










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión On-line ISSN 2078-8452
Pastos y Forrajes vol.40 no.3 Matanzas jul.-set. 2017
ARTÍCULO DE INVESTIGACIÓN
Sistemas silvopastoriles intensivos con Leucaena leucocephala (Lam.) de Wit: alternativa productiva en el trópico ante el cambio climático
Intensive silvopastoral systems with Leucaena leucocephala(Lam.) de Wit: productive alternative in the tropic in view of the climate change
Julián Esteban Rivera-Herrera1, Isabel Molina-Botero1, Julián Chará-Orozco1, Enrique Murgueitio-Restrepo1 y Rolando Barahona-Rosales2
1Centro para la Investigación en Sistemas Sostenibles de Producción Agropecuaria (CIPAV) Carrera 25 #6-62, Cali, Colombia
2 Departamento de Producción Animal, Facultad de Ciencias Agrarias, Universidad Nacional de Colombia, Medellín, Colombia
Correo electrónico: jerivera@fun.cipav.org.co
RESUMEN
En este artículo se presenta un análisis descriptivo y valorativo de los beneficios productivos y ambientales de los SSPi con Leucaena leucocephala (Lam.) de Wit, con énfasis en los aspectos de calidad nutricional, producción de biomasa, consumo voluntario, productividad animal y emisiones de gases de efecto invernadero. Para ello se hizo una revisión bibliográfica acerca de las investigaciones durante los últimos 15 años, referentes a los sistemas de producción animal en condiciones tropicales, con énfasis en las realizadas en Colombia. Se comprobó que la oferta forrajera en SSPi con leucaena tiene un contenido de proteína superior al 22 % y un menor contenido de fibra (20-30 %) respecto a la mayoría de los pastos tropicales. Adicionalmente, la leucaenatiene la capacidad de soportar el ramoneo intenso, y su presencia incrementa la productividad vegetal y animal. Los animales producen al menos dos o tres veces más carne y leche ha-1 año-1 como resultado de un mayor consumo de nutrientes y una mayor oferta forrajera, dado que con esta leguminosa el consumo de materia seca (MS) puede ser 1,3 veces mayor que el observado en sistemas convencionales, y la producción forrajera puede incrementarse de 5 a 18 t de MS ha-1 año-1.Por otra parte, la inclusión de L. leucocephala permite disminuir las emisiones entéricas de CH4 en cantidades cercanas a un 20 % por kilogramo de carne o leche producida y por kilogramo de materia seca consumida. En comparación con los sistemas tradicionales, en los que se incluye L. leucocephala, se incrementa la eficiencia en la producción y disminuyen los efectos ambientales negativos de la ganadería.
Palabras clave: calidad forrajera, metano, sistema de pastoreo.
ABSTRACT
This paper presents a descriptive and evaluative analysis of the productive and environmental of SPSi with Leucaena leucocephala (Lam.) de Wit, emphasizing the nutritional quality, biomass production, voluntary intake, animal productivity and greenhouse gas emission aspects. For such purpose, a bibliographic review was made about the studies in the last 15 years, concerning animal production systems under tropical conditions, stressing the ones conducted in Colombia. It was noticed that the forage offer in SPSi with leucaena has a protein content higher than 22 % and a lower fiber content (20-30 %) with regards to most tropical pastures. Additionally, leucaena has the capacity to withstand intense browsing, and its presence increases plant and animal productivity. The animals produce at least two or three times more meat and milk ha-1 year-1 as the result of a higher nutrient intake and higher forage offer, because with this legume the dry matter (DM) intake can be 1,3 times higher than the one observed in conventional systems, and the forage production can be from 5 to 18 t DM ha-1. On the other hand, the inclusion of L. leucocephala allows to decrease the enteric emissions of CH4 in amounts close to 20 % per kilogram of produced meat or milk and per kilogram of consumed dry matter. Compared with the traditional systems, in the ones in which L. leucocephala is included, the efficiency in production is increased and the negative environmental effects of animal husbandry are decreased.
Keywords: forage quality, methane, grazing system.
INTRODUCCIÓN
Leucaena leucocephala(Lam.) de Wit, comúnmente llamada leucaena, acacia forrajera o carbonero blanco, es una leguminosa perenne, con un amplio potencial para la alimentación de los rumiantes y capaz de crecer con gramíneas acompañantes en condiciones tropicales y subtropicales. Se desarrolla en suelos bien drenados, con pH de neutro a alto, por debajo de los 1 400 metros de altitud y con aceptable fertilidad (Murgueitio et al., 2011). Cuando se maneja como arbusto esta especie resulta tolerante a la sequía, y es reconocida por su alto valor forrajero, debido a su capacidad de rebrote, palatabilidad y características nutricionales superiores, que incluyen un alto contenido de proteína cruda y carbohidratos solubles, además de un bajo contenido de fibra (Cuartas et al., 2015a; Gaviria et al., 2015a).
En los últimos años se ha comprobado que existen abundantes evidencias sobre las ventajas nutricionales de L. leucocephala para la alimentación de rumiantes y su contribución para generar sistemas más productivos y sostenibles en condiciones tropicales (Murgueitio et al., 2015; Gaviria et al., 2015b). Debido a su alta producción de forraje y calidad nutricional, en los sistemas que incluyen esta leguminosa es posible emplear una mayor carga animal por hectárea y obtener ganancias de peso y producciones superiores de leche en rumiantes, respecto a las que se alcanzan en otros sistemas forrajeros tropicales (Mohammed et al., 2015; Gaviria et al., 2015b).
De ahí que la intensificación de la ganadería bovina con el uso de L. leucocephala, principalmente en sistemas silvopastoriles intensivos (SSPi), contribuya a la provisión de forrajes de buena calidad, la rehabilitación de ecosistemas degradados y la mitigación y adaptación al cambio climático (Havlík et al., 2013). Las menores emisiones de gases de efecto invernadero (GEI) asociadas a este tipo de sistemas se deben tanto a las características de sus forrajes como a su mayor eficiencia productiva (Molina et al., 2013; Rivera et al., 2016).
Por tanto, en este artículo se hace un análisis descriptivo y valorativo de los beneficios productivos y ambientales de los SSPi con L. leucocephala, con énfasis en los aspectos de calidad nutricional, producción de biomasa, consumo voluntario, productividad animal y emisiones de GEI.
Principales sistemas de pastoreo objeto de estudio en el trópico
En la extensa y detallada revisión bibliográfica acerca de las investigaciones realizadas durante los últimos 15 años referentes a los sistemas de producción animal en condiciones tropicales, con énfasis en las efectuadas en Colombia, entre los principales sistemas de pastoreo que se identificaron se encuentran aquellos con oferta de L. leucocephala y los de praderas de monocultivo de gramíneas.
Los primeros, que incluyeron la leucaena, fueron arreglos silvopastoriles intensivos (SSPi) caracterizados por una alta densidad del cultivar Cunningham de esta leguminosa (> 20 000 arbustos ha-1), asociado con gramíneas como Cynodon plectostachyus (K. Schum.) Pilg. y Megathyrsus maximus (Jacq.) B. K. Simon& S. W. L. Jacobs, y con otros árboles dispersos en el potrero de diferentes características. Estos SSPi se manejaron con rotación mediante el uso de cercas eléctricas, con períodos de ocupación entre 12 y 24 horas y tiempos de descanso cercanos a los 45 días, con oferta de agua y sal ad libitum. Asimismo, tenían entre 5 y 20 años de establecidos, en condiciones de bosque seco tropical (bs-T) y bosque húmedo tropical (bh-T).
Mientras, entre las praderas de monocultivo de gramíneas, llamadas también sistemas convencionales o tradicionales, sobresalieron las pasturas mejoradas (PM) y las pasturas degradadas (PD). Las PM generalmente estaban compuestas por especies como C. plectostachyus y M. maximus, con algún tipo de fertilización y riego esporádico, manejo rotacional y oferta de agua y sal ad libitum. En tanto, las PD estaban constituidas por gramas nativas solas o mezcladas con C. plectostachyus y Dichanthiumaristatum (Poir.) C.E. Hubb., con muy baja producción de biomasa forrajera y manejadas sin fertilización ni riego. En todos los casos, estos sistemas se encontraban localizados en lotes aledaños a los SSPi y también en condiciones de bs-T y/o bh-T.
Como temas importantes de estos tipos de sistemas productivos, a continuación se analizarán los relacionados con la caracterización nutricional del forraje, el consumo voluntario de los animales, la productividad vegetal y animal, así como las emisiones de metano (CH4) y otros GEI.
Composición química general y consideraciones nutricionales en SSPi con L. leucocephala
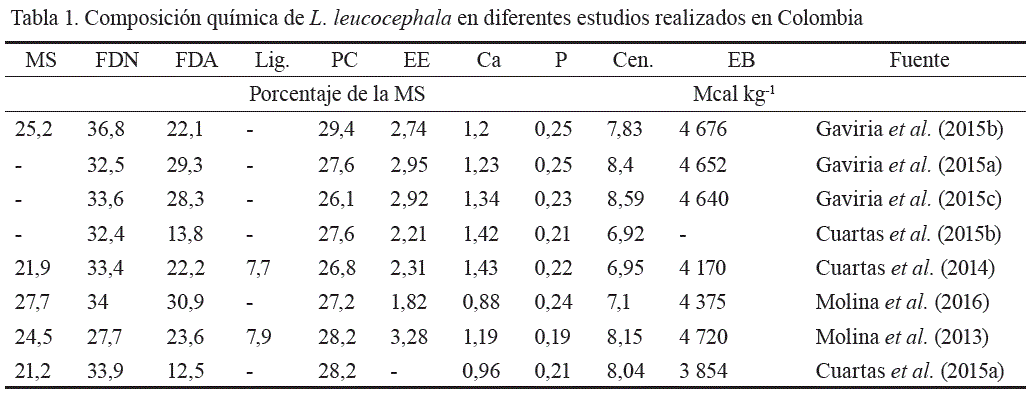
Para la determinación de la composición química y otros aspectos nutricionales de las materias primas y de las dietas en las diferentes investigaciones cuyos resultados se reseñan, se utilizaron métodos convencionales. En general, se evaluaron los siguientes indicadores: contenido de materia seca (MS), fibra en detergente neutro y ácido (FDN y FDA, respectivamente), proteína cruda (PC), cenizas (Cen.), calcio (Ca), fósforo (P), lignina (Lig.), extracto etéreo (EE) y energía bruta (EB), además de la dinámica de degradación y fermentación por medio de la técnica de producción de gases (Mauricio et al., 1999). El contenido de nitrógeno (N) y el de PC fueron determinados por el método de Kjeldahl(ICONTEC, 1999); el de FDN y FDA, según la AOAC (2005); y el EE, por extracción Soxhelet por inmersión (ICONTEC, 1973). El contenido de Cen. se determinó por incineración directa en mufla a 500 °C (AOAC, 2005); el contenido de Ca y P, por espectrofotometría AA y UV-VIS, respectivamente (ICONTEC, 2013); la MS, por la técnica de estufa de aire forzado; y la EB, por calorimetría (ISO, 1998), (tabla 1).
La composición química, la degradabilidad y el comportamiento a nivel ruminal de L. leucocephala mostraron su alto valor nutricional para animales en pastoreo. Singh et al. (2014), Gaviria et al. (2015b) y Cuartas et al. (2015a) informaron que el forraje de L. leucocephala tiene altos niveles de PC (por encima del 22 % de la MS), bajos contenidos de FDN y FDA (menos del 40 y 30 %, respectivamente) y un contenido aceptable de Ca y P. Adicionalmente, L. leucocephala posee niveles intermedios de degradabilidad de la MS (> 50 %) y adecuados porcentajes de fracciones A y B, tanto de carbohidratos (16,7 y 14,94, respectivamente) como de proteínas (34 y 53, respectivamente), por lo que esta leguminosa resulta superior desde el punto de vista nutricional en comparación con los forrajes tradicionalmente utilizados en los sistemas ganaderos tropicales y subtropicales .
La asociación de L. leucocephala con gramíneas en SSPi contribuye a aumentar el contenido de proteína de la dieta (de un 10 % a, por lo menos, un 13 %), en comparación con los mismos pastos en monocultivo (Gaviria et al., 2015a); a la par que se reduce el contenido de FDN, que pasa de 69,8 a 60,7 % en dietas basadas en C.plectostachyus solamente (Cuartas et al., 2013). En consecuencia, la dieta ofrecida en un SSPi se ajusta más a los requerimientos nutricionales de los animales, y así aumenta la eficiencia ganadera.
Consumo de L. leucocephala por bovinos en pastoreo
Para la cuantificación del consumo voluntario, en unas investigaciones se utilizó la técnica de n-alcanos (Mayes et al., 1986), con novillos de 250 a 350 kg de peso vivo en pastoreo, en condiciones de bs-T (Gaviria-Uribe et al., 2015c); mientras que en otros estudios se hizo a través de la cuantificación de la oferta y el rechazo en animales confinados (Molina et al., 2016).
El consumo voluntario, la composición química de los alimentos y su degradabilidad son fundamentales para determinar la cantidad de nutrientes disponibles (Sossa et al., 2015). Un mayor consumo implica que más nutrientes pueden ser aprovechados por los animales, con lo cual pueden incrementar su desempeño productivo.
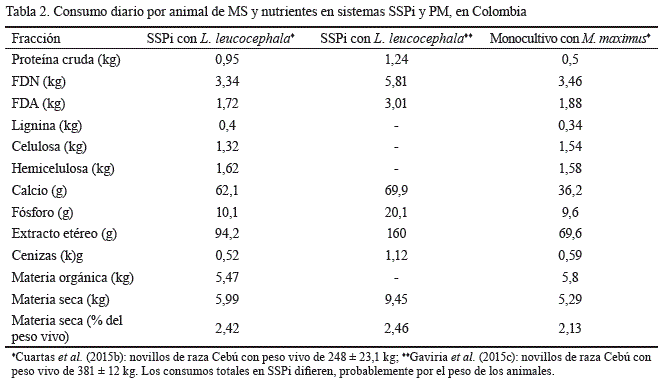
Los animales que pastorearon en SSPi con presencia de L. leucocephalatuvieron consumos de MS hasta 1,3 veces mayores que aquellos que pastorearon en sistemas convencionales de pastoreo (tabla 2). En los SSPi, el consumo de leucaena puede representar entre el 25 y el 30 % del total de MS ingerida (Cuartas et al., 2015b; Gaviria-Uribe et al., 2015c).
Ello puede deberse a los siguientes factores:
1) La mayor oferta de forraje en los SSPi con L. leucocephala, que puede ser hasta el 230 % superior que la de los sistemas convencionales como PM y PD basados solamente en gramíneas tropicales, permite una mayor disponibilidad y selectividad para los animales (Broom et al., 2013; Gaviria-Uribe et al., 2015c).
2) Según Gaviria-Uribe et al. (2015c) y Cuartas et al. (2015a), el contenido de FDN en dietas de M. maximus y C. plectostachyuspuede disminuir en un 15 % cuando se incluye el 25 % de L. leucocephala, mientras que el contenido de FDA puede reducirse en un 20 %. Por otro lado, el bajo contenido de fibra en la dieta mejora el consumo, al permitir tasas de pasaje más altas (Boval y Dixon, 2012). Además, las partículas de leguminosas son cúbicas mientras que las de gramíneas son largas y delgadas, lo que implica mayores tasas de pasaje en especies como L. leucocephala(Barahona-Rosales y Sánchez, 2005).
3) En los SSPi con L. leucocephala, se logra un mayor confort térmico para los animales y estos dedican más tiempo al ramoneo y pastoreo, pues tienen posibilidades de ingerir mayor cantidad de biomasa (Broom et al., 2013). Molina et al. (2016) informaron que al incluir L. leucocephala en un 24 % en la dieta de novillas de levante en confinamiento, el consumo de MS pasó de 2,02 a 2,47 % del peso vivo animal (p = 0,01).
Productividad vegetal y animal
Para la cuantificación de MS disponible en praderas en monocultivo, Cajas-Girón et al. (2011) y Gaviria et al. (2015b) utilizaron la técnica de aforos con doble muestreo, de acuerdo al método descrito por Haydock y Shaw (1975); mientras que para la biomasa proveniente de los arbustos de leucaena emplearon una modificación de ese mismo método, con una escala de cinco estratos en segmentos de un metro lineal, representativos de diferentes niveles de crecimiento. En estos segmentos se cosechó y pesó el material vegetal que era consumido por los animales (hojas y tallos finos), y con la referida escala se realizaron al menos 50 observaciones para estimar la disponibilidad de forraje de cada franja.
Por otra parte, la estimación de la productividad animal, según los estudios de Gaviria et al. (2012), Cuartas et al. (2013), Rivera et al. (2013) y Mohammed et al. (2015), se efectuó mediante pesajes periódicos a los animales y a la leche producida en los sistemas de doble propósito y de lechería especializada; mientras que la producción por hectárea se estimó a partir de las producciones individuales en cada sistema y la carga animal, que a su vez se determinó teniendo en cuenta la oferta (aforos) y la demanda (requerimientos) de MS.
Debido a la mejora de la calidad nutricional y al mayor consumo de nutrientes, en los sistemas que incluyen L. leucocephala se produce más carne y leche que en los convencionales (Cuartas et al., 2014). Además, la ganadería basada en estos sistemas es más rentable (González, 2013).
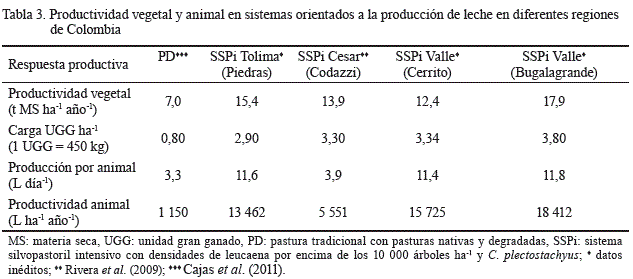
En relación con la productividad vegetal, en los SSPi con leucaena la oferta de biomasa puede ser superior a las 16 t de MS ha-1 año-1, con menores fluctuaciones en el período poco lluvioso y sin la aplicación de fertilizantes (Gaviria et al., 2012; Gaviria et al., 2015b). Esta oferta es similar a la informada por Naranjo et al. (2012) para sistemas de gramíneas con riego y alta dependencia de insumos externos (tabla 3), y superior a las 7 t ha-1 año-1 de MS disponibles que se señalan para PD que cubren una buena parte del país (Cajas-Girón et al., 2011).
Según Gaviria et al. (2015a) y Cuartas et al. (2015b), el aporte de L. leucocephala puede estar alrededor de las 4,3 t ha-1 año-1 de MS. La mayor producción de biomasa en SSPi respecto a los sistemas convencionales se debe a la presencia de estratos aéreos que permiten mayor captura de luz solar y sistemas radiculares más profundos que absorben nutrientes y humedad de manera más eficiente (Murgueitio et al., 2011; Broom et al., 2013), y a la capacidad de esta leguminosa de fijar N atmosférico y aportarlo a las gramíneas acompañantes.
En cuanto a la producción de leche, en la región del Caribe seco colombiano Rivera et al. (2015b) reportaron 5 551 L ha-1 año-1 en un SSPi con L. leucocephala(10 000 arbustos ha-1), C. plectostachyus y M. maximus; y 1 150 L ha-1 año-1en un sistema convencional de pastoreo sin árboles. Asimismo, en el SSPi el contenido de proteína, grasa y sólidos totales en la leche fue significativamente superior (p< 0,05). Las producciones de sólidos totales, grasa y proteína por animal día-1 fueron de 0,59 vs. 0,51; 0,22 vs. 0,17 y 0,15 vs. 0,13 kg para el SSPi y la PD, respectivamente. Por otra parte, Paciullo et al. (2014) informaron que en Brasil la asociación de leucaena con pasturas de Urochloadecumbens(Stapf) R. D. Webster (= BrachiariadecumbensStapf.) permite incrementar la producción de leche de 9,5 a 10,4 litros vaca día-1 (p< 0,05).
Las características de la leche producida en SSPi permiten aumentar el rendimiento de productos lácteos y la eficiencia en la transformación en queso en la época de seca, respecto a la leche producida por animales suplementados con sorgo. A pesar de utilizar menor suplementación, el SSPi no difirió del sistema convencional en cuanto a la producción, según Mohammed et al. (2015), quienes indicaron también una mayor rentabilidad económica en el sistema con leucaena. Por su parte, González (2013) estimó tasas internas de retorno (TIR) del 13 % en sistemas con leucaena y de solo un 0,7 % en sistemas convencionales, en México.
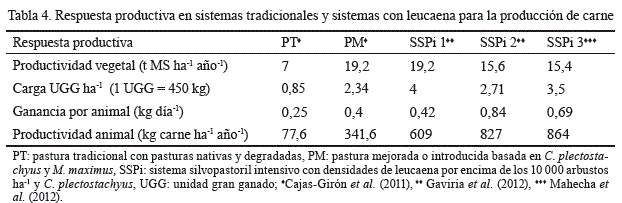
En relación con la productividad cárnica, al analizar en Colombia dos SSPi con leucaena, Gaviria et al. (2012) observaron que las ganancias individuales de peso resultaron mayores en un 45 y 70 %, respecto a las de PM y PD. Además, la productividad por hectárea fue muy superior en los SSPi, gracias a la mayor carga animal (2,34 vs.4 UGG) y ganancia de peso animal (830 kg ha-1 año-1); mientras que en el país el promedio de producción en sistemas de carne está cercano a los 120 kg ha-1 año-1(tabla 4).
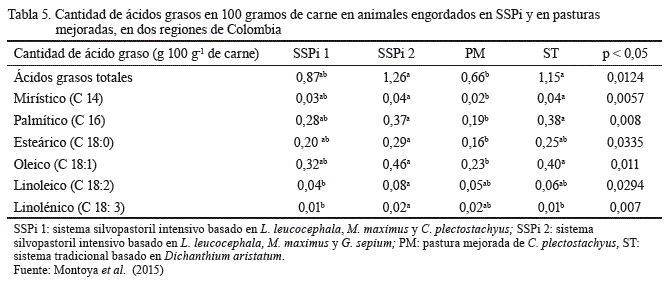
Finalmente, en relación con la calidad de la carne, Montoya et al. (2015) encontraron que animales provenientes de sistemas con leucaena produjeron carne de mayor terneza, mejor pH y color, así como también alcanzaron mayores pesos al beneficio, peso en canal caliente y porcentaje de rendimiento, que los animales de sistemas tradicionales de pastoreo. También se observó que la dieta del SSPi puede mejorar el perfil de ácidos grasos, al incrementar el porcentaje de ácidos grasos insaturados, lo cual genera la posibilidad de entrar a mercados especiales con mayor valor comercial. En la tabla 5 se presentan algunos resultados obtenidos por los autores anteriormente citados en cuanto a la cantidad de ácidos grasos en 100 g de carne.
Emisiones de metano (CH4) y otros GEI
Las mediciones de CH4 emitido por fermentación entérica fueron realizadas por Molina et al. (2016), mediante la técnica del politúnel, en novillas de 218-280 kg de peso vivo, en condiciones de bs-T. A su vez, para determinar los flujos de CH4, N2O y CO2 provenientes de praderas, Rivera et al. (2015) utilizaron la técnica de cámaras cerradas estáticas. Mientras que para estimar la huella de carbono, Rivera et al. (2016) usaron la metodología de la ISO 14044 para el cálculo de emisiones dentro y fuera (fabricación y transporte de insumos) de la finca.
En diferentes estudios, se ha evidenciado la capacidad de los sistemas con L. leucocephala para reducir las emisiones de GEI en bovinos. Estas determinaciones incluyen la producción de CH4 por fermentación entérica, tanto in vivo(Molina et al., 2013; Molina et al., 2016) como in vitro(Huang et al., 2011; Molina et al., 2013; Rivera, 2015), la determinación de la huella de carbono y el balance de GEI (Naranjo et al., 2012; Harrison et al., 2015; Rivera et al., 2016), y la medición de los flujos de gases provenientes de praderas y excreciones bovinas.
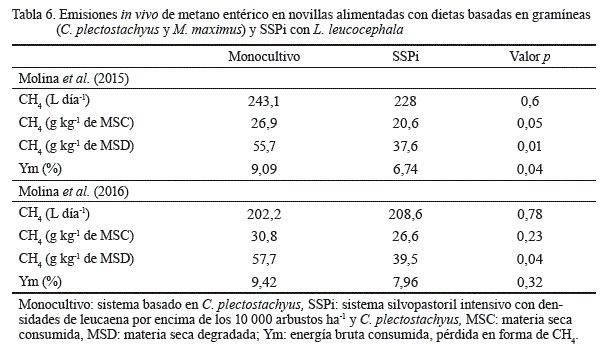
En lo referente a las emisiones entéricas de CH4, Molina et al. (2016) informaron que con dietas basadas en C.plectostachyus e inclusión de aproximadamente 25 % de L. leucocephala, ofrecidas a novillas, estas emisiones pueden disminuir en 15 % por kilogramo de materia seca consumida (MSC), ya que disminuyeron de 43,6 a 37,7 L de CH4 kg-1 de MSC (tabla 6). Además, hubo menor pérdida de energía por producción de CH4 en las dietas con L. leucocephala. Molina et al. (2015) hallaron resultados similares al evaluar las emisiones entéricas de metano en respuesta a una inclusión de 24 % de L. leucocephala en dietas basadas en C.plectostachyus y M. maximus.
En ambas evaluaciones, a pesar de que los animales en los sistemas con leucaena consumieron de un 15 a 20 % más de MS, las emisiones totales por animal solo se incrementaron en un 3 % como promedio, sin diferencias significativas. Ello se debe a que con la inclusión de la leucaena disminuyen los tenores totales de FDN, y esto reduce las emisiones de metano (Archimède et al., 2011). Otro elemento que contribuye a la disminución de las emisiones de CH4 cuando se ofrece leucaena es la presencia de taninos condensados (Barahona-Rosales et al., 2000). Estos componentes posiblemente inhiban el crecimiento de Archaea en el rumen, y su efecto depende de la estructura química y de su cantidad (Archimède et al., 2011; Huang et al., 2011).
En un estudio in vitro realizado por Riveraet al. (2015) en Colombia, se halló que con la inclusión de un 25 % de leucaena en dietas con C.plectostachyus se disminuyó el CH4 en un 8 % por kilogramo de MS potencialmente consumida (p = 0,0029) y en un 13 % por kilogramo de MS degradada (p = 0,0016).
Por otra parte, la inclusión de L. leucocephala resulta en una menor huella de carbono en la carne y la leche producidas en SSPi. Rivera et al. (2016) informaron que en un SSPi con leucaena disminuyeron en un 12 % los kilogramos de CO2-eq emitidos en la producción de un kilogramo de leche corregida por grasa y proteína (LCGP); en relación con un sistema intensivo basado en pasto estrella, con riego, oferta de alimentos concentrados y fertilización, en condiciones tropicales (2,05 vs. 2,34 kg de CO2-eq, respectivamente). A su vez, en el sistema con leucaena decrecieron en un 19 y un 23 % las emisiones de GEI asociadas con la producción de un kilogramo de grasa y de proteína (42,3 vs. 54,9 CO2-eq kg-1 y 47,3 vs 58,3 CO2-eq kg-1, respectivamente).
Rivera et al. (2016) reportaron que en sistemas basados en L. leucocephala se reduce la intensidad de las emisiones gracias a su alta productividad, calidad de la dieta, baja dependencia de insumos externos (como fertilizantes y alimentos concentrados) y alta carga animal. Asimismo, Naranjo et al. (2012)estimaron que, en condiciones de alta densidad arbustiva y presencia de árboles en las zonas de pastoreo, el balance entre emisiones y remociones puede llegar a ser negativo, gracias a que en SSPi se pueden capturar entre 17 y 32 t de CO2ha-1 año-1 y emitir cerca de 12 t de CO2 ha-1 año-1.
En cuanto a las emisiones en praderas con presencia de leucaena, Harrison et al. (2015) informaron que estos flujos de gases pueden ser inferiores a los de sistemas convencionales con similar oferta de N en la dieta. En condiciones de bosque seco tropical, Rivera et al. (2015) hallaron menores pérdidas de N depositado en estiércol y orina por bovinos en forma de N2O en un sistema silvopastoril intensivo, respecto a un sistema tradicional (p = 0,002). Así, en el SSPi solo se emitió el 1,37 % del N excretado vía estiércol, comparado con el 1,77 % emitido en el sistema tradicional; mientras que para el caso de la orina las emisiones fueron de 3,47 vs. 0,3 % para el sistema tradicional y el SSPi, respectivamente. Al observar los flujos de las praderas, estos mismos autor encontró que las emisiones de un sistema con leucaena fueron similares a las de un bosque (p> 0,05) y muy inferiores a las de un sistema de gramíneas en monocultivo con riego y fertilización (p = 0,001). En sus resultados, la alta intensidad de fertilización en el sistema convencional (420 kg de N ha-1 año-1) y el riego, indudablemente, favorecieron las condiciones para incrementar procesos de desnitrificación y, por ende, las emisiones netas de N2O hacia la atmósfera.
Mecanismos que explican las respuestas productivas y las emisiones de CH4 con la presencia de L. leucocephala
Desde el punto de vista nutricional, un aspecto a tener en cuenta en este tipo de especies es su degradabilidad, indicador que describe la cantidad de nutrientes verdaderamente disponibles para los animales. Makkar (2003) y Mota et al. (2005) señalaron que, si bien leucaena no es de alta digestibilidad debido a la presencia de metabolitos secundarios, como los taninos condensados, su combinación con gramíneas incrementa la degradabilidad de la mezcla forrajera si se compara con los valores individuales de cada especie, lo cual aumenta la disponibilidad de nutrientes para ser utilizados a nivel ruminal y por el animal. De igual forma, autores como Barahona-Rosales et al. (2000), Molina et al. (2013) y Gaviria et al. (2015a) hallaron valores de degradabilidad por encima del 60 %, lo que evidencia posibles efectos ambientales que afectan las características nutricionales de L. leucocephala. Es común observar aumentos de un 10 % en la digestibilidad cuando se incluye L. leucocephala junto con gramíneas como M. maximus o C. plectostachyus.
Aunque la degradabilidad de la arbustiva puede parecer poca, su presencia contribuye a una mayor cantidad de N tanto en el rumen como en el duodeno, dado que realiza un aporte considerable de proteína total como de sobrepaso. Esto conlleva un balance de N para la síntesis de proteína microbiana y para satisfacer las necesidades del animal (Mota et al., 2005). La degradabilidad in vitro de la proteína de leucaena varía entre 47,8 (García et al., 2009) y 66,9 % (Gaviria et al., 2015b).
Finalmente, en relación con la dinámica de fermentación a nivel ruminal de la leucaena, diferentes autores informan propiedades que contribuyen al mejor desempeño animal y aprovechamiento de los recursos forrajeros. Sommart et al. (2000) sugirieron que el volumen de gas es un buen parámetro para predecir la degradabilidad y la síntesis de proteína microbiana en el rumen. Además, se ha demostrado que la digestibilidad de la MS y de la materia orgánica (MO) tiene una alta correlación con el volumen de gas producido. Huang et al. (2011), Molina et al. (2013), Rivera et al. (2013) y Rivera et al. (2015) reportaron que cuando L. leucocephalau otras arbustivas son mezcladas con gramíneas hay una mayor producción de gas en horarios tempranos, lo cual favorece una mayor cantidad de sustratos de rápido aprovechamiento por microorganismos presentes en el rumen y, por ende, una mayor eficiencia.
Se ha sugerido que lo que ejerce más influencia en la emisión de metano en el tracto gastrointestinal es la composición de la dieta, en particular el valor proteico y el lipídico y el tipo de carbohidratos (Ulyatty Lassey, 2001); seguido de elementos como la especie forrajera, el estado de madurez, los métodos de conservación, los tratamientos químicos o físicos (Tan et al., 2011) y la presencia de metabolitos secundarios (Archimède et al., 2011), que intervienen directa o indirectamente en la digestibilidad, la tasa de pasaje, el nivel de consumo y el peso vivo del animal (Singh et al., 2014). Como se discutió anteriormente, en la mayoría de estos factores la inclusión de L. leucocephala genera beneficios, lo que a su vez explica por qué con esta leguminosa se emite menos metano durante la fermentación entérica.
Uno de los mecanismos mediante los cuales la inclusión de L. leucocephala puede inhibir la producción de metano es su contenido de taninos condensados (TC), que son polifenoles con la capacidad de unirse y precipitar las proteínas, los carbohidratos y otras moléculas (Haslam, 1986). Considerados componentes vegetales antiherviboría, la mayoría de los estudios en nutrición animal con TC fueron inicialmente realizados para caracterizar el impacto de estos sobre la disponibilidad de proteína y energía en animales que consumían leguminosas taníferas(Barahona-Rosales et al., 2000).
A medida que se avanzó en la investigación, se pudieron identificar taninos con efectos nutricionales beneficiosos (vgr Lotus corniculatus L.) o deletéreos (vgr Lotus uliginosus Schkuhr), los cuales fueron asociados después con diferencias en el peso molecular de los taninos de ambas especies (Foo et al., 1997). En una comparación de siete leguminosas tropicales, Barahona-Rosales et al. (2006) demostraron que los taninos de diferentes leguminosas variaban en sus efectos inhibitorios sobre la actividad de las enzimas celulolíticas y hemicelulolíticas, en respuesta a cambios en su contenido de antocianidinas y su peso molecular.
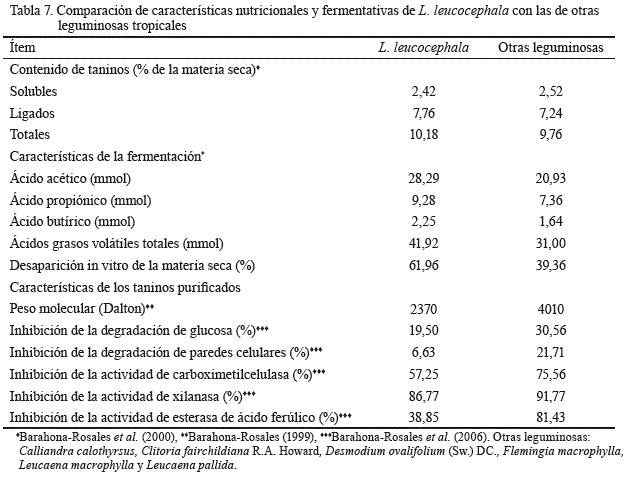
Uno de los avances más significativos de los últimos años es que la presencia de taninos condensados en leguminosas puede contribuir a una reducción en las emisiones de CH4 entre el 13 y el 16 % (Patra y Saxena, 2010). Sin embargo, en la gran mayoría de los casos, dichas disminuciones han ido acompañadas con reducciones de la digestibilidad de la fibra, lo cual contrarresta el beneficio de la reducción en emisiones, al afectar negativamente la productividad en respuesta a la limitada disponibilidad de energía a partir de la fibra del forraje. En los estudios de Barahona-Rosales et al. (2000) y Barahona-Rosales et al. (2006), se demostró que los TC de L. leucocephala son de carácter beneficioso, a diferencia de la gran mayoría de los taninos de otras leguminosas tropicales, que actúan más como agentes antinutricionales (tabla 7).
Al evaluar los TC de L. leucocephala híbrido Rendang, Tan et al. (2011) hallaron que la producción de metano in vitro de M. maximus se redujo linealmente entre el 1 y el 3 %, en presencia de niveles crecientes de TC. Por su parte, Sallam et al. (2010), cuando evaluaron los TC de varias leguminosas entre las que se encontraba leucaena, identificaron que los taninos podrían reducir hasta en 88 % la producción de metano in vitro. El efecto de los TC es atribuido a la acción indirecta sobre la metanogénesis, por la reducción de la producción de H2 resultante de la degradación de MO y por la acción directa sobre las bacterias metanogénicas. De igual forma Tavendale et al. (2005) informaron que los TC pueden reducir hasta el 12 % las poblaciones de bacterias metanogénicas, y Puchala et al. (2012) lograron identificar reducciones en las poblaciones de protozoos; aunque Angarita et al. (2015) no encontraron efecto de la presencia de taninos de leucaena en la población de bacterias metanogénicas.
Como se puede observar en la tabla 7, a pesar de contener casi la misma cantidad de taninos ligados, solubles y totales en su forraje que el promedio de las otras leguminosas, el forraje de L. leucocephala fue 1,57 veces más degradable y tuvo mayor acumulación de ácidos grasos volátiles (p< 0,05) cuando fue fermentado en experimentos de producción de gas (Barahona-Rosales et al., 2000). Al evaluar la capacidad de taninos purificados de las mismas leguminosas, con el fin de inhibir la degradabilidad in vitro de varios sustratos o la actividad de varias enzimas hidrolíticas producidas por el hongo anaeróbico Neocallimastix hurleyensis Vavra & Joyon ex I.B.Heath, en la mayoría de las ocasiones se observaron las menores reducciones en estos parámetros en presencia de taninos de L. leucocephala(Barahona-Rosales et al., 2006).
Es importante destacar que fueron precisamente los taninos de L. leucocephala los que mostraron el menor peso molecular en el experimento de Barahona-Rosales et al. (2006) (tabla 7). El peso molecular de L. leucocephala sugiere una molécula que, en promedio, tiene ocho antocianidinas (i.e. pelargonidina, delfinidina, fisetinidina, entre otras). Debido a su menor peso molecular, los taninos de L. leucocephala no son tan eficientes cuando se unen y precipitan la proteína y otros nutrientes, ya que no están disponibles para los microorganismos en el rumen de los bovinos (Barahona-Rosales et al., 2006). De ahí que, entre las leguminosas evaluadas, L. leucocephala fue la de más alta degradabilidad y la de mayor producción de ácidos grasos volátiles. Esto sugiere que si los taninos de L. leucocephala participan en la reducción de emisión entérica de metano, probablemente lo hacen por mecanismos de toxicidad hacia los microorganismos del dominio Archaea.
Consideraciones finales
Los SSPi que empleen leucaena constituyen una alternativa ventajosa para incrementar la producción de carne y leche entre dos y cinco veces más por unidad de área, ya que en ellos se produce un mayor aporte de MS, PC, energía y algunos minerales, así como un menor contenido de FDN y FDA, y una mayor eficiencia en la fermentación ruminal.
Por otra parte, debido a la oferta de biomasa, en los SSPi con leucaena se puede emplear una mayor carga animal (hasta cuatro veces más que en un sistema convencional degradado y hasta dos veces que en uno con pasturas mejoradas), y se obtienen productos de mejor calidad. Además, como consecuencia de la notable eficiencia en la fermentación ruminal y la menor dependencia de insumos externos, como fertilizantes y alimentos concentrados comerciales, se genera un 15 % menos de emisiones de GEI por unidad de producto y alrededor del 20 % menos por kilogramo de MS consumida o degradada. Tales características podrían favorecer a estos sistemas para ingresar a mercados que demanden productos pecuarios bajos en carbono que contribuyan a mejorar la salud humana.
En cuanto a perspectivas de investigación, se recomienda ampliar el conocimiento en temas como calidad bioactiva de los productos animales para perfilarlos como productos nutracéuticos, determinar el impacto de estos sistemas en emisiones de GEI provenientes del suelo y excreciones animales, y realizar evaluaciones económicas detalladas en diferentes condiciones de producción. Además, se recomienda evaluar la captura de carbono y los efectos beneficiosos sobre la salud del suelo y la fauna asociada.
REFERENCIAS BIBLIOGRÁFICAS
1. Angarita, Erika; Molina, Isabel; Villegas, G.; Mayorga, Olga; Chará, J. & Barahona, R. Quantitative analysis of rumen microbial populations by qPCR in heifers fed on Leucaena leucocephala in the Colombian Tropical Dry Fores. Acta Sci., Anim. Sci. 37 (2):135-142. http://www.scielo.br/pdf/asas/v37n2/1807-8672-asas-37-02-00135.pdf , 2015.
2. AOAC. Animal feed. Determination of ash content by ashing. 942.05. 18 ed. Gaithersburg, USA: AOAC International, 2005.
3. Archimède, H.; Eugène, M.; Magdeleine, Marie; Boval, M.; Martin, C.; Morgavi, D. P. et al. Comparison of methane production between C3 and C4grasses and legumes. Anim. FeedSci. Technol. 59-64:166-167, 2011.
4. Barahona-Rosales, R. Condensed tannins in tropical forage legumes: their characterisation and study of their nutritional impact from the standpoint of structure-activity relationships. PhD Thesis. United Kingdom, University of Reading, 1999.
5. Barahona-Rosales, R.; Lascano, C. E.; Narvaez, N.; Owen, E.; Morris, P. & Theodorou, M. K. In vitrodegradability of mature and immature leaves of tropical forage legumes differing in condensed tannin and non-starch polysaccharide content and composition. J. Sci. Food Agric. 83 (12):1256-1266, 2000.
6. Barahona-Rosales, R. & Sánchez, S. Limitaciones físicas y químicas de la digestibilidad de pastos tropicales y estrategias para aumentarla. Revista CORPOICA. 6 (1):69-82. http://www.redalyc.org/pdf/4499/ 449945018010.pdf, 2005.
7. Barahona-Rosales, R.; Sánchez, Solange; Lascano, C. E.; Owen, E.; Morris, P. & Theodorou, M. K. Effect of condensed tannins from tropical legumes on the activity of fibrolytic enzymes from the rumen fungus Neocallimastyx hurleyensis. Enzyme Microb. Tech. 39 (2):281-288, 2006.
8. Boval, M. & Dixon, R. M. The importance of grass lands for animal production and other functions: a reviewon management and methodological progress in the tropics. Animal. 6 (5):748-762, 2012.
9. Broom, D. M.; Galindo, F. A. & Murgueitio, E. Sustainable, efficient livestock production with high biodiversity and good welfare for animals. P. Roy. Soc. Lond. B Bio. 2025:1471-2954. http://rspb.royalsocietypublishing. org/content/royprsb/280/1771/20132025.full.pdf, 2013.
10. Cajas-Girón, Yazmín S.; Cuesta, P. A.; Arreaza-Tavera, L. C. & Barahona-Rosales, R. Implementación de estrategias tecnológicas para mejorar la productividad y sostenibilidad de sistemas de doble propósito en las sabanas de la región Caribe colombiana. Rev. Colomb. Cienc. Pecu. 24 (3):495, 2011.
11. Cuartas, C. A.; Naranjo, J. F.; Tarazona, A.; Correa, G. & Barahona-Rosales, R. Drymatter and nutrientintake and dietcomposition in Leucaena leucocephala - based intensive silvopastoral systems. Trop. Subtrop Agroecosyst. 18:303-311.http://www.revista.ccba.uady.mx/ojs/index.php/TSA/article/view/2125/991, 2015b.
12. Cuartas, C. A.; Naranjo, J. F.; Tarazona, A. M.; Barahona-Rosales, R.; Rivera, J. E.; Arenas, F. et al. Valor nutritivo y cinética de fermentación in vitro de mezclas forrajeras utilizadas en sistemas silvopastoriles intensivos. Zootecnia Trop. 33 (4):295-306, 2015a.
13. Cuartas, C. A.; Naranjo, J. F.; Tarazona, A. M. & Barahona, R. Uso de la energía en bovinos pastoreando sistemas silvopastoriles intensivos con Leucaena leucocephala y su relación con el desempeño animal. CES Medicina Veterinaria y Zootecnia. 8 (1):70-81. http://revistas.ces.edu.co/index.php/mvz/article/view/2858/ 2065, 2013.
14. Cuartas, C. A.; Naranjo, J. F.; Tarazona, A. M.; Murgueitio, E.; Chará, J. D.; Ku Vera, J. et al.Contribution of intensive silvopastoral systems to the adaptation and mitigation of clima techange. Rev. Cienc. Pecu. 27:76-94. http://rccp.udea.edu.co/index.php/ojs/article/viewFile/920/1048, 2014.
15. Foo, L. Y.; Lu, Y.; McNabb, W. C.; Waghorn, G. & Ulyatt, M. J. Proanthocyanidins from Lotus pedunculatus. Phytochemistry. 45:1689-1696, 1997.
16. García, D. E.; Wencomo, Hilda B.; Medina, María G.; Moratinos, P. & Cova, L. J. Caracterización de la calidad nutritiva de 53 accesiones del género Leucaena en condiciones tropicales. Pastos y Forrajes. 32 (1):65-80.http://scielo.sld.cu/pdf/pyf/v32n1/pyf06109.pdf , 2009.
17. Gaviria-Uribe, Xiomara; Naranjo-Ramírez, J. F.; Bolívar-Vargas, D. & Barahona-Rosales, R. Consumo y digestibilidad en novillos cebuínos en un sistema silvopastoril intensivo. Arch. Zootec. 64 (245):21-27.http://www.uco.es/organiza/servicios/publica/az/php/img/web/17_09_57_03_12_3311_4.pdf, 2015c.
18. Gaviria, Xiomara; Naranjo, J. F. & Barahona-Rosales, R. Cinética de fermentación in vitro de Leucaena leucocephala y Megathyrsus maximus y sus mezclas, con o sin suplementación energética. Pastos y Forrajes. 38 (1):55-63.http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S0864-03942015000100006 , 2015a.
19. Gaviria, Xiomara; Naranjo, J. F. & Barahona-Rosales, R. Nutritional quality and fractionation of carbohydrates and protein in the forage components of an intensive silvopastoral system. Pastos y Forrajes. 38 (2):194-201.http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S0864-03942015000200007 , 2015b.
20. Gaviria, Xiomara; Sossa, C. P.; Chará, J.; Barahona-Rosales, R.; Lopera, J. J. & Córdoba, C. P. Producción de carne bovina en sistemas silvopastoriles intensivos en el trópico bajo colombiano. Memorias VII Congreso Latinoamericano de sistemas agroforestales para la producción animal sostenible. Belém do Pará, Brasil. p. 661-665, 2012. https://www.researchgate.net/publication/262936613_Produccion_de_Carne_Bovina_en_ Sistemas_Silvopastoriles_Intensivos_en_el_Tropico_Bajo_Colombiano. 2012.
21. González, J. M. Costos y beneficios de un sistema silvopastoril intensivo (SSPi), con base en Leucaena leucocephala (Estudio de caso en el municipio de Tepalcatepec, Michoacán, México). RIA. 17 (3):35-50. http://www.redalyc.org/articulo.oa?id=83728497004 , 2013.
22. Harrison, M. T.; McSweeney, C.; Tomkins, N. W. & Eckard, R. J. Improving greenhouse gas emissions intensities of subtropical and tropical beef farming systems using Leucaena leucocephala. Agr. Syst. 136:138-146, 2015.
23. Haslam, E. Hydroxybenzoic acid and the enigma of gallic acid. In: E. E. Conn, ed. The Shikimic acid Pathway, recent advances in Phytochemistry. New York: Plenum Press. vol. 20. p. 163-200, 1986.
24. Havlík, P.; Valin, H.; Mosnier, A.; Obersteiner, M.; Baker, J. S.; Herrero, M. et al.Crop productivity and the global livestock sector: Implications for land use change and greenhouse gas emissions. Am. J. Agric. Econ. 95 (2):442-448, 2013.
25. Haydock, K. P. & Shaw, N. H. The comparative yiel method for estimating dry metter yield of pasture. Aust. J. Exp. Agric. Anim. Husb. 15 (76):663-670, 1975.
26. Huang, X. D.; Liang, J. B.; Tan, H. Y.; Yahya, R. & Ho, Y. W. Effects of Leucaena condensed tannins of differing molecular weightson in vitro CH4 production. Anim. Feed Sci. Technol. 166-167:373-376, 2011.
27. ICONTEC. Alimentos y materias primas. Determinación de los contenidos de grasa y fibra cruda. NTC 668. Bogotá: Instituto Colombiano de Normas Técnicas y Certificación, 1973.
28. ICONTEC. Alimento para animales. Determinación del contenido de nitrógeno y cálculo del contenido de proteína cruda. Método Kjeldahl. NTC 4657. Bogotá: Instituto Colombiano de Normas Técnicas y Certificación, 1999.
29. ICONTEC. Alimento para animales. Determinación de los contenidos de calcio, cobre, hierro, magnesio, manganeso, potasio, sodio y zinc. Método usando espectrometría de absorción atómica. NTC 5151. Bogotá: Instituto Colombiano de Normas Técnicas y Certificación, 2013.
30. ISO. Animal feeding stuffs, animal products, and faeces or urine-Determination of gross calorific value-Bomb calorimetermethod. ISO 9831. Geneva, Switzerland: International Organization for Standardization, 1998.
31. Mahecha, Liliana; Corral, Gabriela; Murgueitio, María M.; Sánchez, C.; Ángulo, J.; Olivera, Martha et al. Calidad de carne SSPi en Colombia y México. Carta Fedegan. 129:60-65. http://es.slideshare.net/Fedegan/carta-fedegan-129 , 2012.
32. Makkar, H. P. Quantification of tannins in tree and shrub foliage. A laboratory manual. Netherlands: Kluwer Academic Publishers, 2003.
33. Mauricio, R. M.; Mould, F. L.; Dhanoa, M. S.; Owen, E.; Channa, K. S. & K., Theodorou. M. A semiautomated in vitro gas production technique for ruminant feeds tuff evaluation. Anim. Feed Sci. Technol. 79 (4):321-330, 1999.
34. Mayes, R. W.; Lamb, C. S. & Colgrove, Patricia M. The use of dosed and herbage n-alkanes as markers fo rthe determination of herbage intake. J. Agric. Sci. 107 (1):161-170, 1986.
35. Mohammed, A. H. M.; Aguilar-Pérez, C. F.; Ayala-Burgos, J.; Bottini-Luzardo, María B.; Solorio-Sánchez, F. J. & Ku-Vera, J. C. Evaluation of milk composition and fresh soft cheese froman intensive silvopastoral system in the tropics. Dairy Sci. Technol. 96 (2):159-172, 2015.
36. Molina, I. C.; Angarita, E. A.; Mayorga, O. L.; Chará, J. & Barahona-Rosales, R. Effect of Leucaena leucocephala on methane production of Lucerna heifers fed a diet based on Cynodon plectostachyus. Livest. Sci. 185:24-29, 2016.
37. Molina, I. C.; Cantet, J. M.; Montoya, S.; Correa, G. & Barahona-Rosales, R. In vitromethane production from two tropical grasses alone or in combination with Leucaena leucocephala or Gliricidia sepium . CES Medicina Veterinaria y Zootecnia. 8 (2):15-31. http://www.scielo.org.co/pdf/cmvz/v8n2/v8n2a02.pdf , 2013.
38. Molina, I. C.; Donney`s, G.; Montoya, S.; Rivera, J. E.; Villegas, G.; Chará, J. et al. La inclusión de Leucaena leucocephala reduce la producción de metano de terneras Lucerna alimentadas con Cynodon plectostachyus y Megathyrsus maximus. LRRD. 27 (5).http://www.lrrd.org/lrrd27/5/moli27096.html, 2015.
39. Montoya, C.; García, J. F. & Barahona-Rosales, R. Contenido de ácidos grasos en carnes de bovinos cebados en diferentes sistemas de producción en el trópico colombiano. Revista de la Facultad de Ciencias Farmacéuticas y Alimentarias. 22 (3):205-214, 2015.
40. Mota, M.; Rodríguez, R.; Solanas, E. & Fondevila, M. Evaluation of four tropical browselegumes as nitrogen sources: comparison of in vitro gas production with oher methods to determine N degradability. Anim. Feed Sci. Technol. 123-124 (1):341-350, 2005.
41. Murgueitio, E.; Calle, Zoraida; Uribe, F.; Calle, Alicia & Solorio, B. Native trees and shrubs for the productive rehabilitation of tropical cattle ranching lands. Forest Ecol. Manag. 261 (10):1654-1663, 2011.
42. Murgueitio, E.; Xóchitl, M.; Calle, Zoraida; Chará, J.; Barahona-Rosales, R.; Molina, C. H. et al. Productividad en sistemas silvopastoriles intensivos en América Latina. En: Florencia Montagnini, E. Somarriba, E. Murgueitio, H. Fassola y B. Eibl, eds. Sistemas agroforestales. Funciones productivas, socioeconómicas y ambientales. Turrialba, Costa Rica, Cali, Colombia: CATIE, CIPAV. p. 59-101, 2015.
43. Naranjo, J. F.; Cuartas, C. A.; Murgueitio, E.; Chará, J. & Barahona-Rosales, R. Balance de gases de efecto invernadero en sistemas silvopastoriles intensivos con Leucaena leucocephala en Colombia. LRRD. 24 (8). http://www.lrrd.org/lrrd24/8/nara24150.htm, 2012.
44. Paciullo, D. S.; Pires, M. F.; Aroeira, L. J.; Morenz, M. J.; Maurício, R. M.; Gomide, C. A. et al.Sward characteristics and performance of dairy cows in organic grass-legume pastures shaded by tropical trees. Animal. 8 (8):1264-1271, 2014.
45. Patra, A. K. & Saxena, J. A new perspective on the use of plant secondary metabolites to inhibit methanogenesis in the rumen. Phytochemistry. 71:1198-1222, 2010.
46. Puchala, R.; Animut, G.; Patra, A. K.; Detweiler, G. D.; Wells, J. E.; Varel, V. H. et al.Methane emissions by goats consuming Sericea lespedeza at different feeding frequencies. Animal. 175 (1-2):76-84, 2012.
47. Rivera, J. Análisis del ciclo de vida (ACV) en un sistema silvopastoril intensivo (SSPi) y un sistema intensivo convencional orientados a la producción de leche bajo condiciones de bs - T. Tesis de Maestría: Facultad de Ciencias Agrarias, Universidad Nacional de Colombia, Sede Medellín, 2015.
48. Rivera, J.; Cuartas, C. A.; Naranjo, J. F. & Barahona-Rosales, R. Comparación del comportamiento productivo y calidad de la leche en vacas bajo un sistema silvopastoril intensivo y un sistema de producción convencional en el valle del Río Cesar. Rev. colomb. Cienc. Pecu. 22 (3):514, 2009.
49. Rivera, J.; Chará, J. & Barahona-Rosales, R. Análisis del ciclo de vida para la producción de leche bovina en un sistema silvopastoril intensivo y un sistema intensivo convencional bajo condiciones de bosque seco tropical. Trop. Subtrop. Agroecosyst. 19 (3):237-251, 2016.
50. Rivera, J.; Molina, I. C.; Donney`s, G.; Villegas, G.; Chará, J. & Barahona-Rosales, R. Dinámica de fermentación y producción de metano en dietas de sistemas silvopastoriles intensivos con Leucaenaleucocephalay sistemas convencionales orientados a la producción de leche. LRRD. 27 (4). http://www.lrrd.org/lrrd27/4/rive27076.html, 2015.
51. Rivera, J.; Naranjo, J. F.; Cuartas, C. A. & Arenas, F. A. Fermentación in vitro y composición química de algunos forrajes y dietas ofrecidas bajo un sistema silvopastoril en el trópico de altura. LRRD. 25 (10). http://www.lrrd.org/lrrd25/10/rive25174.htm, 2013.
52. Sallam, S. M.; Bueno, I. C.; Godoy, P. B.; Nozella, E. F.; Vitti, D. M. & Abdalla, A. L. Ruminal fermentation and tannins bioactivity of some browses using a semi-automated gas productiontechnique. Trop. Subtrop. Agroecosyst. 12 (1):1-10. http://www.redalyc.org/pdf/939/93913074001.pdf , 2010.
53. Singh, S.; Anele, U. Y.; Edmunds, B. & Südekum, K. H. In vitro ruminal dry matter degradability, microbial efficiency, short chain fatty acids, carbohydrate and protein fractionation of tropical grass-multipurpose tre especies diets. Livest. Sci. 160:45-51, 2014.
54. Sommart, K.; Parker, D. S.; Rowlinson, P. & Wanapat, M. Fermentation characteristics and microbial protein synthesis in an in vitro system using cassava, rice straw and dried ruzi grass as substrate. Asian-Australas. J. Anim. Sci. 13 (8):1084-1093, 2000.
55. Sossa, Claudia P.; Correa, G. A. & Barahona-Rosales, R. Consumo y excreción de nutrientes en novillos de carne pastoreando en trópico de altura con y sin suplementación energética. Zootecnia Trop. 33 (2):117-128, 2015.
56. Tan, H. Y.; Sieo, C. C.; Abdullah, N.; Liang, J. B.; Huang, X. D. & Ho, Y. W. Effects of condensed tannins from Leucaena on methane production, rumen fermentation and populations of methanogens and protozoa in vitro. Anim. Feed Sci. Technol. 169 (3-4):185-193, 2011.
57. Tavendale, M. H.; Lane, G. A.; Schreurs, Nicola M.; Fraser, K. &Meagher, Lucy P. Theeffects of condensedtanninsfromDorycniumrectumonskatole and indoleruminalbiogenesisforgrazingsheep. Aust. J. Agric. Res. 56 (12):1331-1337, 2005.
58. Ulyatt, M. J. & Lassey, K. R. Methane emissions from pastorla systems: the situation in New Zealand. Arch. Latinoam. Prod. Anim. 9 (1):118-126, 2001.
Recibido el 21 de marzo del 2017
Aceptado el 1 de julio del 2017


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}