










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes vol.34 no.3 Matanzas jul.-sep. 2011
ARTÍCULO DE INVESTIGACIÓN
Composición funcional de la macrofauna edáfica en cuatro usos de la tierra en las provincias de Artemisa y Mayabeque, Cuba
Functional composition of soil macrofauna in four land uses of Artemisa and Mayabeque provinces, Cuba
Grisel Cabrera1, Nayla Robaina2 y D. Ponce de León3
1 Instituto de Ecología y Sistemática, CITMA, Carretera de Varona km 3½, Capdevila, Boyeros, C.P. 10800, La Habana, Cuba
E-mail: grisel17@ecologia.cu
2 Estación Experimental de Plantas Medicinales, Artemisa, Cuba
3 Facultad de Agronomía, Universidad Agraria de La Habana, Artemisa, Cuba
RESUMEN
El estudio tuvo como objetivo analizar la composición funcional de la macrofauna del suelo en función de diferentes usos de la tierra: bosques secundarios, pastizales, áreas de cultivos varios destinadas a la producción de papa y cañaverales. Estos sistemas de uso de la tierra estaban localizados en las provincias Artemisa y Mayabeque. En cada área la macrofauna edáfica fue recolectada en octubre, al final del período lluvioso del año 2009, siguiendo la metodología del TSBF. El procesamiento de los datos incluyó la determinación de la densidad y la biomasa de los grupos funcionales de la macrofauna, y el análisis estadístico involucró la prueba no paramétrica de Kruskal-Wallis. Los grupos funcionales de mayor representatividad, en densidad, en la mayoría de los usos de la tierra fueron los ingenieros del suelo y/o los detritívoros, y en biomasa igualmente los detritívoros y/o los herbívoros. La proporción entre los grupos funcionales de la macrofauna del suelo en los diferentes usos dependió de la intensidad de uso de la tierra, el nivel de perturbación del medio edáfico y la disponibilidad de recursos.
Palabras clave: Organismos del suelo, uso múltiple de la tierra.
ABSTRACT
The objective of this study was to analyze the functional composition of soil macrofauna in different land use types including secondary forest, pastures, fields with varied crop cultivation destined to potato production and sugarcane fields. The land use systems were located in the Artemisa and Mayabeque provinces. In these systems the edaphic macrofauna was collected in October, at the end of the rainy season of 2009, using the TSBF methodology. The data processing included the determination of the density and biomass of the macrofauna functional groups in each land use, and the statistical analysis implicated the Kruskal-Wallis nonparametric test. The soil engineers and/or detritivores communities were observed to be better represented in density in the majority of land uses, and in biomass, the detritivore and/or herbivore functional groups. The proportion between the functional groups of soil macrofauna in the land uses depended on land use intensity, the disturbance level of the edaphic environment and the availability of resources.
Key words: Soil organism, multiple land use.
INTRODUCCIÓN
La actividad que desempeñan los diferentes grupos funcionales que componen la macrofauna del suelo, entre ellos los ingenieros del suelo, los detritívoros, los herbívoros y los depredadores, permite la regulación de los procesos edáficos y el funcionamiento y equilibrio del ecosistema.
Los ingenieros del suelo, que involucran grupos consumidores de materia orgánica como las lombrices (Haplotaxida) y las termitas (Isoptera), y los organismos omnívoros como las hormigas (Hymenoptera: Formicidae), tienen un impacto específico en el interior del suelo a partir de la transformación de sus propiedades físicas, que favorecen la formación de agregados y la estructura, el movimiento y la retención del agua, así como el intercambio gaseoso (Lavelle, 2000). Los detritívoros, tales como milpiés (Spirobolida, Polydesmida y otros), cochinillas (Isopoda) y caracoles (Archaeogastropoda) fundamentalmente, tienen una función a nivel de la superficie del suelo como organismos epígeos, ya que al alimentarse de la hojarasca ayudan en su fragmentación e inician el proceso de descomposición, aumentando la superficie de exposición para el ataque de la microflora.
Los herbívoros incluyen algunas familias de coleópteros (Coleoptera), hemípteros (Hemiptera) y otros órdenes de insectos, y los depredadores primordialmente a los arácnidos (Araneae, Opiliones, Pseudoscorpionida) y ciempiés (Geophilomorpha, Scolopendromorpha), los cuales intervienen a otros niveles de la cadena trófica al consumir material vegetal y animal vivo, respectivamente, lo que origina la riqueza y calidad de estos materiales en el suelo (Lavelle, 1997). De acuerdo con lo planteado por Zerbino et al. (2008), las interacciones entre todos los grupos funcionales están determinadas por los recursos disponibles en los distintos usos de la tierra.
En un gradiente desde ecosistemas seminaturales hasta agroecosistemas existen características peculiares en el medio edáfico, incluyendo su biota asociada. En los ecosistemas del trópico húmedo tales como bosques, sistemas agroforestales, pastizales y policultivos, numerosos autores han detectado la dominancia en términos de densidad de los artrópodos detritívoros y los grupos ingenieros del ecosistema; mientras que dentro de los ingenieros, las lombrices de tierra endógeas han constituido el mayor componente en biomasa (Barros et al., 2002; Huerta-Lwanga et al., 2008). Granados y Barrera (2007) plantearon que los primeros colonizadores de la macrofauna edáfica son aquellos capaces de explotar el contenido de materia orgánica en los sistemas con un aporte continuo y abundante de esta, seguidos por los depredadores que se alimentan de los grupos favorecidos y, por último, van apareciendo grupos omnívoros y herbívoros. En el caso de los sistemas herbáceos, con influencia casi total de gramíneas y donde hay gran cantidad de fitomasa aérea y subterránea, los mayores componentes de la macrofauna han sido los herbívoros; también han estado favorecidas las lombrices y las termitas, estimuladas por la mineralización del carbono a partir de los exudados radicales (Laossi et al., 2008).
En Cuba aún son insuficientes los estudios que han hecho énfasis en la separación de los grupos funcionales de la macrofauna edáfica para el análisis de su función en los ecosistemas. En este sentido se pueden mencionar algunos trabajos desarrollados específicamente con las comunidades de lombrices de tierra (Rodríguez, 2000; Martínez, 2002), aunque existen estudios de la macrofauna que han tenido en cuenta la caracterización de su composición funcional y trófica en sistemas agroecológicos en el país (Cabrera et al., 2007).
Este estudio tuvo como objetivo el análisis de la composición funcional de la macrofauna del suelo en cuatro usos de la tierra, cuyos sistemas de estudio se localizaron en las provincias Artemisa y Mayabeque.
MATERIALES Y MÉTODOS
Descripción de los usos de la tierra estudiados
Se estudió un gradiente de uso de la tierra que involucró cuatro usos: bosques secundarios, pastizales, áreas de cultivos varios destinadas a la producción de papa y cañaverales. Estos usos se escogieron dentro de la Llanura Roja de la antigua provincia La Habana, actualmente localizada entre las provincias Artemisa y Mayabeque. El tipo de suelo en todos los usos fue Ferralítico Rojo, según la clasificación de Hernández et al. (1999).
Dentro del uso bosques secundarios, se trabajaron tres áreas ubicadas en las localidades de Managua, Nazareno (municipio San José de las Lajas) y Aguacate (municipio Madruga) de la provincia Mayabeque. Estas áreas tuvieron diferentes niveles de regeneración/antropización, edades de 40 a 120 años y extensiones entre dos y cinco hectáreas.
Presentaron vegetación del tipo semidecidua, donde dominaron en el estrato arbóreo especies frutales y maderables, entre ellas: mamey colorado (Pouteria sapota H.E. Moore & Stearn), mango (Mangifera indica L.), caimito (Chrysophyllum cainito L.), framboyán rojo (Delonix regia (Boj. ex Hooker) Raf.), palma real (Roystonea regia (Kunth) O.F. Cook), baría (Cordia gerascanthus L.), ocuje (Calophyllum inophyllum L.), yamagua (Guarea guidonia (L.) Sleumer), teca (Tectona grandis L. fil.), caoba de Cuba (Swietenia mahagoni (L.) Jacq.), cedro (Cedrela odorata L.), pino (Pinus sp.) y majagua (Talipariti elatum (L.) Fryxell).
Para el uso pastizales se escogieron dos áreas de trabajo en el municipio San José de las Lajas de la provincia Mayabeque, con una extensión máxima de 12 ha. Son pastizales naturalizados con más de 25 años de explotación y las especies de pastos principales eran: hierba de guinea (Panicum maximum Jacq.) y pasto estrella (Cynodon nlemfuensis Vanderyst), que presentaban una cobertura entre 80 y 90%. En ellos pastoreaba ganado vacuno de las razas Holstein y Siboney, y el manejo era principalmente rotacional racional intensivo con carga animal media de 2,8 UGM ha-1.
En el caso de los cultivos varios, cuyo cultivo principal fue la papa (Solanum tuberosum L.), se seleccionaron tres áreas en los municipios Güira de Melena (provincia Artemisa), Batabanó y Güines, pertenecientes a Mayabeque. Los sitios seleccionados tenían extensiones entre 20 y 50 ha y el cultivo principal se encontraba en constante rotación con boniato (Ipomoea batata (L.) Lam.), malanga (Colocasia esculenta (L.) Schott), frijol (Phaseolus vulgaris L.), maíz (Zea mays L.), calabaza (Cucurbita moschata (Duch. ex Lam.) Duch. ex Poir) y yuca (Manihot esculenta (L.) Crantz). Entre la vegetación indeseable en las áreas se destacaron: don carlos (Sorghum halepense (L.) Pers.), cebolleta (Cyperus rotundus L.), bledo (Amaranthus hybridus L.) y romerillo (Bidens pilosa L.). Son áreas dedicadas a la agricultura por más de 10 años, fundamentalmente con labranza tradicional y riego eléctrico por aspersión. La aplicación de NPK en estos sistemas fue de 1 490 kg ha-1 año-1 y la de urea osciló entre 224 y 298 kg ha-1 año-1.
Para el uso cañaverales, los tres sitios escogidos con caña de azúcar (Saccharum officinarum L.) se ubicaron en los municipios Güira de Melena (provincia Artemisa), San Nicolás de Bari y Madruga pertenecientes a Mayabeque, ocupados por las variedades CP 52-43 y C 86-12, C 323-68 y C 86-56, respectivamente. Cada área de caña de azúcar estudiada tiene más de 20 años de explotación y una extensión de alrededor de 15 ha. Las plantas arvenses fundamentales en los sistemas fueron don carlos, cebolleta, sancaraña (Rottboellia cochinchinensis L.F.) y pata de gallina(Eleusine indica (L.) Gaertn). Poseen sistemas con riego por gravedad y sistema eléctrico de enrolladores; la fertilización química fue de: 50-60 kg ha-1 año-1 de urea, 20-83 kg ha-1 año-1 de K2O y 25 kg ha-1 año-1 de P2O5.
Muestreo y procesamiento de la macrofauna edáfica
El muestreo se hizo al final del período lluvioso del año 2009, en el mes de octubre. Este es el momento descrito como idóneo para la recolecta, debido a la mayor actividad de la macrofauna. En las diferentes áreas de uso de la tierra, la recolección de la macrofauna se realizó según la Metodología del Programa Internacional «Biología y Fertilidad del Suelo Tropical» o TSBF (Anderson e Ingram, 1993; Lavelle et al., 2003). En cada área o réplica de uso de la tierra se extrajeron ocho monolitos de suelo de 25 x 25 cm, hasta 30 cm de profundidad, distanciados entre sí a 20 m. El muestreo respondió a un diseño completamente aleatorizado, donde la macrofauna se recolectó manualmente in situ.
Desde el punto de vista funcional la macrofauna fue agrupada en cuatro gremios fundamentales: ingenieros del suelo, detritívoros, herbívoros y depredadores, de acuerdo con Lavelle (1997) y Zerbino et al. (2008). Se realizó el análisis solamente de estos cuatro grupos funcionales, ya que tienen definida su función en el ecosistema y el efecto que causan sobre la transformación de las propiedades del suelo. La identificación taxonómica se realizó a nivel de orden y familia, y solo un orden fue identificado hasta subfamilia para definir el grupo funcional (Brinkhurst y Jamieson, 1971; Borror et al., 1976; Sims, 1980; Pérez-Asso, 1995, 1996, 1998; Brusca y Brusca, 2003). Se determinó la densidad en función del número de individuos y la biomasa a partir del peso húmedo en la solución preservante, de los diferentes grupos funcionales para cada uso de la tierra.
Con el propósito de determinar las variaciones de la densidad y la biomasa para cada uno de los grupos funcionales, entre usos, se empleó el análisis no paramétrico de Kruskal-Wallis y como prueba a posteriori para la comparación de medias, se utilizó la U de Mann-Whitney con ajuste de Bonferroni. El procesamiento estadístico se realizó a través del programa PAST-1.75, 2001.
RESULTADOS Y DISCUSIÓN
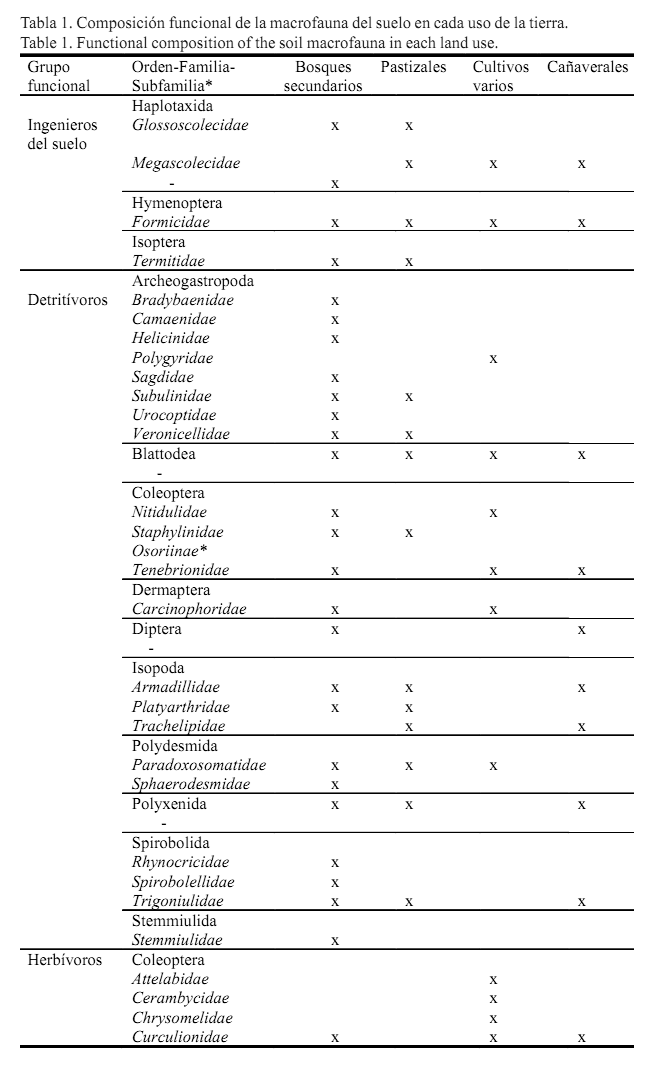
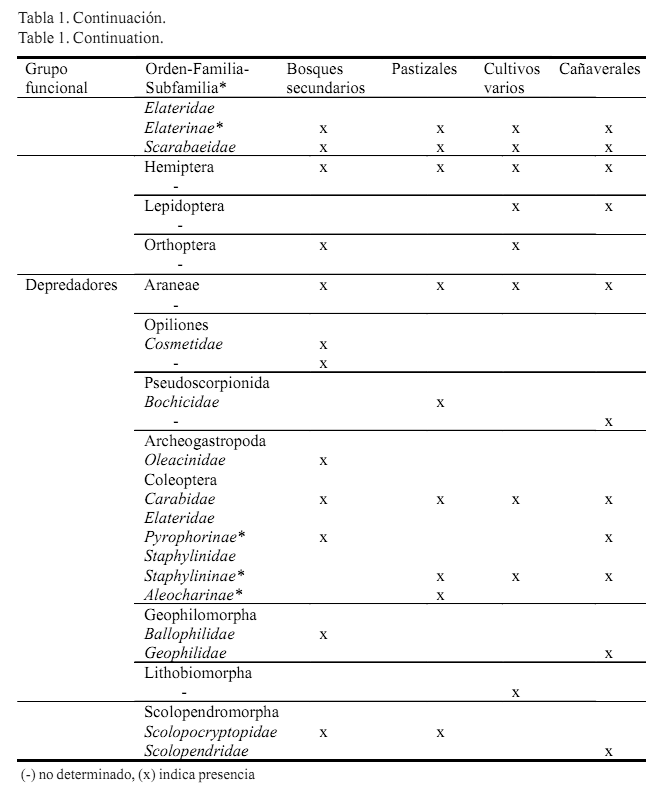
La macrofauna pudo ser asociada a los grupos funcionales de ingenieros del suelo, detritívoros, herbívoros y depredadores, los cuales estuvieron siempre presentes en todos los usos de la tierra. Dentro de los detritívoros, los órdenes de la macrofauna comúnmente encontrados en la mayoría de los usos fueron Blattodea, Isopoda, Polydesmida, Polyxenida y Spirobolida, y de Coleoptera solo la familia Tenebrionidae. En el caso de los herbívoros, se observaron en todos los usos las larvas de coleópteros pertenecientes a las familias Scarabaeidae y Elateridae (Elaterinae), cuyos hábitos alimentarios responden a la herbivoría de raíces, y al orden Hemiptera que se incluye como herbívoros foliares (afectan el follaje pero se pueden encontrar en el suelo). Para los depredadores, las unidades taxonómicas coincidentes fueron Araneae, Scolopendromorpha y adultos de Coleoptera de las familias Carabidae y Staphylinidae (Staphylininae)(tabla 1a y b ).
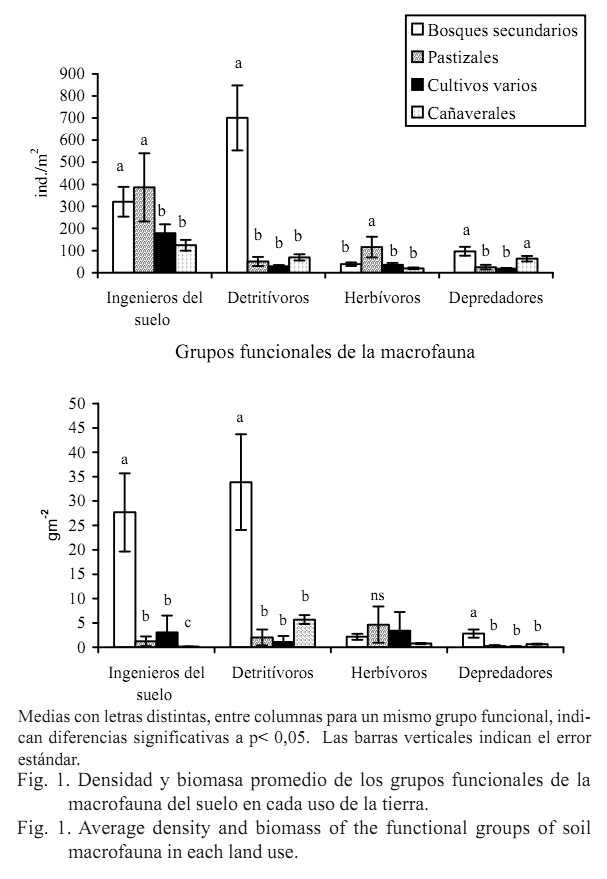
En cuanto a la densidad, en los bosques secundarios prevalecieron los detritívoros (701 ind.m-2) seguidos en una menor cantidad por los ingenieros (320 ind.m-2), los depredadores (96 ind.m-2) y, por último, los herbívoros (38 ind.m-2). En los pastizales y los cultivos varios dominaron los ingenieros del suelo (386 y 179 ind.m-2, respectivamente) seguidos de los herbívoros (116 y 36 ind.m-2), y con menor densidad se encontraron los detritívoros (51 y 28 ind.m-2) y los depredadores (25 y 18 ind.m-2). Respecto a esta misma variable, en los cañaverales también se ubicaron en primer lugar los ingenieros del suelo (124 ind.m-2) y posteriormente, casi a un mismo nivel, los detritívoros (69 ind.m-2) y los depredadores (63 ind.m-2); los menos representados en este uso fueron los herbívoros (19 ind.m-2) (fig.1).
En el caso de la biomasa, los bosques secundarios respondieron al mismo patrón de la densidad, con el orden decreciente de detritívoros (33 gm-2) - ingenieros (27 gm-2) - depredadores (2,8 gm-2) - herbívoros (2,1 gm-2). En los pastizales se hallaron como dominantes los herbívoros (4,6 gm-2) y a continuación los detritívoros (2 gm-2), ingenieros (1,2 gm-2) y depredadores (0,2 gm-2). En los cultivos varios predominaron casi al unísono herbívoros (3,4 gm-2) e ingenieros (3,0 gm-2), y con biomasa muy baja siguieron los detritívoros (1,1 gm-2) y los depredadores (0,1 gm-2). En los cañaverales se destacaron los detritívoros
(5,6 gm-2), y el resto de los grupos funcionales estuvieron pobremente representados en biomasa en este uso (herbívoros: 0,7 gm-2, depredadores: 0,6 gm-2, ingenieros: 0,1 gm2) (fig.1).
Se obtuvieron diferencias significativas entre los usos para los diferentes grupos funcionales, tanto en densidad como en biomasa (p<0,05), excepto en los herbívoros respecto a la biomasa (p>0,05).
Los ingenieros del suelo en los bosques secundarios y en general en todos los usos, respecto a la densidad, tuvieron una fuerte influencia de termitas u hormigas, y en la biomasa se destacaron las lombrices de tierra. Luizão (1995) y Barros et al. (2002) también encontraron mayor densidad de estos insectos sociales entre toda la fauna edáfica en los bosques, los pastizales y los sistemas de cultivos de la Amazonía brasileña, y Linares et al. (2009) detectaron a los ingenieros del suelo, incluyendo las lombrices de tierra, como los más abundantes en los bosques y los sistemas agroforestales de Perú.
En particular, el comportamiento de las lombrices de tierra indicó el nivel de intervención antrópica por la intensidad de uso de la tierra y el grado de perturbación del medio edáfico, como también fue hallado por diversos autores al evaluar el cambio de estas comunidades ante el impacto de diferentes usos de la tierra y por el efecto de distintos manejos del terreno en el trópico (Rodríguez, 2000; Feijoo et al., 2007). Los bosques presentaron los mayores valores de densidad (103 ind.m-2) y biomasa (27 gm-2) de lombrices de tierra, a pesar de ser de formación secundaria pero con mayor estabilidad al no tener actividades de laboreo continuo ni pastoreo, con relación a los restantes usos. Le siguieron los pastizales en cuanto a la densidad (31 ind.m-2), los cuales se caracterizan por un grado de intervención media por acción solo de la actividad de pastoreo, y por último los cultivos varios (21 ind.m-2) y los cañaverales (1,33 ind.m-2) que tienen un nivel superior de perturbación por la práctica reiterada de procedimientos agrícolas y menor sombra debido a la ausencia del estrato arbóreo; esto último también es característico en el uso de pastizales, lo que provoca severos daños en las poblaciones de lombrices. No obstante, los cultivos varios tuvieron valores de biomasa ligeramente más altos (2,93 gm-2) que los pastizales (0,88 gm-2) y los cañaverales (0,012 gm-2), lo que pudo deberse a que en algunos de los sistemas de cultivos varios hubo mayor cobertura del suelo y riqueza de exudados radicales, como producto de la invasión de plantas arvenses y la presencia de cultivos de rotación como el frijol que beneficia el entorno edáfico por el aporte de nutrientes, todo lo cual pudo resultar en provecho de las lombrices de tierra.
La proporción de detritívoros dentro de la comunidad de la macrofauna, también puede indicar el estado de perturbación de un ecosistema. Los detritívoros en el presente estudio, como ya fue mencionado, tuvieron una mejor representación en densidad y biomasa en los bosques secundarios, y los más reducidos valores en los pastizales y los cultivos varios. En el uso cañaverales, en biomasa, los detritívoros constituyeron el grupo predominante en relación con los otros grupos funcionales, pero con valores muy bajos y similares a los de los pastizales y los cultivos varios.
Acerca de la afectación de la comunidad de detritívoros en los sistemas con diferente grado de perturbación, Rodríguez et al. (2002) informaron que un factor definitivo para su establecimiento es la calidad de la hojarasca expresada por una baja relación C/N; en cambio, Zerbino et al. (2008) enfatizaron que los detritívoros, como organismos desprotegidos en la superficie del suelo, se reducen drásticamente por las variaciones bruscas en las condiciones de temperatura y humedad de este debido a la menor cobertura y cantidad de residuos, y a una mayor exposición a la radiación solar en aquellos ecosistemas alterados. Estas características imperaron en los usos de este estudio que tenían menos sombra y un manejo más profundo y constante del suelo; de ahí que hayan manifestado una comunidad reducida de detritívoros respecto a los bosques secundarios, principalmente los pastizales y los cultivos varios.
Por su parte, Barraqueta (2001) y Linares et al. (2009) afirmaron que las transformaciones en las condiciones ambientales del suelo originadas por la actividad agrícola y la consecuente destrucción mecánica de los microhábitats, tienen una repercusión negativa sobre los principales grupos descomponedores de la materia orgánica, como los milpiés, cochinillas y caracoles. También expresaron que la aplicación de tratamientos menos fuertes que permitieran una conservación, en mayor o menor medida, de la vieja capa de hojarasca y de la estructura del suelo, posibilitaría una protección fundamental de estas comunidades faunísticas y de su función.
Los herbívoros no solo tuvieron una fuerte constitución en densidad y biomasa en los usos pastizales y cultivos varios; también en este último estuvieron representados por algunas familias del orden Coleoptera consideradas plagas y que son frecuentes en los cultivos agrícolas (por ejemplo: Attelabidae y Chrysomelidae, tabla 1a y b ).
Es válido referir que en el estudio en general, el orden Coleoptera fue el de mayor variedad de grupos funcionales (familias detritívoras, herbívoras y depredadoras), con incidencia tanto de larvas como de adultos. Villalobos et al. (2000) señalaron que cada estado del desarrollo (larva o adulto) en diferentes organismos del suelo, puede desempeñar un papel exclusivo en el ambiente edáfico en un momento dado. Estos autores destacaron además a Elateridae y Melolonthidae como familias más comunes de Coleoptera y con presencia de larvas y/o adultos, en el perfil edáfico. Otras familias importantes en el suelo son Curculionidae, Tenebrionidae, Nitidulidae, Carabidae y Staphylinidae (Menéndez, 2010). Las larvas de Coleoptera, con su hábito de vida endógeo, pueden actuar en la transformación de las propiedades físicas del suelo, y los adultos explotan principalmente los recursos útiles de la superficie.
En este trabajo, las larvas de Scarabaeidae y Elateridae (Elaterinae) como herbívoros de raíces, pudieron beneficiarse debido a la disponibilidad del recurso alimentario por la mayor densidad de raíces, sobre todo de las gramíneas en los pastizales, y de los cultivos y las plantas arvenses acompañantes en los sistemas de cultivos varios. Según Hernández (1999), en los ecosistemas de sabanas y pastizales el desarrollo radical puede exceder el 80% del peso total de la planta, y menciona que grandes cantidades de raíces sirven de soporte a los microorganismos y a la fauna de la comunidad edáfica.
Las áreas de cultivos varios, por su parte, están sometidas a un sistema de rotación, y de acuerdo con lo planteado por Fraser (1994), la rotación de diferentes cultivos en el tiempo diversifica la biomasa vegetal aérea y produce modificación en el ambiente edáfico por la selectividad en la absorción y el aporte de nutrientes, debido a las excreciones radiculares y la entrada de detritos y raíces en el medio, todo lo cual contribuye a la presencia de diversos organismos, entre ellos los herbívoros foliares.
En la mayor representación de depredadores (en densidad y biomasa) en los usos bosques secundarios y cañaverales, quizás influyó la mayor cantidad de detritívoros en estos usos, que pudieron servir de alimento a dicho grupo funcional. No obstante, Pontégnie et al. (2005) asociaron la presencia de determinados grupos depredadores, entre ellos Araneae, con factores abióticos como la temperatura y la humedad, y no con la disponibilidad de sus presas. Estos mismos autores también señalaron algunos órdenes medianamente sensibles a los disturbios en el medio edáfico, tales como Geophilomorpha y Scolopendromorpha de la clase Chilopoda. Por el contrario, para las familias Carabidae y Staphylinidae del orden Coleoptera, se menciona la importancia de la cantidad de alimento disponible en la selección de sus hábitats, y son consumidores principalmente de huevos, larvas y adultos de otros pequeños insectos (Baloriani et al., 2009).
Anteriormente Lee (1985) informó que los sistemas de cultivos anuales intensivos alojaban menores poblaciones de depredadores, y con posterioridad Zerbino et al. (2008) encontraron que los depredadores presentaban una mayor riqueza y abundancia en los sistemas de cultivos continuos, y Araneae es uno de los taxones fundamentales; ello coincide, en parte, con los resultados del presente trabajo, ya que la aparición de este orden no solo fue en los cañaverales y cultivos varios, también estuvo presente en los pastizales y los bosques secundarios. Estos últimos autores constataron que los ambientes que se caracterizaron por bajos contenidos en materia orgánica, altos valores de fósforo y de arcilla y conductividad eléctrica, se correlacionaron positivamente con este grupo funcional.
De modo general, como aseguró Zerbino et al. (2008) las diferencias en la proporción de los grupos funcionales de las comunidades de la macrofauna en diferentes usos de la tierra, pueden ser atribuibles a la riqueza de las especies vegetales, a los cambios en las propiedades del suelo y al manejo, que a su vez determinan la cantidad y la calidad de los recursos y afectan las interacciones entre los grupos.
CONCLUSIONES
Los grupos funcionales de mayor representatividad en densidad en todos los usos de la tierra fueron los ingenieros del suelo y/o los detritívoros; posteriormente se ubicaron los herbívoros y los depredadores, excepto en los pastizales y los cultivos varios donde los herbívoros ocuparon el segundo lugar en importancia. En biomasa, igualmente los detritívoros y/o los herbívoros fueron dominantes en los usos estudiados; no obstante, en los bosques secundarios este último grupo presentó la biomasa más baja. Con valores muy bajos de biomasa prevalecieron los ingenieros y los depredadores, aunque los ingenieros se destacaron como segundo grupo en los bosques secundarios y los cultivos varios por influencia de las lombrices de tierra. La proporción de los diferentes grupos funcionales de la macrofauna del suelo varió en función de la intensidad de uso de la tierra, el nivel de perturbación del medio edáfico y la disponibilidad de recursos.
AGRADECIMIENTOS
Los resultados se obtuvieron dentro del marco de la Maestría de la Ciencia del Suelo de la Universidad Agraria de La Habana, financiados por el Proyecto de Investigación 08-22 «Causas de la degradación de la estructura de los suelos Ferralíticos Rojos de la Llanura Roja de La Habana», del PRCT Protección del Medio Ambiente y el Desarrollo Sostenible Cubano.
REFERENCIAS BIBLIOGRÁFICAS
1. Anderson, J.M. &. Ingram, J.S.I. Tropical soil biology and fertility. A handbook of methods. CAB International. Wallingford, UK. 221 p.1993.
2. Baloriani, G. et al. Abundancia y riqueza de la macrofauna edáfica (Coleoptera y Araneae) en invernáculos convencionales y en transición agroecológica. Arana. Argentina. Revista Brasileña de Agroecología. 4 (2):1733. 2009
3. Barraqueta, P. La fauna del suelo: entre la repoblación forestal y la tala. En: Conservación, uso y gestión de los sistemas forestales. VI Jornada de Urdaibai sobre desarrollo sostenible (Coord. M.D. Salinas, M.C. de la Huerga y C. Giménez ). Departamento de ordenación del territorio, vivienda y medio ambiente, Gobierno Vasco, España. p. 257. 2001
4. Barros, E. et al. Effects of land-use system on the soil macrofauna in western Brazilian Amazonia. Biology and Fertility of Soils. 35 (5):338. 2002
5. Borror, D. et al. An introduction to the study of Insects. Holt, Rinehart and Winston, Inc. New York. 852 p. 1976
6. Brinkhurst, R.O. & Jamieson, B.G.M. Aquatic Oligochaeta of the world. Oliver and Boyd. Edinburgh, Scotland. 860 p . 1971
7. Brusca, R. & Brusca, G. Invertebrates. Sinauer Associates. Sunderland, Massachusetts, USA. 936 p. 2003
8. Cabrera Dávila, G. et al. La macrofauna del suelo en sistemas agroecológicos en Cuba. Brenesia. 67:45. 2007
9. Feijoo, A. et al. Relaciones entre el uso de la tierra y las comunidades de lombrices en la cuenca del río La Vieja, Colombia. Pastos y Forrajes. 30 (2):235. 2007
10. Fraser, B. The impact of soil and crop management practices on soil macrofauna. In: Soil biota. Management in sustainable farming systems (Eds. C.E. Pankhurst, B.M. Doube, V.V. Gupta y P. R. Grace). CSIRO. Melbourne, Australia. p. 125. 1994
11. Granados, A. & Barrera, J.I. Efecto de la aplicación de biosólidos sobre el repoblamiento de la macrofauna edáfica en la cantera Soratama, Bogotá, DC. Universitas Scientiarum, Revista de la Facultad de Ciencias. Edición especial II. 12:73. 2007
12. Hernández, A. et al. Nueva versión de la clasificación genética de los suelos de Cuba. Ministerio de la Agricultura. La Habana, Cuba. 26 p. 1999
13. Hernández, L. Fitomasa subterránea en un pastizal de Paspalum notatum en la Sierra del Rosario, Cuba. Tesis presentada en opción al título académico de Máster en Ecología y Sistemática Aplicada. Instituto de Ecología y Sistemática - Ministerio de Ciencia, Tecnología y Medio Ambiente. La Habana, Cuba. 81 p. 1999
14. Huerta-Lwanga, E. et al. Relación entre la fertilidad del suelo y su población de macroinvertebrados. Terra Latinoamericana. 26 (2):171. 2008
15. Laossi, K.R. et al. Effects of plant diversity on plant biomass production and soil macrofauna in Amazonian Pastures. Pedobiología. 51:397. 2008
16. Lavelle, P. Faunal activities and soil processes: Adaptive strategies that determine ecosystem function. Adv. Ecol. Res. 24:93. 1997
17. Lavelle, P. Ecological challenges for soil science. Soil Sciences. 165:73. 2000
18. Lavelle, P. et al. Soil macrofauna. In: Trees, crops and soil fertility. Concepts and research methods. (Eds. G. Schroth & F.L.Sinclair). CABI Publishing. UK. p. 303. 2003
19. Lee, K.E. Earthworms: their ecology and relationships with soil and land use. Academic Press. New York, USA. 411 p. 1985
20. Linares, D. et al. Macrofauna del suelo en diferentes sistemas de uso de la tierra en el Parque Nacional Tingo María, Huánuco, Perú. http://www.iamazonica.org.br/conteudo/eventos/biodiversidadeSolo/pdf/resumos /Poster_DaliaL.pdf. 2009
21. Luizão, F.J. Ecological studies in contrasting forest types in Central Amazonia. Tesis presentada en opción al grado científico de Doctor en Ciencias. Universidad de Stirling. Reino Unido. 288 p. 1995
22. Martínez, M.A. Comunidades de oligoquetos (Annelida: Oligochaeta) en tres ecosistemas con diferente grado de perturbación en Cuba. Tesis presentada en opción al grado científico de Doctor en Ciencias Biológicas. Instituto de Ecología y Sistemática-Ministerio de Ciencia, Tecnología y Medio Ambiente. La Habana, Cuba. 93 p. 2002.
23. Menéndez Rivero, Y.I. Caracterización de la coleopterofauna edáfica en tres formaciones vegetales del área protegida de recursos manejados de Escaleras de Jaruco-Loma «El Cheche» (La Habana, Cuba). Tesis de Diploma. Facultad de Biología, Universidad de La Habana. La Habana, Cuba. 80 p. 2010.
24. Pérez-Asso, A.R. A new milliped genus of the family Chelodemidae (Diplopoda: Polydesmida) from Cuba. Insecta Mundi. 9 (1-2):171. 1995
25. Pérez-Asso, A.R. The genus Nesobolus (Diplopoda:Spirobolida: Rhinocricidae) in Cuba. Insecta Mundi. 10(1-4):1 . 1996
26. Pérez-Asso, A.R. Revisión y nuevas especies del género Spirobolellus (Diplopoda: Spirobolellidae) en la Isla de Cuba. Caribbean Journal of Science. 34 (1-2):67. 1998
27. Pontégnie, M. et al. Impacts of silvicultural practices on the structure of hemi-edaphic macrofauna community. Pedobiología. 45:199. 2005
28. Rodríguez, C. Comunidades de lombrices de tierra en ecosistemas con diferente grado de perturbación. Rev. Biología. 14 (2):147. 2000
29. Rodríguez, I. et al. Comportamiento de la macrofauna del suelo en pastizales con gramíneas naturales puras o intercaladas con leucaena para la ceba de toros. Rev. cubana. Cienc. Agríc. 36 (2):181. 2002
30. Sims, R.W. A classification and the distribution of earthworms suborder Lumbricina (Haplotaxida:Oligochaeta). Bull. Br. Mus. Nat. Hist. (Zool.). 39 (2):103. 1980
31. Villalobos, F.J. et al. Patrones de la macrofauna edáfica en un cultivo de Zea mais durante la fase postcosecha en la Mancha, Veracruz, México. Acta Zoológica Mexicana. 80:167. 2000
32. Zerbino, S. et al. Evaluación de la macrofauna del suelo en sistemas de producción en siembra directa y con pastoreo. Agrociencia. 12 (1):44. 2008
Recibido el 25 de noviembre del 2010
Aceptado el 7 de marzo del 2011

{kind=link}
{kind=link}
{kind=link}