










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes vol.35 no.3 Matanzas jul.-set. 2012
ARTÍCULO DE INVESTIGACIÓN
Efecto de los tratamientos de semilla, la procedencia y el riego en el establecimiento de Hibiscus elatus
Effect of seed treatments, provenance and irrigation on the establishment of Hibiscus elatus
Laura Montejo Valdés y J.A. Sánchez
Instituto de Ecología y Sistemática, Ministerio de Ciencia, Tecnología y Medio Ambiente,Carretera Varona, km 3½, Capdevila, Boyeros, C.P. 11900, La Habana 19, Cuba
E-mail: laura@ecologia.cu
RESUMEN
Se estudió la emergencia y el crecimiento de las plántulas de Hibiscus elatus en condiciones de vivero, en semillas sometidas a tratamientos de escarificación ácida en combinación o no con tratamientos de hidratación parcial en agua. Las semillas se colectaron en cuatro procedencias boscosas de la Reserva de la Biosfera Sierra del Rosario (dos bosques maduros siempreverdes estacionales y dos bosques secundarios) y se sometieron a diferentes frecuencias de riego (diario, días alternos y una vez por semana) durante su estancia en vivero. Los tratamientos pregerminativos fueron adecuados para incrementar la emergencia y el crecimiento de las plántulas en relación con el control (semillas no tratadas), para todas las procedencias, aunque el efecto positivo dependió del nivel de riego del sustrato. El estrés hídrico afectó de forma significativa el vigor de las plántulas, independientemente del tratamiento utilizado; sin embargo, bajo estas condiciones de siembra, el tratamiento hídrico resultó el más favorable para mejorar el funcionamiento de las plantas en las semillas colectadas en el bosque maduro; mientras que las del bosque secundario tuvieron una mejor habilidad para responder al estrés hídrico con el de escarificación, combinado con la hidratación parcial en agua. Se demostró que el empleo de tratamientos robustecedores a las semillas disminuyó el efecto dañino provocado por la insuficiente humedad del suelo sobre el establecimiento de las plántulas, independientemente del sitio de colecta.
Palabras claves: crecimiento, emergencia, Hibiscus elatus, hidratación.
ABSTRACT
The emergence and growth of Hibiscus elatus seedlings under nursery conditions was studied, in seeds subject to acid scarification treatments in combination or not- with partial hydration treatments in water. The seeds were collected in four forest provenances of the Sierra del Rosario Biosphere Reserve (two seasonal evergreen mature forests and two secondary forests) and were subject to different irrigation frequencies (daily, alternate days and once a week) during their permanence in nursery. The pregerminative treatments were adequate to increase seedling emergence and growth with regards to the control (untreated seeds), for all provenances, although the positive effect depended on the irrigation rate of the substratum. The water stress significantly affected seedling vigor, independently from the treatment used; nevertheless, under these planting conditions, the water treatment was the most favorable to improve plant functioning in the seeds collected in the mature forest; while those from the secondary forest had a better ability to respond to water stress with scarification treatment, combined with partial hydration in water. The use of hardening treatments in the seeds was proven to decrease the damaging effect caused by the insufficient soil moisture on seedling establishment, independently from the collection site.
Key words: emergence, growth, Hibiscus elatus, hydration.
Hibiscus elatus Sw., conocida hasta su reciente cambio de sinonimia como Talipariti elatum (Sw.) Fryxell (Malvaceae), es un árbol tropical pionero tardío de rápido crecimiento (Herrera et al., 1997). En Cuba, ocupa bosques húmedos, semideciduos y vegetación secundaria (Bisse, 1988) y ha sido ampliamente empleado en planes de reforestación y en la alimentación del ganado (Toral et al., 2001); aunque su germinación en condiciones de laboratorio y de vivero es pobre y errática, cuando no se aplican tratamientos pregerminativos (Álvarez, 1984; Sánchez et al., 2003). También la germinación y la emergencia de la plántula en esta especie se afecta considerablemente bajo condiciones controladas de estrés hídrico y calórico del sustrato (Sánchez et al., 2003). Según Sánchez (2007), la mayoría de sus semillas frescas presenta dormancia fisiológica no profunda y solo un 12% del lote muestra dormancia física o impermeabilidad de las cubiertas al agua. El primer tipo de dormancia señalado para esta especie es el más común para árboles de bosques tropicales húmedos, y el segundo es muy frecuente en especies de bosques secos (Sautu et al., 2007); ambos se eliminan mediante tratamientos pregerminativos de escarificación ácida o mecánica, según lo demostrado por Álvarez (1984), quien señaló, además, una gran variabilidad en la respuesta germinativa de la planta, acorde con la procedencia de las semillas.
En numerosas especies de interés agrícola se ha informado la utilidad de los tratamientos de hidratación-deshidratación, en combinación -o no- con otros tratamientos robustecedores para mejorar el funcionamiento de las semillas, las plántulas y el rendimiento agrícola bajo condiciones ecológicas extremas (Sánchez et al., 2001; 2007). Los resultados con dichos procedimientos son escasos en especies arbóreas tropicales, a pesar de los numerosos beneficios que brindan para asegurar el establecimiento de las plántulas; no obstante, se han realizado valiosas contribuciones, fundamentalmente en especies forestales pioneras y otras de gran importancia económica (Sánchez et al., 2004, 2006; Vargas y Kossmann, 2008; Brancalion et al., 2008, 2010).
En las semillas de H. elatus, talestratamientos se han aplicado para mejorar la germinación y la emergencia de las plántulas bajo condiciones controladas de estrés calórico, y también para revigorizar semillas envejecidas (Sánchez et al., 2003, 2004; Montejo et al., 2005); sin embargo, no se conocen los efectos de estos procedimientos fisiológicos en el establecimiento de la especie en condiciones de déficit de humedad del suelo. Por lo tanto, el objetivo del trabajo fue evaluar el efecto de diferentes tratamientos pregerminativos (escarificación ácida en combinación o no con tratamientos hídricos) en la emergencia de la semilla de diferentes procedencias; así como en el crecimiento y la supervivencia de las plántulas de H. elatus, sometidas a diferentes condiciones de riego durante su estancia en vivero.
MATERIALES Y MÉTODOS
Descripción general del sitio
El estudio se realizó en la Reserva de la Biosfera Sierra del Rosario (RBSR), Artemisa, Cuba (22º 45' N, 82º 50' O), que tiene una extensión de 25 000 ha y varios tipos de formaciones vegetales: boscosas, arbustivas y herbáceas; entre las más extendidas está el bosque siempreverde mesófilo (Herrera et al., 1988). No obstante, una parte importante de la superficie boscosa de la Reserva está cubierta por bosques secundarios con diferentes grados de perturbación, debido a la creación de terrazas, la apertura de caminos y la tala selectiva (Herrera et al., 1997). La temperatura media anual es de 24,4ºC y la precipitación promedio anual, de 2 013 mm; aunque en alturas mayores a los 400 m puede llegar hasta 2 300 mm (Herrera et al., 1997). Los suelos son variados y, según Hernández et al. (1988), predominan los Fersialíticos y los Pardos; y su fertilidad es muy variable, aunque tiende a ser media.
Los sitios de colecta de los frutos se seleccionaron según el grado de conservación de la vegetación. Estos abarcaron dos localidades de zonas boscosas de vegetación secundaria, conocidas como El Taburete (82º56'12", O,22º50'46"N) y Las Delicias (82º56' 11" O, 22º51'00" N), y en bosques siempreverdes mesófilos, en áreas llamadas Helechal (82º57' 43" O, 22º49' 56" N) y Cima del Vallecito (en lo adelante, Cima) (82º57' 52", O 22º 49'42" N), ubicadas dentro de la sección central de la RBSR, la cual se considera una de las áreas de mayor conservación de la estructura original de su vegetación (Herrera et al., 1997). Las semillas frescas de H. elatus se colectaron directamente de la planta madre, a partir de frutos maduros, mediante una vara telescópica o subiendo a los árboles. La colecta se realizó durante la principal época de fructificación de la especie en Cuba, a finales de la estación de seca, en el mes de marzo de 2008. Las semillas se mezclaron y se obtuvieron cuatro poblaciones de disemínulos, una de cada zona de colecta, y se depositaron en frascos de cristal con cierre hermético, a temperatura de 25oC en el laboratorio. Los ensayos de viveros se realizaron inmediatamente después de la colecta.
En los sitios de vegetación secundaria, los árboles de H. elatus tenían una altura entre 8 y 10 m y un diámetro no mayor de 20 cm; mientras que los que crecieron en bosques siempreverdes mesófilos, cuya vegetación era muy cerrada, alcanzaron una altura entre 15 y 25 m y un diámetro de 25 y 30 cm. También la composición de la vegetación en las zonas de colecta fue distinta: en las áreas de vegetación secundaria había menos diversidad de especies arbóreas y predominaban las plantas introducidas, tales como: Syzygum jambos, Roystonea regia, Mangifera indica, Piper sp., Bambusa vulgaris y Tectona grandis; en los sitios de bosque maduro, prevalecía una vegetación típica de lugares húmedos de la reserva y abundaban: Prunus occidentalis, Alchornea latifolia, Ardisia dentata, Ficus sp., Zanthoxilum martinicens y Psycotria horizontalis; aunque algunos taxones, como Calophyllum brasiliense, aparecieron en todos los sitios de colecta. La temperatura del suelo dentro de cada vegetación varió durante la época de producción de semilla: en las áreas menos conservadas, la temperatura máxima y mínima promedio oscilaron entre 25,6 y 18,6ºC, y en los bosques siempreverdes, entre 23,2 y 17,1ºC, respectivamente.
Tratamientos pregerminativos y condiciones de siembra
Las semillas se sometieron a las combinaciones de tratamientos pregerminativos (de escarificación y de hidratación parcial) que resultaron mejores en esta especie, bajo condiciones controladas de estrés calórico, según lo informado por Montejo et al. (2005). Los tratamientos utilizados fueron: semillas no tratadas o control (T1); semillas sometidas a escarificación ácida con H2SO4 al 96%, durante 20', y lavadas posteriormente con agua corriente, durante 10' (T2); semillas intactas, que se hidrataron con agua hasta aproximadamente el final de la fase I del patrón trifásico de absorción de agua (T3); y semillas sometidas a la combinación de los tratamientos T2 y T3, es decir, escarificación ácida más hidratación parcial en agua (T4). La fase de hidratación se realizó a temperatura alterna de 25/35ºC, durante 72 h para el T3 y 7 h para el T4. La deshidratación de las semillas ocurrió a 25 ± 1ºC (45% de humedad relativa) durante 48 h, hasta alcanzar, aproximadamente, el contenido de humedad inicial (13-15% con base en el peso fresco).
La siembra se realizó en condiciones de vivero, en áreas del Instituto de Ecología y Sistemática La Habana, Cuba (23º01'N, 82º21'O). Las condiciones experimentales fueron: temperatura ambiente (entre 20,7ºC y 24,1ºC) y luz natural (aproximadamente entre 30 y 40% de la radiación fotosintéticamente activa). El experimento se efectúo en la estación poco lluviosa, entre el 28 de noviembre de 2008 y el 10 de febrero de 2009; durante su realización no ocurrieron precipitaciones en el área de estudio. El riego se aplicó a capacidad de campo y se establecieron tres frecuencias: riego diario (R-I), riego dos veces por semana (R-II) y riego una vez por semana (R-III). No se aplicaron fertilizantes ni plaguicidas.
Se utilizó suelo de la RBSR, del tipo Fersialitico Pardo Rojizo (Hernández et al., 1988), que es uno de los más extendidos en los sitios de colecta de las semillas y en otras formaciones vegetales de la RBSR. Este se colectó a una profundidad de 0-10 cm y se le retiró la materia orgánica en descomposición (resto de hojarascas y raíces). Sus características químicas fueron: pH (H20): 6,0; materia orgánica oxidable: 5,1%, según Walkey-Black; fósforo asimilable: 10,4 µg g-1, según Machiguin; capacidad de intercambio catiónico (suma del Na, K, Ca y Mg): 15,15 cmol kg-1, por absorción atómica; y nitrógeno total: 0,26%, por micro Kjeldahl. La textura era loam arcillosa y la humedad en saturación fue de 36,7% (determinada por gravimetría).
Las condiciones microambientales durante el periodo del experimento fueron las siguientes: temperatura media del suelo 24,1ºC (máxima 29,0ºC y mínima 19,2ºC) y humedad relativa de 79%.
Las semillas control (T1) y las sometidas a T2, T3 y T4 se sembraron en el suelo contenido en potes plásticos, blancos, de 25,8 cm de ancho por 31,5 cm de altura y 5 dm³ de capacidad, a una profundidad de 2 cm; en cada pote se colocaron cinco semillas. Para cada procedencia se estableció un diseño de bloques al azar con tres repeticiones de 240 potes cada una. A su vez, dentro de cada bloque se establecieron, de forma aleatoria, los tres niveles de riego para todos los tratamientos pregerminativos ensayados. En total, por procedencia se emplearon 720 potes, y por tratamiento (riego por tratamiento pregerminativo), 60 réplicas.
El conteo de la emergencia se realizó diariamente, durante 30 días, a partir del comienzo del experimento. Se determinó el día de inicio de la emergencia y el porcentaje de emergencia final de las plántulas. Los potes se rotaron periódicamente hasta el final del experimento, para eliminar los efectos causados por la posición, y las plántulas se regaron de manera diferenciada a partir de la segunda semana después de la siembra. Posterior a los 30 días de esta (a los 15 días de iniciarse la emergencia) se practicó el raleo de las plántulas para dejar un individuo por pote, y se seleccionaron al azar 15 individuos por tratamiento, para determinar el vigor de las plántulas (en este momento todas tenían, al menos, dos hojas verdaderas, además de los cotiledones). La obtención de las plántulas intactas se logró por inmersión de cada pote en agua corriente, después se lavaron cuidadosamente las raíces para eliminar la tierra. Se determinó, como variable del vigor, la masa seca (g) de las raíces, del tallo y de las demás partes aéreas (hojas), mediante el secado de estas en una estufa a 70ºC, durante 48 h.
Después del entresaque de las plántulas, las que quedaron por cada pote permanecieron en las mismas condiciones de riego ya descritas. El experimento se mantuvo por 40 días más, con el objetivo de evaluar el crecimiento de las plantas hasta los 60 días de su estancia en el vivero. Se escogieron al azar 15 plántulas por tratamiento y procedencia. Las hojas, el tallo y las raíces se secaron separadamente, a 70ºC por 48 h, y después se pesaron. La obtención total de las plantas se realizó con agua corriente.
Sobre la base de los datos de biomasa seca, se calculó la biomasa seca total (BS total) en gramos (g) y las fracciones de distribución de biomasa (g g-1) a las hojas (FM hojas), el tallo (FM tallo) y la raíz (FM raíz total), en relación con el peso total de la plántula. Además, se cálculo la relación biomasa seca subterránea/biomasa aérea total (g g-1) (FB subterránea/aérea). Las plántulas se examinaron diariamente, para evaluar la supervivencia al final del experimento. A partir de la biomasa seca total de la planta, se estimó la tasa de crecimiento relativo (TCR, g g-1día-1) después de los 60 días de permanencia en el vivero; como biomasa a un tiempo inicial se utilizó la producción de materia seca en los primeros 30 días del experimento. Esta variable se estableció de acuerdo con Poorter y Garnier (2007), según la fórmula: TCR = ln P2 - ln P1/ t2 - t1. Donde: ln: logaritmo neperiano; P1: peso seco de la plántula a un tiempo inicial; P2: peso seco a un tiempo final; t1: tiempo inicial; y t2: tiempo final. Este índice muestra una medida de la producción de materia seca por unidad de biomasa y tiempo. Para el cálculo de la TCR primeramente se determinó los logaritmos de los pesos individuales de las plantas, y después, la media de esos logaritmos.
Se calculó la media y el error estándar de cada variable de emergencia y crecimiento estudiada. La normalidad de los datos se verificó mediante la prueba de bondad de ajuste de Kolmogorov-Smirnov, y la homogeneidad de varianza a través de la prueba de Bartlett. Los datos expresados en porcentaje se transformaron en arcsen√%. Las variables de emergencia y de vigor de las plántulas se procesaron mediante el análisis de varianza (ANOVA) de clasificación simple, con arreglo factorial de los tratamientos. No se aplicaron pruebas de comparaciones múltiples de medias a posteriori, debido a que las variables independientes representan combinaciones de tratamiento cualitativo no estructurado (tratamiento pregerminativo y procedencia seminal), con tratamiento cuantitativo (cantidad de agua adicionada, o sea, tipo de riego) (Blanco, 2001). A partir de una matriz de correlación, se aplicó el análisis de componentes principales (ACP) para determinar cómo se interrelacionaron y agruparon las procedencias de H. elatus, según los tratamientos pregerminativos y las condiciones de estrés hídrico (con y sin estrés). En este último análisis se utilizaron los valores individuales de cada muestra o réplica por tratamiento.
Las semillas colectadas en los sitios de bosques de vegetación secundaria (El Taburete y Las Delicias) presentaron una viabilidad inicial (porcentaje de germinación final a la temperatura óptima de la especie) de 91%; mientras que en los disemínulos provenientes de Helechal y de Cima (bosques siempreverdes mesófilos) fue de 89 y 69%, respectivamente. El contenido inicial de humedad de los lotes varió entre 13,0 y 15,7%, y las poblaciones del bosque maduro tuvieron una mayor humedad. La masa seca promedio de las semillas, determinada mediante el secado durante 17 h a temperatura constante de 103 ± 2ºC (ISTA, 1999), en El Taburete y Las Delicias fue de 0,0197 g (± 0,0009) y 0,0207 g (± 0,001), y en Helechal y Cima varió entre 0,0116 (± 0,0006) y 0,0139 g (± 0,0007), respectivamente.
El análisis de varianza indicó que el inicio de la emergencia de las plántulas no se afectó por la interacción de los factores probados (procedencia, frecuencia de riego y tratamiento pregerminativo), aunque los dos últimos efectos principales sí afectaron significativamente este indicador (tabla 1). En cambio, el porcentaje de emergencia final y la biomasa seca total de las plántulas de la primera cosecha dependieron, de forma significativa, de la interacción de los tres factores.
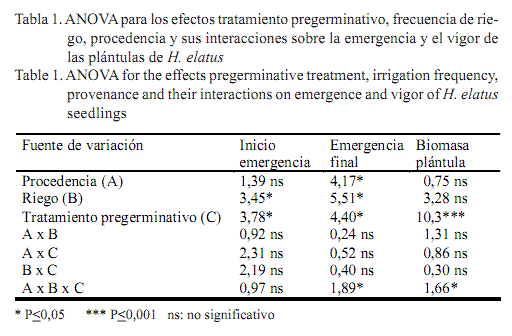
En las semillas procedentes de El Taburete y Las Delicias, en cualquier condición de riego, los tratamientos pregerminativos resultaron adecuados para acelerar el inicio de la emergencia de la radícula, en relación con el control (tabla 2). El porcentaje de emergencia final en estos lotes, en condiciones de R-I, fue mayor en las semillas procedentes de T2 y T4; mientras que T3 no mostró diferencias con respecto al testigo. Por su parte, cuando se aplicó R-II, los tratamientos pregerminativos incrementaron la emergencia de las plántulas en relación con el control. Sin embargo, al disminuir aun más la periodicidad del riego (R-III), se afectó el porcentaje final de emergencia, principalmente en las semillas no tratadas y sometidas a T2 (tabla 2). Al parecer, la escarificación en H2SO4 bajo esta condición de déficit de humedad del sustrato provocó el deterioro de las semillas y la pérdida de viabilidad.
En estos lotes de semillas de bosques de vegetación secundaria, el T4 fue el más apropiado para incrementar el porcentaje de emergencia, en cualquier condición de riego. Con dicho tratamiento y con la menor humedad del sustrato, el inicio de la emergencia en El Taburete y Las Delicias ocurrió en menos tiempo (a los ocho y nueve días de la siembra) respecto al resto de los tratamientos, y el porcentaje de emergencia final fue de 52 y 64%, respectivamente. En cambio, en las semillas de T1 con igual condición de riego, la emergencia de las plántulas ocurrió a partir de los 18 días en El Taburete y a los 21 en Las Delicias, y solo se obtuvo al final del experimento un 28 y 12% de emergencia, respectivamente (tabla 2). Ello evidenció la efectividad de combinar los dos tratamientos en el establecimiento de dichos lotes. No obstante, a medida que disminuyó el riego el T3 fue igualmente adecuado para acelerar e incrementar la emergencia, en condiciones desfavorables para el establecimiento.
Por su parte, en los disemínulos colectados en Helechal y sometidos a riego diario, se alcanzaron los mayores porcentajes y velocidad de emergencia con el T2, respecto al T1. Sin embargo, con R-II y R-III se afectó la respuesta regenerativa del lote en dicho tratamiento, al disminuir drásticamente el número de plántulas emergidas (tabla 2). Por ello, en condiciones de riego en días alternos, los tratamientos hídricos (T3 y T4) fueron los idóneos para incrementar el número de plántulas, con los que se alcanzó más de un 50% de emergencia al final del ensayo; mientras que el control no superó el 20%. Por otra parte, bajo condiciones severas de déficit de humedad del suelo, solo el T4 fue efectivo para mejorar la capacidad de emergencia, al propiciar un incremento significativo en el número de plántulas producidas (tabla 2).
En las semillas colectadas en Cima, una vez que la periodicidad de riego disminuyó, ningún tratamiento logró acelerar el inicio de la emergencia en relación con el control, y únicamente T3 provocó un incremento del porcentaje de emergencia final, en cualquier condición de riego (tabla 2). El método robustecedor no fue adecuado, pues disminuyó el número final de plántulas en todas las condiciones de hidratación del sustrato. Al parecer, en este lote el de menor viabilidad inicial (69%) pero el más vigoroso, dado el alto porcentaje de emergencia en las semillas control cuando el riego fue diario T4 provocó un sobreacondicionamiento de las semillas, el cual agotó las reservas nutricionales, redujo el número de plántulas emergidas y retardó el inicio de la emergencia de la radícula. Resultados similares en semillas de hortalizas sometidas a hidratación parcial fueron obtenidos por Welbaum y Bradford (1991), quienes concluyeron que la efectividad de estos tratamientos depende del grado de maduración o poder germinativo inicial de los disemínulos en el momento de la colecta. También Sánchez et al. (2001) y Moradi-Dezfuli et al. (2008) plantearon que las características de la especie, la variedad o lote, la correcta estandarización de los tratamientos pregerminativos y las condiciones ecológicas determinan la efectividad de dichos tratamientos.
La biomasa seca total de las plántulas, cosechadas a los 30 días de iniciado el experimento, se afectó significativamente (P<0,001) debido a los tratamientos pregerminativos y la interacción de los factores (tabla 1). En semillas colectadas en El Taburete, en condiciones de riego diario y en días alternos, los mayores valores de biomasa total se alcanzaron con la aplicación de T2 y T4; mientras que en el suelo de menor humedad solo el T4 produjo plántulas de mayor vigor, con respecto al resto. Por su parte, en Las Delicias, en cualquier condición de riego, T3 y T4 produjeron las plántulas de mayor biomasa total (tabla 2). No obstante, solo cuando se provocó el déficit hídrico en ambos lotes se incrementaron las diferencias entre los tratamientos antes mencionados y el control (tabla 2). De hecho, en las plántulas de El Taburete, sometidas a riego en días alternos y procedentes de semillas escarificadas (T2) y acondicionadas (T4), se obtuvieron incrementos de la masa seca total de un 83% en relación con T1; mientras que en Las Delicias, cuando el riego disminuyó a una frecuencia, con los tratamientos hídricos mencionados se duplicaron los valores de la biomasa total con respecto a las semillas no tratadas (tabla 2).
En Helechal, con todas las frecuencias de riego, T3 fue el más efectivo, al producir plántulas con un mayor desarrollo y duplicar los incrementos en la masa total, en condiciones extremas de déficit de humedad del suelo en relación con el control (tabla 2); mientras que en las plántulas procedentes de Cima, dicho tratamiento solo fue el más idóneo cuando el riego comenzó a disminuir a dos veces y una vez por semana, al producir un crecimiento de la masa total de 95 y 89%, respectivamente. También con el empleo de T4 se obtuvieron resultados similares cuando el riego se aplicó diariamente y una vez por semana (tabla 2).
Este crecimiento en el vigor de la plántula con el empleo de los tratamientos hídricos, principalmente en un suelo sometido a baja irrigación, podría deberse a la capacidad que adquieren las plantas para evitar o disminuir el daño celular provocado por el estrés hídrico (Kozlowski y Pallardy, 2002). No obstante, estos resultados difieren de los obtenidos por Murungu y Madanza (2010), quienes consideraron que el acondicionamiento en semillas de trigo no tuvo efecto sobre el peso seco de la plántula.
Los resultados con el T2 fueron superiores que con el T1 en El Taburete, en R-I y R-II; en Las Delicias, en todas las frecuencias de riego; en el Helechal y la Cima, cuando el riego ocurrió en días alternos, en las plántulas de la Cima cuando se regó una vez por semana (tabla 2). T2, en estas condiciones, favoreció el desarrollo vegetativo de las plántulas. Resultados similares fueron obtenidos por Mabundza et al. (2010) en semillas de Passiflora edulis, con el empleo de tratamientos de escarificación en H2SO4, los cuales incrementaron la emergencia y el vigor de las plántulas. No obstante, cuando el riego disminuyó se obtuvieron los valores más bajos de esta variable en cualquier lote sometido a dicho tratamiento (tabla 2). Al parecer, la escarificación en H2SO4 bajo esta condición de déficit de humedad del sustrato afectó el vigor y el desarrollo vegetativo de la plántula.
Estudios previos, realizados por Montejo et al. (2005), demostraron que el empleo de tratamientos acondicionadores en las semillas, combinados con la escarificación ácida, acelera e incrementa la emergencia final en esta especie y produce plántulas con un mayor desarrollo vegetativo. Igualmente, en semillas de especies de árboles pioneros, Sánchez et al. (2003) obtuvieron resultados similares con la aplicación de tratamientos hídricos en combinación o no con el tratamiento robustecedor de choque térmico. Con este tratamiento, Sánchez et al. (2006) lograron plántulas de Trichospermum mexicanum con un mayor crecimiento que las procedentes de semillas testigo; igualmente a lo informado para otras plantas forestales pioneras como Guazuma ulmifolia, con el empleo de procedimientos hídricos y osmóticos (Brancalion et al., 2010), y para leguminosas arbóreas (Albizia lebbeck y Gliricidia sepium), en las que un ciclo de hidratación-deshidratación incrementó el establecimiento de las plántulas en vivero (González et al., 2009).
En plantas de interés agrícola, Sánchez et al. (2001) probaron con éxito estos métodos fisiológicos bajo estrés calórico, y Casenave y Tosenill (2007) lo hicieron en condiciones de sequía y salinidad del sustrato. Según Sánchez et al. (2003, 2007), las ventajas de los tratamientos robustecedores son más evidentes en condiciones de estrés, posiblemente como consecuencia de la estimulación de los mecanismos involucrados en evadir la deshidratación celular, los cuales inducen la tolerancia al estrés ambiental y la reparación de daños celulares (Farooq et al., 2009).
De acuerdo con lo anterior, Henckel (1982) concluyó que los cambios metabólicos y fisiológicos provocados por dichos tratamientos aceleran el desarrollo vegetativo de las plantas en sus primeras etapas de vida, a través de una mayor síntesis de proteína, DNA, RNA, así como un incremento en la actividad de la amilasa (Farooq et al., 2009); esta enzima está correlacionada positivamente con la movilización de reservas y la velocidad de la germinación (Basra et al., 2005), la cual incide directamente en el incremento del establecimiento y crecimiento de la plántula.
La biomasa seca total al final del experimento mostró diferencias significativas, debido a la interacción entre la procedencia y el riego, y entre la procedencia y el tratamiento pregerminativo (tabla 3). Por su parte, la distribución de biomasa a las hojas y a la raíz total solo se afectó debido a la frecuencia de riego, lo cual no ocurrió en el tallo, que mostró diferencias significativas en la interacción procedencia-riego, y riego-tratamiento pregerminativo (tabla 3).
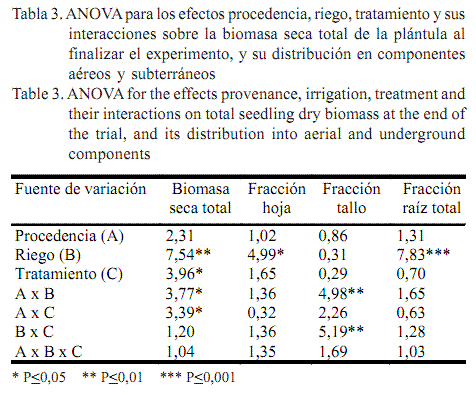
En general, en todos los lotes, la mayor cantidad de biomasa seca se produjo en las plantas que crecieron con el nivel óptimo de humedad del sustrato (tabla 4). En condiciones de R-I, los valores de masa seca total de todas las procedencias variaron entre 0,45 y 1,79 g. No obstante, este indicador tuvo una tendencia a disminuir en la medida que se redujo la frecuencia de riego, independientemente del sitio de colecta: en el sustrato con R-II los valores de biomasa estuvieron entre 0,26 y 1,73 g, y en R-III, entre 0,24 y 0,63 g (tabla 4).
La máxima producción de biomasa de las plántulas se alcanzó en El Taburete, independientemente del nivel de humedad del sustrato y el tratamiento, con excepción de las plántulas del Helechal procedentes del T3, y las de la Cima sometidas al T2 en condiciones de riego diario. Las plántulas control del Helechal humedecidas en días alternos, y las procedentes de semillas escarificadas de la Cima y de semillas acondicionadas del Helechal, en condiciones extremas de déficit de humedad del suelo, alcanzaron el mayor peso total en relación con el resto de las procedencias. Por su parte, el menor valor de esta variable lo presentaron las plántulas de Las Delicias provenientes del T2, cuando el riego se aplicó una vez por semana (tabla 4).
También Sánchez (2007) obtuvo en esta especie y en otras plantas arbóreas pioneras, y de otros estadios sucesionales, los mayores valores de biomasa seca al incrementarse el nivel de nutrientes del sustrato. Como es conocido, esta planta ocupa preferentemente ecosistemas húmedos o semihúmedos (Herrera et al., 1997), donde su crecimiento está influenciado por la presencia de nutrientes y agua en el suelo; aunque puede encontrarse en ambientes con cierto grado de aridez, debido quizá a su gran plasticidad fenotípica, que le permite germinar y establecerse en sitios más heterogéneos en cuanto a recursos nutrimentales e hídricos (Sánchez, 2007).
Los tratamientos pregerminativos de hidratación-deshidratación tuvieron un efecto significativo sobre el crecimiento de las plántulas, y la repuesta dependió de la procedencia seminal (tabla 4). Con la aplicación de T3 y T4 se obtuvieron los mayores valores de biomasa total en las plántulas de semillas colectadas en el bosque secundario (El Taburete y Las Delicias). El T4 fue el más idóneo en cualquier condición de riego, al propiciar una mayor acumulación de biomasa absoluta tanto en las hojas como en el tallo y las raíces, que varió según el riego (datos no mostrados) en relación con las plántulas sin tratamiento.
Sin embargo, el comportamiento de las semillas procedentes del bosque siempreverde fue diferente. En Helechal la biomasa total solo se incrementó considerablemente con la aplicación del T3, en cualquier condición de riego; mientras que el resto de los tratamientos resultaron perjudiciales para el crecimiento, cuando el riego se realizó diariamente y en días alternos (tabla 4). La escarificación en ácido afectó la respuesta reproductiva de esta procedencia, aún más cuando se combinó con el tratamiento hídrico, pues provocó una disminución en el crecimiento de las plántulas. Sin embargo, en condiciones extremas de sequía, en T2, T3 y T4 se produjeron incrementos de la biomasa total de 0,14; 0,32 y 0,18%, respectivamente, respecto a las plántulas no tratadas. Del mismo modo, Sánchez et al. (2007), en Leucaena leucocephala, y Basra et al. (2005), en semillas de arroz, lograron efectos beneficiosos del tratamiento robustecedor en el desarrollo vegetativo de la planta. Estos autores atribuyeron dichos resultados a los procesos fisiológicos estimulados por la alternancia de la hidratación y la deshidratación de la semilla. En este sentido, Halmer (2004) consideró que la respuesta de la semilla al acondicionamiento fue el incremento del rendimiento y la calidad de la cosecha, principalmente bajo condiciones subóptimas y de estrés para la especie.
En las semillas procedentes de la Cima, sometidas a riego diario, todos los tratamientos incrementaron la biomasa final en relación con el control (tabla 4). Sin embargo, cuando disminuyó la frecuencia de riego (R-II), solo T3 fue adecuado para incrementar dicha variable; mientras que en R-III, T3 y T2 superaron el valor final de biomasa total de las plántulas con respecto al testigo, aunque se afectó significativamente la supervivencia del lote. Al parecer, el método hídrico provocó el robustecimiento de las plántulas, las cuales son más resistentes a la deshidratación y poseen sistemas regenerativos mejor protegidos contra daños ambientales (Henckel, 1982).
Estos resultados coinciden con los obtenidos por Murungu et al. (2003), quienes emplearon tratamientos pregerminativos en cultivos de maíz y algodón en suelos secos, y corroboraron que el acondicionamiento mejoró el crecimiento temprano en suelos sin irrigación, por lo que las diferencias en las condiciones del suelo también podrían explicar la respuesta diferente.
En general, en esta especie el riego tuvo un efecto marcado en el crecimiento de las plantas, al incidir significativamente en la producción de hojas (P<0,05) y de raíz total (P<0,001) (tabla 3). Cuando el riego fue diario, todas las procedencias distribuyeron más biomasa a las hojas que a la raíz, independientemente del tratamiento; de manera similar sucedió cuando la frecuencia de riego fue en días alternos, con excepción de las plántulas control de Las Delicias y Helechal. Sin embargo, cuando el riego se aplicó una vez por semana hubo una disminución en la producción de masa seca destinada a las hojas en todos los lotes (tratados o no), a expensas de un incremento en la biomasa de la raíz total, excepto en las plántulas control de Las Delicias (donde se favoreció la producción de hojas), en relación con los rendimientos de las plantas en condiciones de riego diario (tabla 4). No obstante, cuando se regó una vez por semana, solo las plántulas control de El Taburete sometidas a T3 y T4, y también las de Las Delicias con dichos tratamientos, distribuyeron más biomasa a la raíz que a las hojas; mientras que en el lote del Helechal T2 y T4 propiciaron un mayor crecimiento de la raíz respecto a las hojas, a diferencia de la Cima donde únicamente T3 propició un incremento de esta variable (biomasa a la raíz), dado el poco aumento alcanzado con T2 (tabla 4).
En general, la máxima producción de hojas y raíces se alcanzó en las semillas sometidas a tratamientos pregerminativos, en relación con las no tratadas; excepto en Las Delicias y Cima, en sustratos bien hidratados, donde se logró los valores máximos de producción de raíces en semillas sin tratamiento previo, al igual que en Helechal (T1), en un suelo con nivel de humedad medio.
Por su parte, la biomasa destinada al tallo solo dependió, significativamente, de la interacción que se estableció entre procedencia y riego (tabla 3). No obstante, en condiciones de riego diario, las plántulas de El Taburete procedentes de T2 y las plántulas control de Cima distribuyeron el máximo valor de biomasa al tallo (0,17 y 0,18 g, respectivamente); mientras que en suelos humedecidos en días alternos, las plántulas testigo de Las Delicias y Cima alcanzaron valores entre 0,20 y 0,22 g, respectivamente. Sin embargo, en condiciones severas de sequía, solo las del Helechal sometidas a T4 lograron los más altos valores de esta variable; en El Taburete y la Cima, con igual nivel de humedad (R-III), los resultados difirieron independientemente del tratamiento (tabla 4). Por su parte, en el resto de las poblaciones no hubo diferencias significativas entre los tratamientos (tabla 4). Villar et al. (2004), al estudiar el crecimiento en 24 especies leñosas, concluyeron que, a diferencia de la compensación en la distribución de la biomasa a la parte aérea y la subterránea, la proporción que se destina al tallo no parece estar relacionada con el crecimiento de la planta.
En general, la relación biomasa seca subterránea/biomasa seca aérea, en todas las procedencias y en condiciones de riego diario y en días alternos, mostró tendencia a una mayor producción de biomasa aérea (tallo más hojas) que hacia las raíces, con independencia del tratamiento (tabla 4); ello pudo ser una vía para asegurar un rápido crecimiento, y con esto escapar de las fluctuaciones del ambiente en que se desarrollan las especies forestales pioneras tropicales (Bazzaz, 1996; Sánchez et al., 2003). No obstante, independientemente del sitio de colecta de las semillas, cuando hubo disminución extrema del riego se observó un incremento de esta variable en relación con las plántulas sometidas a riego diario, debido a una mayor producción de raíces secundarias en todas las procedencias estudiadas (datos no mostrados), provenientes del tratamiento acondicionador (T3), como ocurrió en Las Delicias, donde las semillas sometidas a T3 alcanzaron valores por encima de 1,0 (tabla 4). Esto permitió una mayor superficie de absorción para captar el agua y los nutrientes del suelo y, por lo tanto, una mayor sobrevivencia en un hábitat donde estos recursos son limitados (Villar et al., 2004).
Este resultado coincide con lo expresado por Vilagrosa et al. (2003), quienes señalaron que, a medida que avanza el estrés hídrico, hay una disminución de la biomasa foliar como estrategia de la planta para evitar la pérdida del individuo. En este sentido, Henckel (1982) concluyó que, en condiciones de sequía, las plántulas robustecidas tienden a distribuir mayor cantidad de biomasa a la raíz, que aquellas que proceden de semillas no tratadas.
La TCR tendió a disminuir con el nivel de humedad del sustrato (tabla 4). En general, todas las poblaciones alcanzaron los mayores valores de velocidad de crecimiento en suelos sometidos a una mayor irrigación: con riego diario los valores de TCR variaron de 0,07 hasta a 0,11 mg mg-1día-1; con riego intermedio, entre 0,05 y 0,11 mg mg-1día-1; y en condiciones de menor humedad, entre 0,05 y 0,08 mg mg-1día-1. Las plántulas de El Taburete procedentes de semillas acondicionadas (T3 y T4) tuvieron los valores máximos de velocidad de crecimiento en cualquier frecuencia de riego, lo que permitió que alcanzaran una mayor biomasa total en menos tiempo, en relación con el resto de las procedencias (tabla 4). Villar et al. (2004) afirmaron que en las plantas se manifiesta una estrecha relación entre una mayor asignación de biomasa a las hojas y, en consecuencia, una mayor velocidad de crecimiento; o bien una mayor asignación de biomasa a la raíz, con lo que se consigue una mayor absorción de agua y nutrientes del suelo, pero a expensas de un menor crecimiento. Esto sucedió en las plántulas de Las Delicias en los suelos de menor humedad, las cuales mostraron una mayor asignación de masa a la raíz (fracción biomasa subterránea/biomasa aérea > 1) y una menor TCR (tabla 4).
Tal efecto coincide con experiencias anteriores sobre la variación de la velocidad de crecimiento relativo de las plántulas, en función de las condiciones del sustrato en plantas arbóreas tropicales (Sánchez, 2007) y también en especies leñosas del mediterráneo (Antúnez et al., 2001).
La supervivencia de las plántulas al finalizar el experimento (40 días después de la primera cosecha), cuando el riego se efectuó diariamente, fue máxima para las de El Taburete, Las Delicias y el Helechal, y de un 80% para las de la Cima. Sin embargo, esta variable comenzó a disminuir con la frecuencia de riego y la aplicación del tratamiento pregerminativo. En este sentido, cuando el riego se realizó en días alternos en las plántulas procedentes del T2, en El Taburete, Las Delicias y el Helechal se afectó la supervivencia en un 20, 40 y 20%, respectivamente (tabla 4). También se obtuvo un incremento significativo de la mortalidad cuando el riego se realizó una vez por semana en los lotes de Las Delicias, Helechal y la Cima, sometidos a T2 (tabla 4). Al parecer, la escarificación ácida de las semillas pudo haber afectado de forma significativa la integridad de la cubierta seminal, principalmente cuando las condiciones ambientales fueron desfavorables para el establecimiento.
T3 y T4 mantuvieron un alto porcentaje de sobrevivencia en todas las poblaciones, independientemente de la periodicidad del riego, excepto en la Cima, donde intervinieron de manera negativa cuando las plántulas crecieron en sustrato de poca disponibilidad de agua, al afectar en un 40% este indicador. Ello corrobora el sobreacondicionamiento que sufrieron las semillas ante dichos tratamientos fisiológicos (tabla 4).
En ambas condiciones de riego del sustrato, el ACP concentró más del 70% de la variación total de los datos originales en los dos primeros componentes principales (figs. 1A y B). Las variables que más contribuyeron a la formación del primer eje fueron la FM hojas, la FM raíz total, la relación masa subterránea/parte aérea (S/A) y el porcentaje de emergencia final de las plántulas (PEF), que presentaron un coeficiente de correlación lineal (valor absoluto) mayor que 0,80. Ello evidenció que dichos rasgos tuvieron una mayor variabilidad entre las procedencias seminales y después de tratar y sembrar las semillas, en diferentes condiciones de riego. Por consiguiente, estas variables definieron la ubicación de los individuos de H. elatus en el espacio bidimensional del ACP. También en ambas condiciones de siembra se apreció que la FM hojas se asoció de forma negativa con las variables que representaron la producción de biomasa subterránea (FM raíz total y S/A); sin embargo, en las condiciones de estrés hídrico (fig. 1B), el PEF se asoció negativamente con la FM hojas, lo que demostró que las semillas con mayor capacidad de emergencia de plántulas también produjeron individuos con mayor desarrollo del sistema radical.
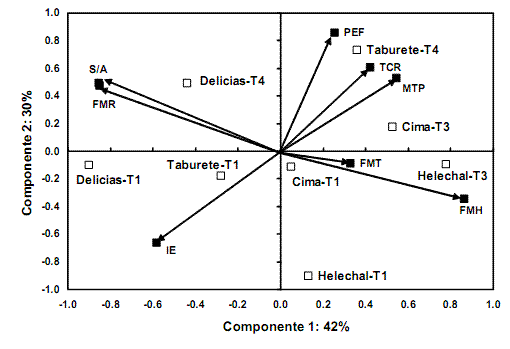
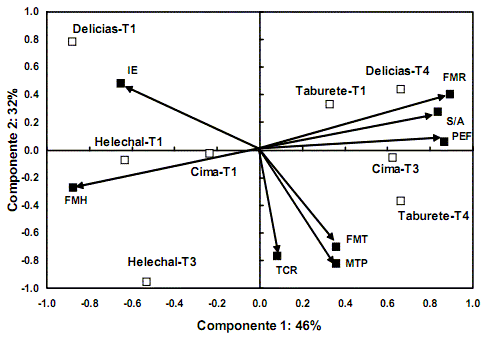


Las variables que más contribuyeron a la formación del segundo componente principal fueron la masa total de plántula (MTP), la velocidad de crecimiento (TCR) y el inicio de la emergencia (IE). En condiciones de riego normal, las dos primeras variables estuvieron asociadas positivamente entre sí y con el eje 2, y negativamente con el IE. Sin embargo, bajo esta condición de siembra, las semillas que presentaron los mayores valores de emergencia final de las plántulas también presentaron la mayor velocidad de crecimiento y el mayor valor absoluto de producción de biomasa. En este análisis, la FM tallo tuvo muy poca variabilidad entre procedencias y tratamientos pregerminativos; por consiguiente, aportó muy poco a la formación de los dos primeros componentes principales. Esto cambió cuando el análisis se realizó bajo condiciones de estrés hídrico: la FM tallo se correlacionó negativamente con la MTP, la TCR y el segundo componente principal. En esta condición de siembra, el IE apareció débilmente correlacionado con el eje 2, y tuvo una correlación negativa con el porcentaje de emergencia final en ambas condiciones de siembra, al incrementarse el número de plántulas disminuyó el tiempo necesario para realizar este proceso.
Igualmente, la ubicación de las procedencias en el espacio bidimensional del ACP evidenció que tuvieron patrones de regeneración distintos, aun en condiciones de riego adecuado (fig. 1A). Las procedencias de los bosques siempreverdes (Cima y Helechal) tendieron a ubicarse donde se incrementaron los valores de FM hojas, TCR y MTP, sobre todo aquellas con tratamientos pregerminativos; aunque la procedencia de El Taburete con tratamiento robustecedor (T4) se ubicó donde se obtuvieron los máximos valores de TCR, MTP y PEF. Esta última combinación de tratamiento bajo condiciones de estrés hídrico (fig. 1B) se agrupó hacia las variables antes mencionadas, pero también tendió a ubicarse donde se obtuvieron los máximos valores de FM raíz, S/A y FM tallo. Por otra parte, las procedencias de Las Delicias se separaron más, lo que demostró que los tratamientos robustecedores indujeron diferentes potenciales para adquirir tolerancia al estrés hídrico durante la emergencia de las plántulas y su posterior crecimiento. Es de señalar que estas pertenecen a un bosque secundario semiabierto, donde posiblemente las semillas se encuentren sometidas a condiciones de estrés hídrico y calórico, durante su estancia en el suelo y su posterior crecimiento como plántula.
En cambio, las procedencias del Helechal (T1 y T3) mantuvieron una ubicación bastante similar en las diferentes condiciones de siembra, aunque los tratamientos hídricos ocasionaron diferencias en su comportamiento regenerativo. De forma general, tendieron a producir los mayores valores de FM hojas, lo cual podría deberse a que están adaptadas a vivir en el ambiente del bosque siempreverde (sitio de colecta), donde la humedad del suelo no es una limitante para el crecimiento de las plantas, pero sí lo es la iluminación del sustrato; ello podría inducir cambios considerables en la producción foliar para aumentar la captación de la luz, en un ambiente lumínicamente heterogéneo, como puede ser bajo el dosel de la vegetación (Bazzaz, 1996; Herrera et al., 1997).
De acuerdo con los resultados de los análisis de ordenamiento, se evidenció que las distintas procedencias de H. elatus tuvieron un comportamiento regenerativo distinto, incluso aquellas que procedían de un tipo de vegetación similar, lo cual pudo deberse a su capacidad de adaptarse fácilmente a diferentes escenarios ambientales, tal y como se ha reportado en Cuba y otras regiones del trópico (Sánchez et al., 2003; Dalling et al., 2004). Este aspecto se reafirmó con la respuesta de las plántulas procedentes de las semillas tratadas en condiciones desfavorables de humedad del suelo. Ello se ha informado para un gran número de especies cultivadas (Sánchez et al., 2001; Nicasio-Arzeta et al., 2011), así como para especies arbóreas tropicales (Sánchez et al., 2003, 2006; Vargas y Kossmann, 2008; Brancalion et al., 2008, 2010).
Esta disimilitud en el comportamiento regenerativo de la especie podría estar relacionada, además, con la forma de fecundación de la planta. Se conoce que las flores de H. elatus son visitadas y polinizadas principalmente por murciélagos nectarívoros, los cuales son abundantes no solo en número, sino también en diversidad de especies (Mancina et al., 2007). Los murciélagos neotropicales pueden volar, diariamente, cientos de metros o varios kilómetros, desde sus refugios hasta los árboles florecidos (Mancina y García-Rivera, 2011), lo que permite el flujo genético entre procedencias y garantiza el éxito reproductivo de las plantas. En este sentido, Bazzaz (1996) señaló que las especies de estadios tempranos de la sucesión experimentan ambientes más heterogéneos que las de estadios más maduros; por tanto, las plantas pioneras suelen ser fenotípicamente más plásticas y tienen una amplia norma de reacción sobre una gran diversidad de ambientes.
Se confirmó que las semillas colectadas en bosques secundarios tuvieron una mejor habilidad para responder al estrés hídrico con el empleo de T4, a diferencia de las colectadas en bosques siempreverdes, las que se favorecieron con T3, en condiciones hídricas desfavorables. Por su parte, la Cima fue la población más susceptible a la poca irrigación y presentó una mayor mortalidad, aun cuando las semillas fueron robustecidas antes de la siembra. Sin embargo, en estas condiciones de estrés hídrico, una vez que la planta se establece destina una gran cantidad de biomasa a las raíces, posiblemente como estrategia de sobrevivencia para compensar la deficiencia de humedad del sustrato, lo cual incrementa la adquisición de este recurso.
En conclusión, quedó demostrado que H. elatus es sensible a la sequía, y que el empleo de tratamientos robustecedores a las semillas disminuyó el efecto dañino ocasionado por la insuficiente humedad del suelo en el establecimiento, el crecimiento y la supervivencia. Esta tecnología de bajos insumos fue efectiva para mejorar el funcionamiento reproductivo de la especie en condiciones ecológicas muy variadas (estrés hídrico), e indicó su plasticidad fenotípica en situaciones ambientales cambiantes.
Se agradece a Alejandro Gamboa por la asistencia técnica en el laboratorio y el vivero, y al proyecto "Ecofisiología de semillas y plántulas de árboles y arbustos de La Sierra del Rosario", del Programa Ramal de Diversidad Biológica, por el financiamiento de esta investigación. El autor J.A. Sánchez agradece a la Fundación Internacional para la Ciencia (IFS), por la ayuda ofrecida a través del donativo D/3536-2.
1. Álvarez, A. Las semillas de Hibiscus elatus Sw. (I). Comportamiento de las características que definen la calidad intrínseca de las semillas, atendiendo a diferentes fuentes productoras, edades y cosechas. Boletín Técnico Forestal. 84:1. 1984
2. Antúnez, I. et al. Relative growth rate in phylogenetically related deciduos and evergreen woody species. Oecologia. 128:172. 2001
3. Basra, S.M.A. et al. Physiological and biochemical aspects of seed vigor enhancement treatment in fine rice (Oriza sativa L.). Seed Sci. Technol. 33:623. 2005
4. Bazzaz, F.A. Plants in changing environments: linking physiological, population, and community ecology. Cambridge University Press, United Kingdom. 321 p. 1996
5. Bisse, J. Árboles de Cuba. Editorial Científico-Técnica, La Habana. 384 p. 1988
6. Blanco, F.A. Métodos apropiados de análisis estadístico subsiguiente al análisis de varianza (ANDEVA). Agronomía Costarricense. 25:53. 2001
7. Brancalion, P.H.S. et al. Priming of Mimosa bimucronata seeds: a tropical tree species from Brazil. Acta Horticulturae. 782:163. 2008
8. Brancalion, P.H.S. et al. Priming of pioneer tree Guazuma ulmifolia (Malvaceae) seeds evaluated by an automated computer image analysis. Scientia Agricola. 67:274. 2010
9. Casenave, E.C. & Toselli, M.E. Hydropriming as a pre-treatment for cotton germination under thermal and water stress conditions. Seed Sci. Technol. 35:88. 2007
10. Dalling, J.W. et al. Variation in growth the responses of neotropical pioneers to simulated forest gaps. Funct. Ecol. 18:725. 2004
11. Farooq, M. et al. Plant drought stress: effects, mechanisms and management. Agron. Sustain. Dev. 29:185. 2009
12. González, Yolanda et al. Efecto de los tratamientos de hidratación-deshidratación en la germinación, la emergencia y el vigor de las plántulas de Albizia lebbeck y Gliricidia sepium. Pastos y Forrajes. 32:255. 2009
13. Halmer, P. Methods to improve seed performance in the field. In: Handbook of seed physiology; application to agriculture. (Eds. R.L. Benech-Arnold and R.A. Sanchez). The Haworth Press, New York p. 125. 2004
14. Henckel, P.A. Physiology of heat and drought resistance in plants. Nauka, Moscow. 280 p. 1982
15. Hernández, A. et al. Los suelos de la Reserva. En: Ecología de los bosques siempreverdes de la Sierra del Rosario, Cuba. Proyecto MAB Nº 1, 1974-1987. (Eds. R.A. Herrera, Leda Menéndez, María Rodríguez y Elisa García).ROSTLAC, Montevideo. p. 88. 1988
16. Herrera, R.A. et al. Ecología de los bosques siempreverdes de la Sierra del Rosario, Cuba. Proyecto MAB No 1, 1974-1987. ROSTLAC, Montevideo. 760 p. 1988
17. Herrera, R.A. et al. 1997. Ecotechnologies for the sustainable management of tropical diversity. Nature and Resources. 33:2
18. ISTA. International rules for seed testing. Seed Sci. Technol. 27:155 (Suppl.). 1999
19. Kozlowski, T.T. & Pallardy, S.G. Acclimation and adaptive responses of woody plants to environmental stresses. Bot. Rev. 68:270. 2002
20. Mabundza, R.M. et al. Effects of different pre-germination treatment methods on the germination of passion (Passiflora edulis) seeds. J. Agric. Soc. Sci. 6:57. 2010
21. Mancina, C.A. & García-Rivera, Lainet. Murciélagos fitófagos. En: Mamíferos en Cuba. (Eds. R. Borroto-Páez y C.A. Mancina). UPC Print. Vaasa, Finlandia. p. 134. 2011
22. Mancina, C.A. et al. Habitat use by phyllostomid bat assemblages in secondary forest of the Sierra del Rosario Biosphere Reserve, Cuba. Acta Chiropterologica. 9:203. 2007
23. Montejo, Laura et al. Tratamientos pregerminativos de escarificación ácida y de hidratación parcial en la germinación y el vigor de Talipariti elatum. Pastos y Forrajes. 28:107. 2005
24. Moradi-Dezfuli, P. et al. Influence of priming techniques on seed germination behavior of maize inbred lines (Zea mays L.). J. Agric. Biol. Sci. 3:22. 2008
25. Murungu, F.S. & Madanza, T. Seed priming, genotype and sowing date effects on emergence, growth and yield of wheat in a tropical low altitude area of Zimbabwe. Afr. J. Agric. Res. 5:2341. 2010
26. Murungu, F.S. et al. Effects of seed priming, aggregate size and soil water potential on emergence of cotton (Gossypium hirsutum L.) and maize (Zea mays L.). Soil Till. Res. 74:161. 2003
27. Nicasio-Arzeta, S. et al. Effect of priming and saline substrate on germination and seedling growth of chalqueño maize (Zea mays). Agrociencia. 45:195. 2011
28. Poorter, H. & Garnier E. Ecological significance of inherent variation in relative growth rate and its components. In: Functional plant ecology. 2nd ed. (Eds. F.I. Pugnaire and F. Valladares). CRC Press, Taylor & Francis Group. London/New York. p. 724. 2007
29. Sánchez, J.A. Dormancy and nutrient contents in seeds and there with the establishment of tropical trees. Informe Final del Proyecto de la Fundación Internacional para la Ciencia (D/3536-1) Stockholm, Sweden. 97 p. 2007
30. Sánchez, J.A. et al. Tratamientos pregerminativos de hidratación-deshidratación de las semillas y sus efectos en plantas de interés agrícola. Agronomía Costarricense. 25:67. 2001
31. Sánchez, J.A. et al. Effects of seed hardening treatments on germination and establishment of pioneer trees under strees conditions. Ecotrópicos. 16:91. 2003
32. Sánchez, J.A. et al. Invigoration of pioneer tree seeds using prehydration treatments. Seed Sci. Technol. 32:355. 2004
33. Sánchez, J.A. et al. Tratamientos robustecedores de semillas para mejorar la emergencia y el crecimiento de Trichospermum mexicanum, árbol tropical pionero. Agronomía Costarricense. 30:7. 2006
34. Sánchez, J.A. et al. Enhanced germination, emergence and seedling vigour of Leucaena leucocephala using hardening hydration and acid shock treatments. Seed Sci. Technol. 35:224. 2007
35. Sautu, A. et al. Classification and ecological relationships of seed dormancy in a seasonal moist tropical forest, Panama, Central America. Seed Sci. Res. 17:127. 2007
36. Toral, Odalys et al. Colecta y potencialidades del germoplasma forrajero arbóreo en diferentes ecosistemas. Pastos y Forrajes. 24:105. 2001
37. Vargas, G.J. & Kossmann, I.D. Hydropriming of Parkia pendula [Penth. ex Walp.]: seeds with physical dormancy from Amazon tree. Revista Árvore.32:39. 2008
38. Vilagrosa, A.J. et al. Cavitation, stomatal conductance, and leaf dieback in seedling of two co-occurring Mediterranean shrubs during an intense drought. Journal of Experimental Botany. 54:2015. 2003
39. Villar, R. et al. Tasas de crecimiento en especies leñosas: aspectos funcionales e implicaciones ecológicas. En: Ecología del bosque mediterráneo en un mundo cambiante. (Ed. F. Valladares). Ministerio del Medio Ambiente, EGRAF, SA. Madrid. p. 191. 2004
40. Welbaum, G.E. & Bradford, K.J. Water relations of seed development and germination in Cucumis melo L.. VI. Influence of priming on germination responses to temperature and water potential during seed development. J. Exp. Bot. 42:393. 1991
Aceptado el 25 de junio del 2012

{kind=link}
{kind=link}