimación. Se consideró como variable independiente los días y como variable dependiente, el número de rebrotes y su longitud.
Como norma de selección de la ecuación de mejor ajuste se tomaron en consideración los criterios de Guerra et al. (2003): nivel de significación, coeficiente de determinación, R2 mayor que 0,70, varianza residual V(e), análisis de residuos (e1) y el error estándar de los parámetros estimados ES (âi). Todos los análisis se realizaron a través del programa estadístico SPSS versión 11.5 para Microsoft Windows® (Visuata, 1998).
Selección de las accesiones. En el proceso de selección se evaluó el comportamiento de cada accesión en las variables que más contribuyeron en la formación de los componentes (tabla 2) y se tomó en consideración algunas de las características deseables (en este caso agronómicas) que deben reunir los árboles y arbustos utilizados en los sistemas silvopastoriles, según criterios de Borel (1997), Gómez et al. (2002) y Simón et al. (2005). Se estableció que las accesiones seleccionadas debían cumplir con todos o con dos de los criterios evaluativos considerados.

RESULTADOS Y DISCUSIÓN
Al realizar el análisis de componentes principales se obtuvo una sola componente, la cual explicó el 86,11% de la varianza total (tabla 3); a ello contribuyeron todos los indicadores evaluados.
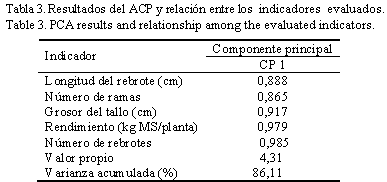
La variabilidad mostrada a través de los indicadores puede que se deba a la alta relación que existió entre estos, o a la dinámica de crecimiento de cada accesión en particular (Dávila y Urbano, 1996), lo cual puede estar relacionado con la reserva acumulada por las plantas y con la capacidad de utilización, y es expresión de la potencialidad genética particular de las especies de Leucaena. Dicho aspecto es de sumo interés para los sistemas silvopastoriles, ya que la capacidad para rebrotar, así como su adecuado desarrollo y rápido crecimiento, determinan su utilización y pueden contribuir como indicadores de suma importancia a la selección de especies y accesiones forrajeras.
Es importante enfatizar lo señalado por Seguí et al. (1989), quienes plantearon que el conocimiento de las relaciones entre los caracteres de interés agronómico desempeña un papel significativo en el proceso de selección, e indicaron que pueden ser ventajosas o no (relaciones deseables o no deseables). De esta forma, las accesiones que combinen caracteres deseables, como aceptable longitud de los rebrotes y número de rebrotes, se convierten en un material a considerar para los fines de explotación en los sistemas silvopastoriles, en los que coexisten animales y plantas.
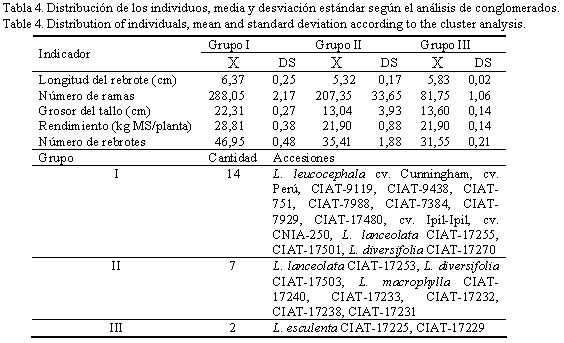
El análisis de conglomerados sobre la base de los resultados del ACP durante esta etapa, permitió la formación de tres grupos (tabla 4). De acuerdo con los resultados, las accesiones L. leucocephala cv. Cunningham, cv. Perú, CIAT-9119, CIAT-9438, CIAT-751, CIAT-7988, CIAT-7384, CIAT-7929, CIAT-17480, cv. Ipil-Ipil y cv. CNIA-250; Leucaena lanceolata CIAT-17255 y CIAT-17501; y Leucaena diversifolia CIAT-17270, pertenecientes al grupo I, se caracterizaron por ser las plantas con mayor número de ramas, longitud de los rebrotes, grosor, número de rebrotes y rendimiento, con respecto a las accesiones de los grupos II y III.
En la figura 1 se muestra un diagrama de líneas con una curva de ajuste que aproxima la relación entre el número de rebrotes y los días en cada uno de los grupos formados, donde se nota la tendencia ascendente en la emisión de nuevos rebrotes en cada uno de los grupos, a medida que transcurrían los días. El modelo que explicó con mayor bondad de ajuste esta relación (R2=0,98***, R2=0,96*** y R2=0,97*** respectivamente) fue el cuadrático. Como se puede observar, para todas las accesiones en los primeros 28 días hubo una lenta emisión de nuevos rebrotes, y a partir de los 35 días fue rápido su incremento, fundamentalmente en las que formaron el grupo I. En sentido general, estas accesiones tuvieron una capacidad de recuperación ascendente después de la poda, aspecto que pudo estar influido por su capacidad para reciclar las reservas acumuladas durante el período de establecimiento.
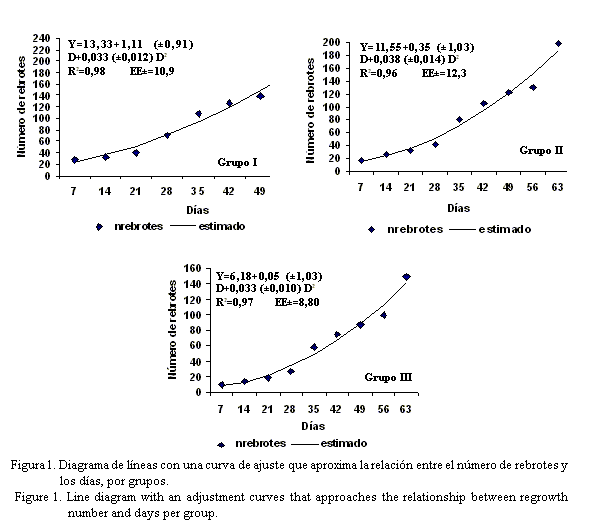
Este comportamiento quizás sea de índole específica de la especie (Pezo e Ibrahim, 1999), pero también es posible atribuirlo a los siguientes factores: la repartición de las reservas de carbohidratos no estructurales entre las diferentes partes de la planta (aérea y subterránea), la capacidad fotosintética del área foliar residual (Greaves et al., 1999), la presencia de un área adecuada de tejido parenquimático reservante y de tejido meristemático activo; y la capacidad de movilización y utilización de esas reservas. Estos son necesarios para el desarrollo del rebrote (Francisco y Simón, 2001) y en esta investigación se manifestaron de forma más eficiente en las accesiones pertenecientes al grupo I que en las de los demás grupos.
En estudios anteriores se determinó que las plantas de este género, especialmente de la especie L. leucocephala, poseen alta capacidad de rebrote aun en plena sequía, debido a las características de su sistema radical, el cual tiene un alto componente de raíces permanentes estructurales, así como un sistema de raicillas que son las responsables de la asimilación de agua y nutrientes en las capas más profundas del suelo (Pezo e Ibrahim, 1999).
Al respecto Ruiz y Febles (1987) refirieron que las plantas del género Leucaena, especialmente las accesiones de L. leucocephala, tienen la capacidad de rebrotar después de la defoliación o el ramoneo. En el caso de las demás especies no se conoce con exactitud cómo ocurre, por ser las menos utilizadas tanto nacional como internacionalmente en las investigaciones. No obstante, en la presente investigación se comportó de forma diferente, tanto entre especies como entre accesiones.
En relación con lo planteado, existe un consenso generalizado de que las reservas son importantes en los primeros días de rebrote (entre seis y ocho semanas), y que a partir de ese momento el área foliar es capaz de suplir los carbohidratos y otras sustancias necesarias para el aumento ulterior; ello se hace evidente cuando se produce un reposo prolongado, sobre todo durante el período poco lluvioso. Sin embargo, este comportamiento en los primeros días puede ser variable, como se observó en las accesiones pertenecientes a los grupos II y III, en las cuales la variabilidad se evidenció más que en las accesiones del grupo I (fig. 1). En investigaciones realizadas al respecto en Cuba, Díaz (2006b) demostró que el consumo de lípidos fue intenso entre las primeras dos y cuatro semanas, y que varió para el período lluvioso y el poco lluvioso.
A su vez, los elementos anteriores están vinculados con la disponibilidad de los recursos abióticos, como el agua y los nutrientes, y las deficiencias nutricionales y el estrés hídrico disminuyen el rango de formación del rebrote y la actividad del tejido fotosintético. Ello puede acelerar la senescencia de la hoja y disminuir la producción de biomasa comestible, aspectos que suceden independientemente de las similitudes y contrastes que existen entre los grupos e incluso entre las accesiones que los forman, como pudo haber ocurrido en esta investigación.
En la figura 2 se muestra un diagrama de líneas con una curva de ajuste que aproxima la relación entre la longitud de los rebrotes y los días, en cada uno de los grupos formados. El modelo que explicó con mayor bondad de ajuste esta relación (R2=0,95***, R2=0,95*** y R2=0,96*** respectivamente) fue el cúbico. Se observa que a medida que transcurrieron los días hubo un incremento en la longitud (velocidad de crecimiento) de los rebrotes para los tres grupos hasta los 56 días, independiente de la respuesta particular de las accesiones en estos; aunque de manera análoga a la emisión de rebrotes, su crecimiento para el grupo I fue más acelerado que para las del grupo II. No obstante, entre los 56 y 63 días diminuyó la velocidad de crecimiento de las plantas, el cual coincidió con la altura de 2,48 y 180,6 cm.
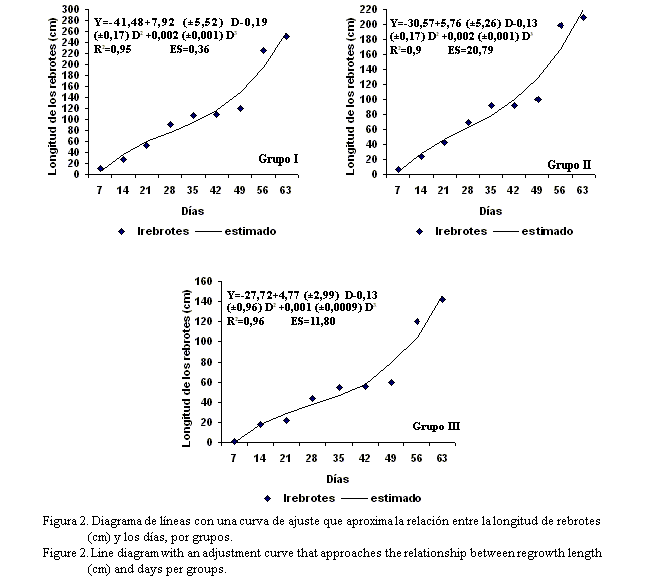
En el grupo I (tabla 4) se constató la agrupación de tres accesiones (L. lanceolata CIAT-17255 y CIAT-17501, y L. diversifolia CIAT-17270) que no son de la especie L. leucocephala, lo cual pudiera indicar que en ellas el ritmo de crecimiento de los rebrotes es muy similar al de esta especie, en lo que influye no sólo la accesión en cuestión, sino también la integración de varios factores. En este sentido, Voisin (1963) refirió que todas las plantas en su dinámica de crecimiento, en condiciones climáticas estables, describen una curva sigmoidea en tres etapas, a la cual le llamó la curva biológica universal. Los órganos de cualquier organismo también presentan estas características, de manera que el crecimiento de cualquier planta resulta de la integración de muchas curvas de cada uno de sus órganos, aspecto señalado también por Stür et al. (1994) para el caso de árboles medianos como la Leucaena.
En condiciones de campo, por efecto de los elementos del clima, estas curvas suelen alternarse en ocasiones, de manera que el carácter sigmoideo no se observa de forma clara. En Cuba varios autores, entre ellos Blanco (1996), han descrito las curvas de crecimiento para los pastos tropicales, donde puede observarse que los intervalos de tiempo en que se producen sus estadios difieren de lo informado para los pastos templados.
En el caso de los árboles y los arbustos con fines forrajeros estos principios también son válidos. Las plantas leñosas (sobre todo los árboles) se caracterizan por crecer lentamente, en comparación con las gramíneas, lo que constituye una dificultad en el establecimiento y después de esta etapa, en los sistemas silvopastoriles.
Al respecto Díaz (2006b) planteó que en el caso de las gramíneas o de las leguminosas herbáceas, las causas de este fenómeno están relacionadas, en primera instancia, con la dinámica de crecimiento y expansión foliar (que es muy lenta en sus inicios) y también con la partición de la biomasa hacia el sistema radical durante las primeras semanas. Este último aspecto tiene mayor importancia para las plantas adaptadas a condiciones de aridez (como el buffel y la alfalfa) y para las plantas leñosas está vinculado con el costo energético de la producción de tejido lignificado.
En los tres grupos, hasta los 28 días, hubo un lento crecimiento de los rebrotes (fig. 2), el cual pudo depender de la especie y el grado de intensidad con que fue defoliada (por corte o por pastoreo con animales). Este comportamiento puede que esté relacionado con la poca cantidad de área foliar, la capacidad de utilización de la reserva acumulada en las raíces y en la base de los tallos, el desarrollo del sistema asimilativo y la distribución de los fotoasimilados en los diferentes órganos. Según refiere Díaz (2006a), en este período la tasa fotosintética es alta y la respiración es baja, lo cual establece una relación positiva en la asimilación neta de la planta y, por consiguiente, un aumento en la acumulación de masa seca con predominio en las hojas.
Desde los 35 hasta los 56 días el crecimiento de los rebrotes se incrementó para todas las accesiones de los tres grupos, pero de forma más acelerada en las del grupo I (fig. 2). En este período, según indican Stür et al. (1994), se produce un marcado aumento en el área foliar, con un balance positivo entre la fotosíntesis y la respiración hasta alcanzar la máxima producción fotosintética. En dicha etapa el crecimiento se hace relativamente constante e independiente de la variable que se mide, la cual expresa la tasa máxima de crecimiento del cultivo y acumulación de reservas.
Esto se corresponde con lo informado por Bonilla (2002), acerca de que este período se caracteriza por el retraso en la tasa de muerte de las estructuras morfológicas de la planta, en relación con el correspondiente aumento en el crecimiento de los tallos, lo que hace que se exprese un incremento en la producción de tejidos y una disminución en la muerte de sus distintos componentes.
Por ello, según refiere Díaz (2006a) la producción máxima por hectárea puede que no se corresponda con la fotosíntesis máxima, ya que depende de cómo se comporte la tasa de pérdida del tejido por muerte.
El comportamiento que presentaron las accesiones del grupo I con respecto a las de los grupos II y III puede que se deba a la interacción de diversos factores, independientemente de la respuesta de cada accesión en particular. Debe considerarse además la respuesta de las plantas a las condiciones climáticas, sobre todo las lluvias caídas, que aunque no fueron tan significativas propiciaron el buen desarrollo de los rebrotes y resultaron más efectivas en unas accesiones que en otras.
La altura de corte es otro de los elementos que pudieron influir en la emisión de rebrotes y en su crecimiento (Benjamín et al., 1999); a pesar de ello, la respuesta de las accesiones fue diferente. En este sentido, debe tenerse en consideración su profundo sistema radical y la cantidad de sustancias de reserva que tenga la planta.
Se debe señalar también que el corte del forraje en las distintas estaciones del año (seca y lluviosa) y en diferentes estadios de desarrollo (vegetativo y floración), tiene una estrecha relación y actúa en el comportamiento del rebrote. Los cortes al inicio o durante el período poco lluvioso, cuando existe disminución del área fotosintéticamente activa y de la disponibilidad de agua, según describen Llamas et al. (2001), pueden provocar el agotamiento de las reservas orgánicas (como máxima fuente de su sustento), lo que conduce a la disminución del rango de formación del rebrote y los procesos de crecimiento de las plantas, como parece haber ocurrido en las accesiones pertenecientes a los grupos II y III.
El incremento de la emisión de rebrotes y su longitud de manera general, demostró la capacidad de recuperación de las accesiones; mientras que las diferencias entre los tres grupos formados evidenciaron contrastes entre las especies, según su capacidad de utilización de las reservas acumuladas. La dinámica de emisión de rebrotes y la velocidad de crecimiento de éstos en forma acelerada, tanto en la etapa de establecimiento como en la recuperación después de la poda, son de gran importancia, ya que no sólo le permiten al cultivo la supervivencia, sino también que sea utilizado como alimento animal, especialmente en el período poco lluvioso. Las accesiones mostraron una capacidad de recuperación aceptable a través del incremento de los rebrotes y su crecimiento.
La emisión de nuevos rebrotes por parte de las plantas de este género, al igual que su crecimiento, mostraron diferencias entre los grupos formados; por ello, la capacidad de recuperación no debe indicarse de forma estática, sino tomando en consideración la interacción de diversos factores tanto bióticos como abióticos, así como la posibilidad o capacidad del genoma individual de cada accesión de responder o no al medio circundante.
De acuerdo con los resultados se concluye que existieron diferencias en los indicadores y que estos explicaron la variabilidad hallada. Las especies y accesiones del género Leucaena, de forma general, mostraron alta capacidad de recuperación después de la poda según su potencialidad y exigencia para el uso de sus reservas, acorde con las variables medidas. En ese sentido, se seleccionaron las accesiones L. leucocephala CIAT-9119, CIAT-9438, CIAT-751, CIAT-7988, CIAT-7384, CIAT-7929 y CIAT-17480; L. lanceolata CIAT-17255 y CIAT-17501, y L. diversifolia CIAT-17270, además de las variedades comerciales L. leucocephala cv. Cunningham, cv. Perú, cv. CNIA-250 y cv. Ipil-Ipil, por ser las de mayor capacidad de recuperación después de la defoliación.
Se recomienda, en futuras investigaciones relacionadas con la caracterización, evaluación y selección de estas especies, realizar estudios que incluyan su persistencia en el tiempo bajo condiciones de poda, así como su empleo en el fomento de nuevos sistemas silvopastoriles.
REFERENCIAS BIBLIOGRÁFICAS
1. Academia de Ciencias de Cuba. Nuevo Atlas Nacional de Cuba. Instituto Cubano de Geodesia y Cartografía. La Habana, Cuba. p. 41. 1989
2. Alonso, J. Factores que intervienen en la producción de biomasa de un sistema silvopastoril leucaena (Leucaena leucocephala cv. Perú) y guinea (Panicum maximum cv. Likoni). Tesis presentada en opción al grado de Doctor en Ciencias Agrícolas. La Habana, Cuba. 109 p. 2003
3. Benjamín, A.K. et al. Productivity of five tree legumes species in the tropics. In: AAAP-ASAP Congress. Sydney, Australia. [cd-rom]. 1999
4. Blanco, F. Dinámica de crecimiento y variación de las reservas en Andropogon gayanus CIAT-621. Pastos y Forrajes. 19:47. 1996
5. Bonilla, I. Introducción a la nutrición mineral de las plantas. Los elementos minerales. En: Fundamentos de Fisiología vegetal. (Azcón-Bieto, J. y Talón, M., Eds.). Ediciones Universitat de Barcelona, España. 198 p. 2002
6. Borel, R. Diseño y manejo de los sistemas silvopastoriles. En: VI Curso internacional de entrenamiento. (Ed. L. Krishnamurthy). Centro de Agroforestería para el Desarrollo Sostenible. Universidad Autónoma de Chapingo. México. p. 442. 1997
7. Dávila, C. & Urbano, D. Leguminosas arbóreas en la zona sur del Lago de Maracaibo. En: Leguminosas forrajeras arbóreas en la agricultura tropical. (Ed. T. Clavero). Centro de Transferencia de Tecnología en Pastos y Forrajes. Universidad del Zulia, Venezuela. p. 101. 1996
8. Díaz, Maykelis Fisiología del crecimiento y el desarrollo. En: Compendio de conferencias del Programa de la Maestría en Pastos y Forrajes. Curso: Fundamentos de la producción de pastos. EEPF «Indio Hatuey». Matanzas, Cuba. 25 p. (Mimeo). 2006a
9. Díaz, Maykelis Fotosíntesis. En: Compendio de conferencias del Programa de la Maestría en Pastos y Forrajes. Curso: Fundamentos de la producción de pastos. EEPF «Indio Hatuey». Matanzas, Cuba. 13 p. (Mimeo). 2006b
10. Francisco, Geraldine et al. Efecto de tres alturas de corte en el rendimiento de biomasa de Leucaena leucocephala cv. CNIA-250. Pastos y Forrajes. 21:337. 1998
11. Francisco, Geraldine & Simón, L. Estudios del nivel de poda en una plantación de Leucaena leucocephala CNIA-250. Pastos y Forrajes. 24:139. 2001
12. Gómez, M.E. et al. Árboles utilizados en la alimentación animal como fuente proteica: Matarratón (Gliricidia sepium), Nacedero (Trichanthera gigantea), Pízamo (Erythrina fusca) y Botón de oro (Tithonia diversifolia). Centro para la Investigación en Sistemas Sostenibles de Producción Agropecuaria (CIPAV). Colombia. p. 3. 2002
13. Greaves, A.J. et al. Carbon supply froms starch reserves to spring growth: Modelling spacial patterns in kiwifruit canes. Annals of Botany. 83:431
14. Guerra, C.W. et al. 2003. Criterios para la selección de modelos estadísticos en la investigación científica. Rev. cubana Cienc. agríc. 73:3. 1999
15. Hernández, A. et al. Nuevos aportes a la clasificación genética de suelos en el ámbito nacional e internacional. Instituto de Suelos. Ministerio de la Agricultura. AGRINFOR. La Habana, Cuba. 145 p. 2003
16. Hernández, I. Utilización de las leguminosas arbóreas L. leucocephala, A. lebbeck y B. purpurea en sistemas silvopastoriles. Tesis presentada en opción al grado de Dr. en Ciencias Agrícolas. Instituto de Ciencia Animal. Estación Experimental de Pastos y Forrajes "Indio Hatuey", Matanzas, Cuba. 118 p. 2000
17. Hernández, I. et al. Utilización de L. leucocephala, A. lebbeck y B. purpurea en sistemas silvopastoriles. Memorias. IV Taller Internacional Silvopastoril "Los árboles y arbustos en la ganadería tropical". EEPF "Indio Hatuey". Matanzas, Cuba. p. 284. 2000
18. Llamas, E. et al. Trees forage production and quality on a quarry soil in Mérida, Yucatán, México. En: International Symposium on Silvopastoral Systems. Second Congress on Agroforestry and Livestock Production in Latin America. San José, Costa Rica. p. 355. 2001
19. Molina, C.H. et al. Advances in the implementation of high tree density in sylvopastoral systems. En: International Symposium on Sylvopastoral Systems. Second Congress on Agroforestry and Livestock Production in Latin America. San José, Costa Rica. p. 299. 2001.
20. Morrison, D. Multivariate statistical methods. Mc Graw-Hill Book Company. New York, USA.150 p. 1967.
21. Pezo, D. & Ibrahim, M. Sistemas silvopastoriles. Módulo de Enseñanza Agroforestal No. 2. CATIE-GTZ. Turrialba, Costa Rica. 275 p. 1999.
22. Ruiz, T.E. & Febles, G. Leucaena una opción para la alimentación bovina en el trópico y el subtrópico. EDICA. La Habana, Cuba. 200 p. 1987.
23. Ruiz, T.E. & Febles, G. Factores que influyen en la producción de biomasa durante el manejo del sistema silvopastoril. En: Memorias del curso «Sistemas silvopastoriles una opción sustentable». Tantakin, México. p. 62. 2001.
24. Sánchez, Saray. Acumulación y descomposición de la hojarasca en un pastizal de Panicum maximum Jacq. y en un sistema silvopastoril de Panicum maximum y Leucaena leucocephala (Lam) de Wit. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas. ICA. La Habana, Cuba. 123 p. 2007.
25. Seguí, Esperanza et al. Asociaciones entre caracteres individuales y su importancia en el mejoramiento genético de la especie Panicum maximum Jacq. Pastos y Forrajes. 12:219. 1989.
26. Simón, L. Impacto bioeconómico y ambiental de la tecnología del silvopastoreo racional. En: El Silvopastoreo: Un nuevo concepto de pastizal (Ed. L. Simón). Estación Experimental de Pastos y Forrajes "Indio Hatuey". Matanzas, Cuba-Universidad de San Carlos de Guatemala, Guatemala. p. 203. 2005.
27. Stür, W.W. et al. Defoliation and management of forage tree legumes. En: Tree legumes in tropical agriculture. (Gutteridge R.C. and Shelton, H.M, eds). CAB International. Wallingford, UK. p. 144. 1994.
28. Toral, Odalys et al. Comportamiento del germoplasma arbóreo forrajero en condiciones de Cuba. Pastos y Forrajes. 29:337. 2006.
29. Torres, Verena et al. Modelo estadístico para la medición del impacto de la innovación o transferencia tecnológica en la rama agropecuaria. Informe técnico. Instituto de Ciencia Animal. La Habana, Cuba. 2006.
30. Visuata, B. Análisis estadístico con SPSS para Windows. Vol. II. Estadística multivariante. (Ed. C. Fernández). Editoral Mc Graw-Hill. Madrid, España. p. 24. 1998
31. Voisin, A. Productividad de la hierba. Editorial Tecnos. S. A. Madrid. 499 p. 1963.
Recibido el 22 de septiembre del 2010
Aceptado el 23 de noviembre del 2010
