










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión On-line ISSN 2078-8452
Pastos y Forrajes vol.38 no.1 Matanzas ene.-mar. 2015
ARTÍCULO DE INVESTIGACIÓN
Efecto de la inoculación con bacterias rizosféricas y Trichoderma en trigo (Triticum aestivum L.)
Effect of the inoculation with rhizospheric bacteria and Trichoderma in wheat (Triticum aestivum L.)
C. J. Bécquer1, G. Lazarovits2, Laura Nielsen2, Maribel Quintana1, Modupe Adesina2, Laura Quigley2, I. Lalin2 y C. Ibbotson2
1 Estación Experimental de Sancti Spíritus, Apdo. 2255, Z. P. 1, C. P. 62200, Sancti Spíritus, Cuba. Correo electrónico: pastossp@enet.cu
2 Southern Crop Protection and Food Research Centre, London, Ontario, Canada (Agriculture and Agri-Food Canada)
RESUMEN
Se realizó un experimento en invernadero para evaluar en trigo el efecto de la inoculación con las rizobacterias Sinorhizobium meliloti, Azospirillum zeae y Azospirillum canadense, y con el hongo Trichoderma harzianum. El diseño experimental fue completamente aleatorizado, con cuatro réplicas y cinco plantas por tratamiento. Se utilizó un tratamiento fertilizado con NH4NO3 (150 ppm/kg de suelo), un control absoluto y 13 tratamientos inoculados. Se evaluó el peso seco aéreo, el peso seco radical, la longitud del tallo, la germinación y el contenido de clorofila foliar. La aplicación de Trichoderma por separado tuvo un efecto positivo en la longitud del tallo (5,58 cm); pero su presencia directa en algunas combinaciones con las bacterias rizosféricas no fue positiva ni mostró resultados uniformes en las variables en el momento de la siembra, o posterior a esta. Sin embargo, se destacó el tratamiento Trichoderma + NRG34-DS2 5d en el peso seco radical (0,45 g/planta) y en el peso seco aéreo (0,58 g/planta); así como el tratamiento A2-N7, en el contenido de clorofila (40,35 spads/planta) y la longitud del tallo (5,50 cm). Se concluye que la aplicación simple de Trichoderma no ejerció un efecto positivo en la mayoría de las variables ni los tratamientos influyeron positivamente en la germinación. Se recomienda evaluar Trichoderma + NRG34-DS2 5d y A2-N7 en experimentos de campo.
Palabras clave: Azospirillum, hongos, plantas, Sinorhizobium.
ABSTRACT
A trial was conducted in greenhouse to evaluate in wheat the effect of the inoculation with the rhizobacteria Sinorhizobium meliloti, Azospirillum zeae and Azospirillum canadense, and with the fungus Trichoderma harzianum. The experimental design was completely randomized, with four replications and five plants per treatment. A treatment fertilized with NH4NO3(150 ppm/kg of soil), an absolute control and 13 inoculated treatments, were used. The aerial dry weight, root dry weight, stem length, germination and content of leaf chlorophyll were evaluated. The separate application of Trichoderma had a positive effect on stem length (5,58 cm), but its presence in some combinations with rhizospheric bacteria was not positive and at the moment of sowing, or afterwards did not show uniform results in the variables. Nevertheless, the treatment Trichoderma + NRG34-DS2 5d stood out in the root dry weight (0,45 g/plant) and the aerial dry weight (0,58 g/plant); as well as the treatment A2-N7, in the chlorophyll content (40,35 SPADS/plant) and stem length (5,50 cm). It is concluded that the simple application of Trichoderma did not exert a positive effect on most variables, and none of the treatments positively influenced germination. It is recommended to evaluate Trichoderma + NRG34-DS2 5d and A2-N7 in field experiments.
Key words: Azospirillum, fungi, plants, Sinorhizobium.
INTRODUCCIÓN
La preocupación actual por los efectos colaterales de los agroquímicos ha motivado un interés creciente por aprender más sobre la cooperación entre las poblaciones microbianas de la rizosfera y la forma en que estas pudieran ser aplicadas en la agricultura (Barea et al., 2004).
Los rizobios se consideran, generalmente, como socios microbianos simbióticos de las leguminosas, a través de la formación de nódulos fijadores de nitrógeno (Antoun y Prévost, 2005). Sin embargo, estas bacterias pueden producir fitohormonas y solubilizar fosfatos orgánicos e inorgánicos, que desempeñan un papel importante en el crecimiento vegetal (Antoun et al., 1998).
Las especies pertenecientes al género Azospirillum también se consideran promotoras del crecimiento vegetal (Zahir et al., 2004). Estas bacterias, además de su actividad dinitrofijadora, tienen una producción significativa de fitohormonas, las cuales afectan la morfología radical y, por lo tanto, mejoran la toma de nutrientes del suelo.
Asimismo, el hongo filamentoso Trichoderma spp., que ejerce un antagonismo eficaz con fitopatógenos (Howell, 1998), induce mecanismos de defensa y estimula el crecimiento vegetal (Harman et al., 2004).
Existen informes previos sobre las ventajas de la coinoculación de plantas con bacterias rizosféricas y Trichoderma (Bécquer et al., 2004; Rudresh et al., 2005; Saber et al., 2009; Shaban y El-Bramawy, 2011). No obstante, aún quedan muchas interrogantes por resolver, como, por ejemplo, el modo en que estas combinaciones se pueden aplicar en dependencia del tipo de planta, el suelo y las condiciones ambientales, y qué especies microbianas son las más adecuadas en dichas combinaciones. Por ello, el objetivo de este experimento fue evaluar el efecto de la inoculación combinada de Sinorhizobium, Azospirillum y Trichoderma en trigo, en condiciones controladas y en diferentes momentos, para su futura aplicación en la práctica agrícola.
MATERIALES Y MÉTODOS
Procedencia de las cepas de Azospirillum y Sinorhizobium. Se utilizaron las cepas comerciales N7 y DS2, pertenecientes a Azospirillum zeae y Azospirillum canadense, respectivamente donadas por Agriculture and Agri-Food Canada (London, Ontario, Canadá); así como las cepas comerciales A2 y NRG34, pertenecientes a Sinorhizobium meliloti donadas por Agriculture and Agri-Food Canada (Crops Research and Development Centre, Québec, Canadá).
Procedencia de la cepa de Trichoderma. Se utilizó la cepa comercial TH-382 (Trichoderma harzianum), donada por Agriculture and Agri-Food Canada (Southern Crop Protection and Food Research Centre, Ontario, Canadá).
Variedad de trigo utilizada y procedencia. Se evaluó la variedad de trigo (Triticum aestivum L.) Pioneer 25R47, donada por A&L Canada Laboratories (Ontario, Canadá), y recomendada por esta institución para el tipo de suelo que se utilizó en el experimento (Lazarovits, G., com. pers.).
Procedimiento experimental
Crecimiento y aplicación de las cepas bacterianas. Las cepas crecieron en medio sólido levadura-manitol (Vincent, 1970) y se resuspendieron en medio líquido levadura-manitol, hasta lograr un título de 107-108 UFC/mL. El inóculo bacteriano se aplicó en el momento de la siembra (1,0 mL/semilla) o posteriormente (1,0 mL/planta) a esta, con dilución previa de 1:10 en una solución salina estéril de 0,85 % de NaCl. A los cinco días de la siembra se aplicó una segunda inoculación, con la misma concentración celular (Lupwayi et al., 2004).
Crecimiento y aplicación de la cepa fúngica. El inóculo fúngico se preparó en forma de suspensión conidial, al mezclar un cultivo que tenía 10 días de sembrado en PDA (papa dextrosa agar) con agua destilada y TWEEN-80 (0,01 %), para lo cual se raspó la superficie del medio con un agitador de vidrio estéril. El título final de la suspensión fue de 106-107 conidios/mL (Wolffhechell y Jensen, 1992). Las suspensiones conidiales se inocularon a las plantas de acuerdo con el tratamiento utilizado, de forma simple, o en combinación con los inóculos bacterianos.
Siembra y evaluación del experimento. Se utilizaron macetas plásticas que contenían 0,90 kg de suelo sin esterilizar, que se colectó en áreas pertenecientes a Agriculture and Agri-Food Canada, Estación Experimental Delhi (London, Ontario, Canadá) (42° 52 N, 80° 32 O). Este suelo se clasifica como Luvisol Marrón-Brunosólico Gris (Brunisolic Gray Brown Luvisol, Canadian System of Soil Classification) según lo informado por Chapman y Putman (1966) y se caracteriza por el predominio de arena (88 %), con la siguiente composición química: N: 0,01 %; MO: 1,0 %; P2O5: 217 ppm; K2O: 101,0 ppm, NaCl: 6,0 ppm, Mg: 115,0 ppm; Ca: 62,0 ppm; pH: 6,0. En cada maceta se sembraron nueve semillas, sin esterilizar. La evaluación se realizó a las tres semanas de la siembra, de acuerdo con lo recomendado por Somasegaran y Hoben (1994).
Diseño experimental y tratamientos. Se utilizó un diseño completamente aleatorizado, con 15 tratamientos y cuatro réplicas:
- Control fertilizado químicamente (NH4NO3: 150 ppm N/kg de suelo), según las recomendaciones de fertilización nitrogenada para este tipo de suelo (Lazarovits, G., com. pers.).
- A2-N7 (aplicación en el momento de la siembra).
- A2-DS2 (aplicación en el momento de la siembra).
- Trichoderma (aplicación en el momento de la siembra).
- A2-DS2-Trichoderma (aplicación de las tres cepas bacterianas en el momento de la siembra).
- A2-N7-Trichoderma (aplicación de las tres cepas en el momento de la siembra).
- NRG34-DS2-Trichoderma (aplicación de las tres cepas en el momento de la siembra).
- Trichoderma (aplicación en el momento de la siembra) + A2-DS2 5d. (aplicación de ambas cepas bacterianas a los cinco días posteriores a la siembra).
- Trichoderma (aplicación en momento de la siembra) + A2-N7 5d. (aplicación de ambas cepas bacterianas a los cinco días posteriores a la siembra).
- Trichoderma (aplicación en el momento de la siembra) + NRG34-DS2 5d. (aplicación de ambas cepas bacterianas a los cinco días posteriores a la siembra).
- NRG34-DS2 (aplicación de ambas cepas bacterianas en el momento de la siembra).
- A2-DS2 (aplicación de ambas cepas bacterianas en el momento de la siembra) + Trichoderma 5d. (aplicación del hongo a los cinco días posteriores a la siembra).
- A2-N7 (aplicación de ambas cepas bacterianas en el momento de la siembra) + Trichoderma 5d. (aplicación del hongo a los cinco días posteriores a la siembra).
- NRG34-DS2 (aplicación de ambas cepas bacterianas en el momento de la siembra) + Trichoderma 5d. (aplicación del hongo a los cinco días posteriores a la siembra).
- Control absoluto.
Se utilizaron cinco plantas por réplica y se evaluaron las variables: peso seco aéreo (g/planta); peso seco radical (g/planta); longitud del tallo (cm/planta); germinación (%) y contenido de clorofila foliar, con un detector de clorofila MINOLTA SPAD-501 (Rodríguez-Mendoza et al., 1998).
Análisis estadístico. Se aplicó un análisis de varianza y las diferencias entre medias se hallaron mediante la dócima de comparación de Duncan (1955). Los datos porcentuales se transformaron mediante arcsen»x + 0,375 (Lerch, 1976). Se utilizó el programa estadístico Statgraphics plus 5.1 para Windows.
RESULTADOS Y DISCUSIÓN
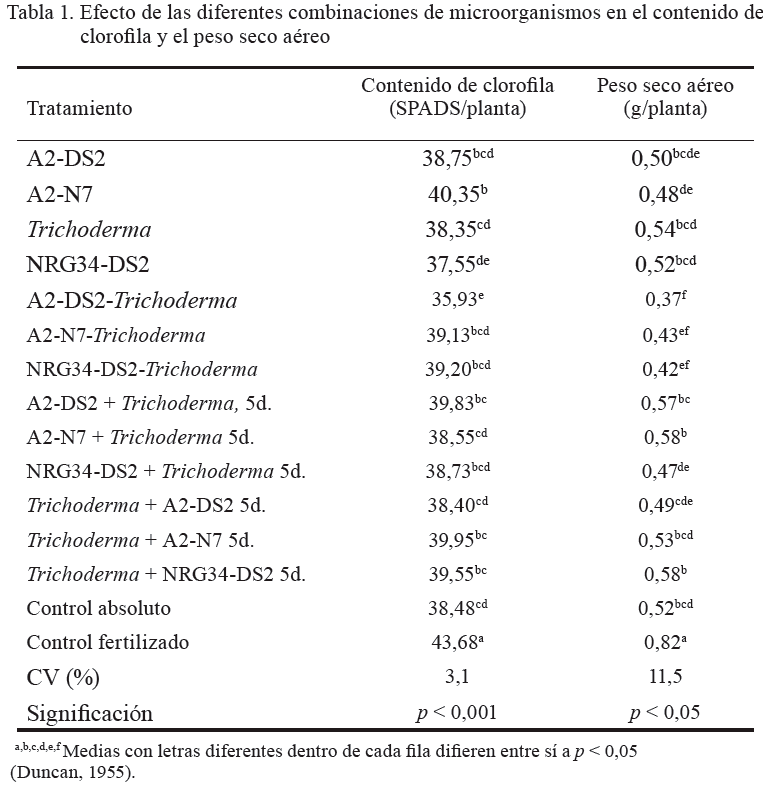
Contenido de clorofila
En cuanto al contenido de clorofila (tabla 1), el control fertilizado presentó el mayor valor (43,68 SPADS/planta, p < 0,001), en comparación con el resto de los tratamientos. Le siguió el tratamiento donde se combinaron las cepas A2 y N7, aplicadas en el momento de la siembra (40,35 SPADS/planta), el cual compartió superíndices comunes con otros tratamientos; pero resultó estadísticamente superior (p < 0,001) al control absoluto (38,48 SPADS/planta), así como a A2-N7 + Trichoderma 5d. (38,55 SPADS/planta), Trichoderma + A2-DS2 5d. (38,40 SPADS/planta), Trichoderma (38,35 SPADS/planta), NRG34-DS2. (37,55 SPADS/planta) y A2-DS2 + Trichoderma 5d. (35,93 SPADS/planta).
El hecho de que el tratamiento fertilizado presentara el mayor valor en esta variable es un indicio de la alta concentración de N inducida por la aplicación del fertilizante nitrogenado, ya que el contenido de clorofila es directamente proporcional al de N en la planta (Biswas et al., 2000). La combinación microbiana A2 (S. meliloti) y N7 (A. zeae), al momento de la siembra, fue la más efectiva. Según Askary et al. (2009), Sinorhizobium es un género altamente eficiente, debido a su efecto estimulador del crecimiento vegetal en trigo, en combinación con Azospirillum. En este sentido, Bécquer et al. (2012a) informaron que la combinación de A2 y N7 indujo el mayor contenido de clorofila, en esta especie.
La presencia de Trichoderma en las diferentes combinaciones no fue positiva, lo cual se pudiera deber a que este hongo, por su carácter antagonista con muchas bacterias (Harman et al., 2004), interfirió de forma discreta en el posible beneficio por el empleo de algunos de los tratamientos combinados, y ejerció un efecto depresor más palpable en tres de estos (A2-DS2 + Trichoderma 5d., Trichoderma + A2-DS2 5d., y A2-N7 + Trichoderma 5d.). Ello quizás dependió del grado de resistencia antifúngica de las especies bacterianas utilizadas en la combinación.
Peso seco aéreo
En el peso seco aéreo (tabla 1), independientemente de que el control fertilizado (0,82 g/planta) mostró valores estadísticamente superiores (p < 0,05) al resto de los tratamientos, se constató el efecto positivo de A2-N7 + Trichoderma 5d. (0,58 g/planta), así como de Trichoderma + NRG34-DS2 5d. (0,58 g/planta). Estos, aunque fueron estadísticamente superiores a Trichoderma + A2-DS2 5d., A2-N7, NRG34-DS2 + Trichoderma 5d., NRG34-DS2-Trichoderma, A2-N7-Trichoderma y A2-DS2-Trichoderma, no difirieron del control absoluto y de otros tratamientos, como A2-DS2. Tales resultados indican que el efecto positivo de dichas combinaciones en el incremento de la biomasa vegetal fue enmascarado por los microorganismos presentes en el suelo (no estéril) utilizado en el control absoluto, en el que la comunidad bacteriana autóctona de la rizosfera debió desempeñar un papel importante. Según Chelius y Triplett (2000), estas rizobacterias, al igual que las introducidas, se asocian a las raíces de la planta, debido a la influencia de los compuestos orgánicos provenientes de los exudados radicales. Asimismo, el hecho de que se igualaran estadísticamente los mejores tratamientos de la inoculación combinada con los del control absoluto corrobora, de cierta forma, la hipótesis de que las combinaciones de bacterias con Trichoderma se pueden afectar por la acción antagonista del hongo.
Longitud del tallo
En cuanto a la longitud del tallo (tabla 2), Trichoderma (5,58 cm), A2-N7 (5,50 cm) y NRG34-DS2 (5,50 cm) fueron estadísticamente superiores (p < 0,001) al control absoluto y a Trichoderma + A2-DS2 5d. (5,05 cm), NRG34-DS2 + Trichoderma 5d. (5,03 cm), NRG34-DS2-Trichoderma (4,95 cm), Trichoderma + A2-N7 5d. (4,80 cm), A2-N7-Trichoderma (4,73 cm) y A2-DS2-Trichoderma (4,68 cm); y no difirieron del resto de los tratamientos y del control fertilizado (5,18 cm).
Aunque el tratamiento inoculado solamente con Trichoderma presentó valores estadísticamente superiores a la mayoría de los tratamientos, los otros dos que le siguieron en importancia estaban compuestos solo por bacterias. Esto pudiera constituir una prueba palpable de la influencia antagonista del hongo, que se manifestó en determinadas variables agronómicas; a pesar de que su aplicación, por separado, ejerció un efecto positivo en la longitud del tallo. En este sentido, Avis et al. (2008) y Shoresh et al. (2010) informaron acerca de las propiedades biofertilizantes de Trichoderma, sobre la base del incremento de la absorción de minerales y su solubilización, así como en la producción de sustancias estimuladoras del crecimiento vegetal. Por otra parte, el antagonismo del hongo con alguna cepa en específico se puede deber a que este realiza su acción biocontroladora mediante la antibiosis, el micoparasitismo y la competencia por los nutrientes según lo planteado por Reino et al. (2008). Estos autores señalan que la mayoría de las cepas de Trichoderma producen metabolitos tóxicos volátiles y no volátiles que impiden la colonización de otros microorganismos antagonistas. Estos metabolitos fueron detectados por Saber et al. (2009), en T. harzianum. No se descarta que algunos de estos compuestos hayan influido negativamente en determinadas funciones de las bacterias que estimulan indicadores específicos del crecimiento vegetal, como las variables anteriormente descritas. Aunque Bécquer et al. (2001) encontraron que A. brasilense contrarrestó a T. harzianum en un estudio in vitro, Bécquer et al. (2013) determinaron que esta misma especie de hongo realizó actividad antagonista con A. canadense y A. zeae.
Peso seco radical
Con respecto al peso seco radical (tabla 2), Trichoderma + A2-DS2 5d. fue estadísticamente superior (p < 0,001) a: A2-N7-Trichoderma, A2-DS2-Trichoderma, NRG34-DS2, A2-N7, control absoluto, control fertilizado, NRG34-DS2 + Trichoderma 5d. y Trichoderma. Le siguió Trichoderma + NRG34-DS2 5d., que también fue estadísticamente superior al resto de los tratamientos, con excepción de A2-N7-Trichoderma.
En dicha variable influyeron positivamente dos tratamientos combinados que no se destacaron en las variables anteriores. El evidente antagonismo entre los microorganismos utilizados en este experimento no se evidenció en el peso seco radical, debido quizás a que Trichoderma, al emplear sus mecanismos de degradación celulolítica en las raíces del hospedero (Infante et al., 2009), permitió la entrada de las bacterias que fueron inoculadas a los cinco días posteriores a la aplicación del hongo, y lograron incidir directamente en el desarrollo radical (Sarig et al., 1992).
Es importante el hecho de que la aplicación simple de Trichoderma no ejerciera ningún efecto positivo en las raíces; aunque Gravel et al. (2006) y Chacón et al. (2007) observaron un aumento del peso seco radical en tomate y tabaco, respectivamente, al inocularlos con Trichoderma. Por otra parte, Benítez et al. (2004) consideraron que la colonización de las raíces por el hongo, independientemente de su género y especie, conlleva la producción de metabolitos tóxicos (fitoalexinas, flavonoides y terpenoides, derivados fenólicos y otros compuestos antimicrobianos) por parte de la planta, como respuesta a la invasión fúngica. La capacidad de sobrevivir a la acción de dichos compuestos debe variar de acuerdo con la cepa utilizada (Harman et al., 2004).
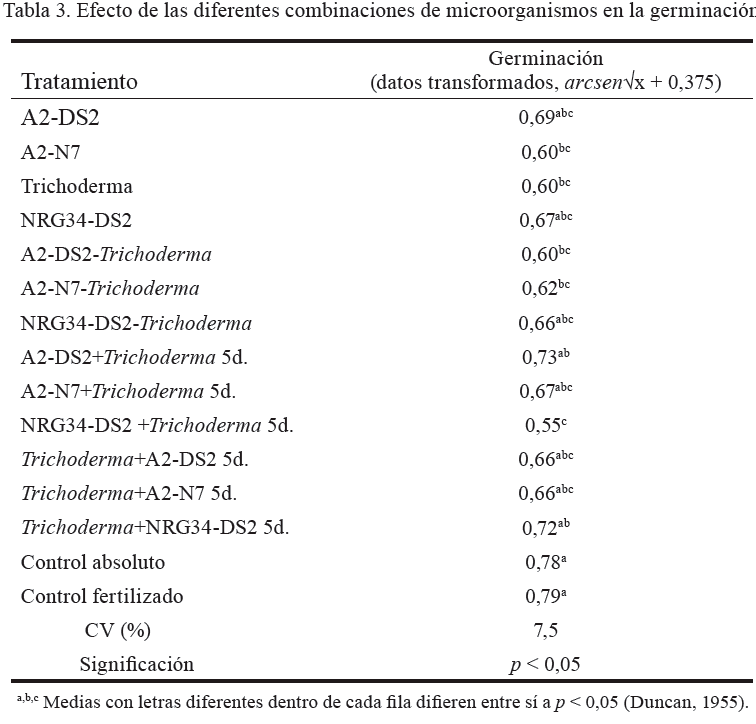
Germinación
Los valores de germinación (tabla 3) mostraron poca o ninguna influencia positiva de las cepas inoculadas. Tanto el control fertilizado como el control absoluto resultaron estadísticamente superiores a A2-N7, Trichoderma, A2-DS2-Trichoderma, A2-N7-Trichoderma, NRG34-DS2 + Trichoderma 5d.; y no difirieron del resto de los tratamientos.
Estos resultados no coinciden con los encontrados por Benítez et al. (1998), quienes aseguran que Trichoderma produce sustancias estimuladoras del crecimiento que influyen en el aumento de la tasa de germinación en las semillas. Asimismo, Arora et al. (2001) reportaron que cepas productoras de sideróforos de S. meliloti incrementaron la germinación en maní, en presencia de Macrophomina phaseolina. Sin embargo, Canto-Martin et al. (2004) no observaron diferencias en la germinación de semillas de ají inoculadas con Azospirillum; al igual que Torres et al. (2003), en semillas de Phaseolus vulgaris, inoculadas con A. brasilense, Azotobacter chroococcum y Rhizobium leguminosarum, de forma simple o combinada. Bécquer et al. (2012b) tampoco encontraron diferencias en la germinación, en trigo, al aplicar inóculos formados por Azospirillum y Sinorhizobium. Al parecer, la capacidad estimuladora o depresora de los microorganismos en la germinación depende no solo del género y la especie de estos, sino también del tipo de planta a que pertenecen las semillas.
Consideraciones generales sobre las variables analizadas
A pesar de que varios tratamientos mostraron elevados valores en, al menos, una de las cinco variables, Trichoderma + NRG34-DS2 5d. y A2-N7 se destacaron con evidente superioridad en el contenido de clorofila y en la longitud del tallo (A2-N7); así como en el peso seco aéreo y en el peso seco radical (Trichoderma + NRG34-DS2 5d.). Esta última variable es fundamental, ya que el desarrollo de la planta depende de las raíces. Según señalan Sarig et al. (1992), los cambios en la morfología y fisiología del sistema radical son uno de los efectos más conocidos de las bacterias en el crecimiento vegetal. El incremento del número de raíces laterales y pelos radicales provoca el aumento de la superficie radical disponible para la captación de agua y nutrientes (Bai et al., 2003). De aquí se infiere la importancia de la aplicación temprana de Trichoderma, seguida por una combinación de bacterias rizosféricas que colonizan el interior de las raíces, gracias a la acción celulolítica del hongo (Infante et al., 2009). Sobre la base de estas consideraciones, el tratamiento Trichoderma + NRG34-DS2 5d. puede aportar los mejores resultados en la práctica agrícola; aunque la combinación de las cepas bacterianas A2 y N7 en los experimentos de campo que se deriven de esta investigación también podría ser exitosa.
Es de señalar que el tratamiento fertilizado presentó diferencias significativas respecto al control absoluto en el contenido de clorofila y el peso seco aéreo, no así en el resto de las variables (peso seco radical, longitud del tallo y germinación). En este sentido, Díaz et al. (2008) obtuvieron resultados similares en la longitud del tallo, en experimentos de campo con sorgo, y también Bécquer et al. (2012a), en la germinación. Estos últimos autores reportaron que dos variedades de trigo presentaron valores estadísticamente inferiores en el peso seco radical, en el caso del tratamiento fertilizado respecto al control absoluto.
En la variable peso seco radical, el tratamiento fertilizado y el control absoluto fueron estadísticamente inferiores a algunos de los tratamientos inoculados indistintamente con Sinorhizobium, Azospirillum y Trichoderma, lo cual también demuestra el efecto positivo de estos microorganismos en el desarrollo de la planta.
CONCLUSIONES
Aunque la combinación de las bacterias rizosféricas con Trichoderma en el momento de la siembra, o posterior a esta, no mostró resultados uniformes en las variables estudiadas, el tratamiento Trichoderma + NRG34-DS2 5d. se destacó en el peso seco radical y el peso seco aéreo. También sobresalió el tratamiento A2-N7, en el contenido de clorofila y la longitud del tallo. La aplicación simple de Trichoderma no mostró un efecto positivo en la mayoría de las variables, y ninguno de los tratamientos influyó positivamente en la germinación.
Se recomienda evaluar los tratamientos Trichoderma + NRG34-DS2 5d. y A2-N7 en experimentos de campo con trigo.
REFERENCIAS BIBLIOGRÁFICAS
1. Antoun, H.; Beauchamp, C. J.; Goussard, N.; Chabot, R. & Lalande, R. Potential of Rhizobium and Bradyrhizobium species as plant growth promoting rhizobacteria on non-legumes: effect on radishes (Rhaphanus sativus L.). Plant Soil. 204:57-61, 1998.
2. Antoun, H. & Prevost, D. Ecology of plant growth promoting rhizobacteria. In: Z. A. Siddiqui, ed. PGPR: Biocontrol and biofertilization. Dordrecht, Holland: Springer. p. 1-38, 2005.
3. Arora, N. K.; Kang, S. C. & Maheswari, D. K. Isolation of siderophore-producing strains of Rhizobium meliloti and their biocontrol potential against Macrophomina phaseolina that causes charcoal rot of groundnut. Curr. Sci. India. 81:673-677, 2001.
4. Askary, Mehray; Mostajeran, Akbar; Amooaghaei, Rehaeh & Mostajeran, Maryam. Influence of the Co-inoculation Azospirillum brasilense and Rhizobium meliloti plus 2,4-D on grain yield and N, P, K content of Triticum aestivum (Cv. Baccros and Mahdavi). American-Eurasian J. Agric. & Environ. Sci. 5:296-307, 2009.
5. Avis, T. J.; Gravel, Valerie; Antoun, Hani & Tweddell, R. J. Multifaceted beneficial effects of rhizosphere microorganisms on plant health and productivity. Soil Biol. Biochem. 40:1733-1740, 2008.
6. Barea, J. M.; Azcón, R. & Azcón-Aguilar, C. Mycorrhizal fungi and plant growth promoting rhizobacteria. In: A. Varma, L. Abbott, D. Werner and R. Hampp, eds. Plant surface microbiology. Heidelberg, Germany: Springer-Verlag, p. 351-371, 2004.
7. Bai, Y.; Zhou, X. & Smith, D. L. Crop ecology, management and quality. Enhanced soybean plant growth resulting from coinoculation of Bacillus strains with Bradyrhizobium japonicum. Crop Sci. 43:1774-1781, 2003.
8. Bécquer, C. J.; Antoun, Hani & Prévost, Danielle. Estudio de la interacción de Trichoderma harzianum con bacterias dinitrofijadoras de vida libre. Pastos y Forrajes. 24 (1):35-39, 2001.
9. Bécquer, C. J.; Lazarovits, G. & Lalin, I.. Interacción in vitro entre Trichoderma harzianum y bacterias rizosféricas estimuladoras del crecimiento vegetal. Rev. cub. Cienc. agríc. 47:97-102, 2013.
10. Bécquer, C. J.; Lazarovits, G.; Nielsen, Laura; Quintana, Maribel, Adesina, Modupe; Quigley, Laura et al. C. Effect of inoculation with rhizospheric bacteria in two varieties of wheat. Phase II: greenhouse. Mexican J. Agric. Sci. 3:985-997, 2012a.
11. Bécquer, C. J.; Prévost, Danielle; Juge, Christine; Gauvin, Carole & Delaney, Sandra. Efecto de la inoculación con bacterias rizosféricas en dos variedades de trigo. Fase I: condiciones controladas. Rev. Mex. Cienc. Agríc. 3:973-984, 2012b.
12. Bécquer, C. J.; Ramos, Yamilka; Nápoles, J. A. & Arioza, María D. Efecto de la interacción Trichoderma-rizobio en Vigna luteola SC-123. Pastos y Forrajes. 27 (2):139-145, 2004.
13. Benítez, Tahía; Rincón, Ana M.; Limón, M. Carmen & Codón, A. C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 7:249-260, 2004.
14. Benítez, Tahía; Delgado-Jarana, J.; Rincón, A. M.; Rey, M. & Limón, M. Carmen. Biofungicides: Trichoderma as a biocontrol agent against phytopathogenic fungi. In: S. G. Pandalai, ed. Recent research developments in microbiology. Trivandrum, India. vol. 2. p. 129-150, 1998.
15. Biswas, J. C.; Ladha, J. K. & Dazzo, F. B. Rhizobia inoculation improves nutrient uptake and growth of lowland rice. Soil Sci. Soc. Am. J. 64:1644-1650, 2000.
16. Canto, J. C.; Medina, S. & Morales, A. D. Efecto de la inoculación con Azospirillum sp. en plantas de Chile Habanero (Capsicum chínense Jacquin). Tropical and Subtropical Systems. 4:21-27, 2004.
17. Chacón, M. R.; Rodríguez, O.; Benítez, T.; Sousa, S.; Rey, M.; Llobell, A. et al. Microscopic and transcriptome analyses of early colonization of tomato roots by Trichoderma harzianum. Int. Microbiol. 10:19-27, 2007.
18. Chapman, L. J. & Putman, D. F. Physiography of southern Ontario. Toronto: University of Toronto Press, 1966.
19. Chelius, M. K. & Triplett, E. W. Prokaryotic nitrogen fixation; model system for the analysis of a biological process. England: Horizont Scientific Press, 2000.
20. Díaz, A.; Garza, Idalia; Pecina, V. & Montes, N. Respuesta del sorgo a micorriza arbuscular y Azospirillum en estrés hídrico. Rev. Fitotec. Mex. 31:35-42, 2008.
21. Duncan, D. B. Multiple range and multiple F tests. Biometrics. 11:1-42, 1955.
22. Gravel, V.; Martínez, C.; Antoun, H. & Tweddell, R. J. Control of greenhouse tomato root rot (Pythium ultimum) in hydroponic systems, using plant-growth promoting microorganisms. Can. J. Plant Pathol. 28:475-483, 2006.
23. Harman, G. E.; Howell, C. R.; Viterbo, A.; Chet, I. & Lorito, M. Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2:43-56, 2004.
24. Howell, C. R. The role of antibiosis in biocontrol. In: G. E. Harman and C. P. Kubicek, eds. Trichoderma and Gliocladium. London: Taylor & Francis. vol. 2. Enzymes, biological control and commercial applications. p. 173-184, 1998.
25. Infante, Danay; Martínez, B.; González, Noyma & Reyes, Yusimy. Mecanismos de acción de Trichoderma frente a hongos fitopatógenos. Rev. Protección Veg. 24 (1), 2009. http://scielo.sld.cu/scielo.php?pid=S1010-27522009000100002&script=sci_arttext .
26. Lerch, G. La experimentación en las ciencias biológicas y agrícolas. La Habana: Editorial Ciencia y Técnica, 1976.
27. Lupwayi, N. Z.; Clayton, G. W.; Hanson, K. G.; Rice, W. A & Biederbeck, V. O. Endophytic rhizobia in barley, wheat and canola roots. Can. J. Plant Sci. 84:37-45, 2004.
28. Reino, J. L.; Guerrero, R. F.; Hernández-Galán, R. & Collado, I. G. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 7:89-123, 2008.
29. Rodríguez-Mendoza, María de las N.; Alcántar, G.; Agulilar, A., Etchevers, D. J. & Santizo, J. A. Estimación de la concentración de nitrógeno y clorofila en tomate mediante un medidor portátil de clorofila. Terra Latinoamericana. 16:135-141, 1998.
30. Rudresh, D. L.; Shivaprakash, M. K. & Prasad, R. D. Effect of combined application of Rhizobium, phosphate solubilizing bacterium and Trichoderma spp. on growth, nutrient uptake and yield of chickpea (Cicer aritenium L.). Applied Soil Ecol. 28:139-146, 2005.
31. Saber, W. I. A.; Abd El-Hai, K. M. & Ghoneem, K. M. Synergistic effect of Trichoderma and Rhizobium on both biocontrol of chocolate spot disease and induction of nodulation, physiological activities and productivity of Vicia faba. Res. J. Microbiol. 4:286-300, 2009.
32. Sarig, S.; Okon, Y. & Blum, A. Effect of Azospirillum brasilense inoculation on growth dynamics and hydraulic conductivity of Sorghum bicolor roots. J. Plant Nutr. 15:805-819, 1992.
33. Shaban, W. I. & El-Bramawy, M. A. Impact of dual inoculation with Rhizobium and Trichoderma on damping off, root rot diseases and plant growth parameters of some legumes field crop under greenhouse conditions. Int. Res. J. Agric. Sci. Soil Sci. 1:98-108, 2011.
34. Shoresh, M.; Harman, G. E. & Mastouri, Fatemeh. Induced systemic resistance and plant responses to fungal biocontrol agents. Annu. Rev. Phytopathol. 48:21-43, 2010.
35. Somasegaran, P. & Hoben, H. J. Handbook for rhizobia. New York: Springer-Verlag, 1994.
36. Torres, R.; Pérez, C. & Suárez, Norma. Influencia de la inoculación de rizobacterias sobre la germinación de semillas de frijol común (Phaseolus vulgaris L.). Centro Agrícola. 30:56-60, 2003.
37. Vincent, J. M. A manual for the practical study of root nodules bacteria. Oxford: Blackwell Publications, 1970.
38. Wolffhechel. H. & Jensen, D. F. Use of Trichoderma harzianum and Gliocladium vitens for the biological control of post-emergence damping-off and root rot of cucumbers caused by Pythium ultimum. J. Phytopathol. 136: 221-230, 1992.
39. Zahir, Z. A.; Arshad, M. & Frankenberger, W. T. Plant growth promoting rhizobacteria: applications and perspectives in agriculture. Adv. Agron. 81:97-168, 2004.
Recibido el 13 de junio de 2014
Aceptado el 4 de noviembre de 2014


{kind=link}
{kind=link}
{kind=link}