










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión On-line ISSN 2078-8452
Pastos y Forrajes vol.40 no.4 Matanzas oct.-dic. 2017
ARTÍCULO DE INVESTIGACIÓN
Macrofauna edáfica como indicador de conversión agroecológica de un sistema productivo de Moringa oleifera Lam. en Nicaragua
Soil macrofauna as indicator of agroecological conversion of a productive system of Moringa oleifera Lam. in Nicaragua
Álvaro Noguera-Talavera, Nadir Reyes-Sánchez, Bryan Mendieta-Araica y Martha Miriam Salgado-Duarte
Universidad Nacional Agraria, km 12 ½ Carretera Norte, Apdo. 453, Managua, Nicaragua
Correo electrónico: nogueratalavera@yahoo.es
RESUMEN
Con el objetivo de determinar la diversidad y funcionalidad de la macrofauna edáfica como indicador biológico de la salud del suelo y el efecto de prácticas de manejo en sistemas productivos de Moringa oleifera Lam., se llevó a cabo un estudio en predios de la Universidad Nacional Agraria, Nicaragua. Los ensayos tuvieron una duración de nueve meses, en los que se implementaron prácticas de manejo agroecológico y manejo convencional. La macrofauna edáfica fue muestreada mediante la metodología desarrollada por el Programa Internacional Biología y Fertilidad del Suelo Tropical. Se realizó identificación taxonómica a nivel de phylum, clase, orden y familia; así como de grupos funcionales: detritívoros, ingenieros del suelo, herbívoros y depredadores. La prueba no paramétrica Kruskal-Wallis se aplicó para determinar diferencias estadísticas en la variación de la densidad por taxón y grupo funcional por sistema de manejo. La densidad de individuos resultó estadísticamente diferente (p < 0,05) entre sistemas de manejo a nivel de clase, orden y familia. La diversidad fue mayor en conversión agroecológica, y se destacó una mayor equitatividad de familias. Los grupos funcionales fueron diferentes entre sistemas, con dominancia de los ingenieros del suelo (64,22 %) en conversión agroecológica, y de los detritívoros (74,19 %) en manejo convencional. Hubo una asociación entre las prácticas de manejo con la densidad y la diversidad de la macrofauna del suelo; y se identificaron los organismos Formicidae y Termitidae y del Coleoptera, que interactuaron con M. oleifera en diferentes estados de desarrollo. El número de unidades taxonómicas constituyó un indicador que permitió distinguir entre los sistemas de manejo, la salud del suelo y la dinámica de transformación de las propiedades químicas, físicas y biológicas de cada sistema.
Palabras clave: biodiversidad, manejo del suelo, organismos indicadores.
ABSTRACT
In order to determine the diversity and functionality of soil macrofauna as biological indicator of soil health and the effect of management practices in productive systems of Moringa oleifera Lam., a study was conducted in areas of the National Agricultural University, Nicaragua. The essays lasted nine months, in which agroecological management and conventional management practices were implemented. The soil macrofauna was sampled through the methodology developed by the International Tropical Soil Fertility and Biology Program. Taxonomic identification at phylum, class, order and family level was performed, as well as of functional groups: detritivores, soil engineers, herbivores and predators. The non-parametric Kruskal-Wallis test was applied to determine statistical differences in the variation of density per taxon and functional group per management system. The density of individuals was statistically different (p < 0,05) between management systems at class, order and family level. Diversity was higher in agroecological conversion, and a higher equitability of families stood out. The functional groups were different between systems, with dominance of soil engineers (64,22 %) in agroecological conversion, and of detritivores (74,19 %) in conventional management. Association was found of the management practices with the density and diversity of the soil macrofauna; and the organisms Formicidae, Termitidae and Coleoptera, which interacted with M. oleifera in different development stages, were identified. The number of taxonomic units constituted an indicator that allowed to distinguish between the management systems, soil health and transformation dynamics of the chemical, physical and biological properties of each system.
Keywords: biodiversity, soil management, indicator organisms.
INTRODUCCIÓN
La agricultura convencional es contextualizada en diversos estudios (Nieto-Gómez et al., 2013; Altieri et al., 2015; Hatt et al., 2016) como el factor causante de la actual crisis ambiental, que conlleva la baja sostenibilidad de la agricultura (Gliessman, 2013), la degradación del suelo, la pérdida de la biodiversidad por simplificación de los ecosistemas (Hatt et al., 2016; Altieri et al., 2017) y la alteración del ciclo hidrológico; en este sentido la agroecología, como ciencia con enfoque multidisciplinario (Nieto-Gómez et al., 2013), representa el resurgimiento de sistemas productivos social, ambiental y culturalmente sostenibles, a través de la promoción de procesos ecológicos beneficiosos para el suelo, la conservación del agua y el manejo de la biodiversidad (Altieri et al., 2017).
En la transición hacia la agricultura ecológica, un principio predominante, de acuerdo con lo expresado por Blanco-Lobaina et al. (2013) y Nicholls et al. (2016), es la mejora en la transformación de las propiedades físicas y químicas y en la funcionalidad biológica del suelo (Matienzo-Brito et al., 2015), debido a que la habilidad de un cultivo de resistir o tolerar el ataque de insectos plagas y enfermedades está ligada a las propiedades biológicas de este recurso (Nicholls y Altieri, 2008).
Según Navarrete et al. (2011), la habilidad que tiene el suelo de sostener la productividad biológica se debe evaluar sobre la base de su funcionalidad específica, ya que integra los componentes biológicos, químicos y físicos en determinadas situaciones de manejo, lo que sugiere una relación entre biodiversidad y productividad. Estos autores refierenque la evaluaci ón de la calidad del suelo permite entender el grado en que las prácticas de manejo aportan a la sostenibilidad. Desde este enfoque, la macrofauna edáfica es un indicador para la valoración de la eficiencia de la agricultura sostenible.
La macrofauna edáfica agrupa los invertebrados mayores de 2 mm de diámetro (Cabrera-Dávila, 2012) y presenta las siguientes características: hábito sedentario, variabilidad a corto plazo en su diversidad y tamaño poblacional (Cabrera-Dávila, 2014), un corto período entre generaciones, alta densidad y capacidad de reproducción que posibilitan un muestreo intensivo. Tales características le permiten que sea utilizada como indicador de monitoreo y diagnóstico de la intensidad de uso del suelo (Díaz-Porres et al., 2014), su estado de conservación o perturbación y salud (Cabrera-Dávila, 2012; Cunha-Neto et al., 2012); así como del efecto, en el tiempo, de prácticas productivas (Socarrás e Izquierdo, 2014; Gómez-Pamies et al., 2016), lo cual está relacionado con lo que señala Cabrera-Dávila (2014) acerca de una mayor variedad y cantidad de organismos en los suelos con un manejo adecuado.
El objetivo del estudio fue determinar la diversidad y funcionalidad de la macrofauna edáfica como indicador biológico de salud del suelo, en sistemas productivos de Moringa oleifera Lam.
MATERIALES Y MÉTODOS
Ubicación geográfica y características edafoclimáticas del área de estudio
El estudio se realizó entre junio de 2013 y marzo de 2014, en la unidad experimental Santa Rosa de la Universidad Nacional Agraria, localizada geográficamente en Managua, en las coordenadas 12o 09' 30.65" N y 86o 10' 06.32" W, a una altitud de 50 msnm (INETER, 2015).
La precipitación y la temperatura media anual históricas registradas son de 1 099 mm y 27o C, respectivamente, y la humedad relativa es de 74 % (INETER, 2015); con predominio de dos estaciones definidas por una época seca que va desde noviembre a abril y una época lluviosa de mayo a octubre.
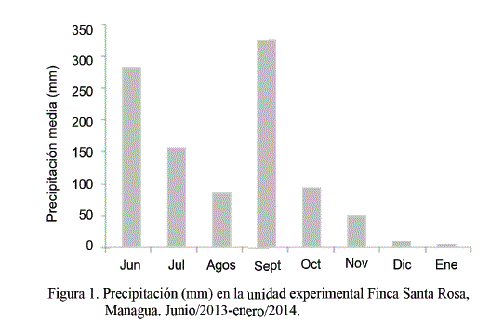
En el año 2013 la precipitación anual en el área fue de 1 070,4 mm (fig. 1), menor en comparación con la media histórica; junio (285,3 mm) y septiembre (330,2 mm) fueron los meses de mayor precipitación (INETER, 2015).
El suelo pertenece al orden taxonómico Andosol; por su clase textural es franco arcilloso arenoso, y presenta buen drenaje.
Descripción de los experimentos
Los ensayos tuvieron una duración de nueve meses, con fecha de establecimiento en junio 2013 y de cosecha en marzo 2014, tiempo durante el cual se aplicaron prácticas de manejo agroecológico y de agricultura convencional.
El área con manejo agroecológico correspondió a un lote de una hectárea, y el agroecosistema con manejo convencional consistió en un área de plantación, con una extensión de 5 ha; en ambos casos se utilizó un área efectiva de muestreo de 0,18 ha. En cada sistema fueron delimitadas cuatro unidades de muestreo de forma rectangular (15 x 30 m). El método definido para muestrear la macrofauna edáfica fue sistemático con monolitos separados a 15 m entre sí, distribuidos en transepto diagonal dentro de las unidades de muestreo.
Vegetación colindante a los sistemas de manejo
- Conversión agroecológica. La flora complementaria al agrosistema estuvo compuesta por cercas vivas con especies arbóreas como Eucalyptus camaldulensis (Dehnh.), Azadirachta indica (A. Juss.), Cordia dentata (Poir.), Pithecellobium dulce (Roxb.) Benth., Albizia saman (Jacq.) Merr. y Stemmadenia obovata (Hook. & Arn.) K. Shum.; mientras que los lotes circundantes correspondieron a uso de suelo agrícola con Moringa oleifera (Lam.) y Sorghum bicolor (L.).
- Manejo convencional. La flora complementaria la constituyeron cercas vivas con especies arbóreas como A. indica (Neem), Spondias sp. y A. saman; mientras que los lotes circundantes correspondieron a producción de pasturas, como CT-15 y Cynodon nlemfuensis (Vanderyst.), además de cultivos como Saccharum officinarum (L.), M. oleifera y S. bicolor (L.).
Diseño experimental y tratamientos. El diseño consistió en unidades experimentales de forma rectangular, seleccionadas de manera aleatoria. Cada unidad contaba con plantas establecidas a una distancia de 3 x 3 m, en arreglo lineal.
- Tratamiento 1. Manejo convencional, el que consistió en un área de monocultivo de moringa, con actividades de preparación mecanizada del suelo, control mecanizado y químico de arvenses, fertilización inorgánica y riego.
- Tratamiento 2. Conversión agroecológica, cuyo enfoque fue el establecimiento de un sistema de policultivo, labranza mínima en la preparación del suelo, rotación de leguminosas, fertilización orgánica con compost, incorporación de abonos verdes, sin riego, y control de arvenses con cobertura de leguminosas.
Evaluaciones. Se realizaron colectas de la macrofauna edáfica antes de la cosecha o explotación de los sistemas. El periodo de colecta fue entre diciembre de 2013 y enero de 2014, al finalizar la época lluviosa, definido a partir de los criterios propuestos por Cabrera-Dávila et al. (2011).
Para la colecta se utilizó la metodología propuesta por el Programa Internacional Biología y Fertilidad del Suelo Tropical (Lavelle et al., 2003). De cada sistema de manejo se extrajeron 12 monolitos de suelo de 25 x 25 cm, hasta una profundidad de 30 cm.
Los especímenes colectados fueron depositados en viales con alcohol al 70 %, y posteriormente se identificaron a nivel de phylum, clase, orden y familia, mediante el uso de claves desarrolladas por Roldan-Pérez (1988) y Castner (2001), en el laboratorio de biología de la Facultad de Recursos Naturales y del Ambiente.
La macrofauna fue clasificada en detritívoros, herbívoros, ingenieros del suelo y depredadores, según los grupos funcionales propuestos por Cabrera-Dávila et al. (2011).
Los resultados se correspondieron con los indicadores: densidad (individuos por m2) de cada taxón y grupo funcional identificado, y diversidad por sistema de manejo; la diversidad y la composición a nivel de familia fueron analizadas como indicadores relacionados con la salud del suelo, según Rendón-Pareja et al. (2011), y su interacción con el cultivo de moringa.
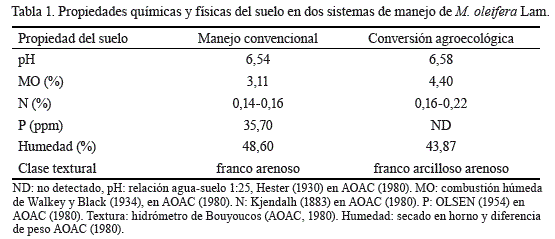
Para la evaluación de los sistemas de manejo, del conjunto de monolitos se colectaron dos muestras compuestas de 2 kg de suelo y se determinaron las propiedades químicas y físicas en el laboratorio de suelos y agua (LABSA) de la Universidad Nacional Agraria (tabla 1).
Procedimiento experimental. La composición del área con manejo agroecológico estuvo constituida por una plantación de moringa con densidad de 1 111 plantas, manejada en policultivo con rotaciones de Canavalia ensiformis (L.), Canavalia brasiliensis (Mart. ex Benth.), Cajanus cajan (L.) Millsp., y Vigna unguiculata (L.) Walp.
El establecimiento y manejo consistió en labranza mínima, control manual de arvenses, fertilización orgánica con compost (N: 0,35 %; P: 0,22 ppm; K: 0,86 meq/100 g de suelo; Cu: 96 ppm, y humedad: 32,07 %); se aplicó 0,03 kg a cada planta al momento de la siembra y después de seis meses de realizada esta, para un total por aplicación de 33,3 kg/ha, incorporación de leguminosas y residuos de cosechas y arvenses, y manejo de arvenses e insectos a través de la cobertura de las leguminosas.
El sistema convencional correspondió a una plantación de moringa en monocultivo con densidad de 1 111 plantas/ha. El establecimiento y manejo se definió por un conjunto de prácticas, como labranza mecanizada en la preparación del suelo (control de arvenses con desbrozadora); volteo del suelo, que consistió en tres pases de gradas; y roturación del suelo entre 20 y 30 cm de profundidad para la elaboración de surcos, con subsolador mecánico.
Se aplicó fertilización inorgánica (N:15-P:30-K:10 a razón de 50 kg/ha) al momento de la siembra y después de un año de establecido el agrosistema; además se empleó riego por aspersión, prácticas de poda, control de plagas a través de químicos inorgánicos (Cipermetrina 100) y control de arvenses con desbrozadora, con una frecuencia de dos a tres veces durante el ensayo, además de control químico (herbicida 2-4-D y glifosato).
Procesamiento estadístico. La estadística no paramétrica (Kruskal-Wallis) se empleó para determinar diferencias en la variación de la densidad por taxón y grupo funcional por sistema de manejo. Como parte de la diversidad de la macrofauna, a nivel de sistema de manejo para los taxones, clases y órdenes, se determinó el índice de dominancia de la comunidad (D) mediante el método propuesto por Turner y Gardner (1991), con comparaciones de t de Student en el programa PAST versión 1.29.
Se realizó un análisis de diversidad a través del método de conglomerados, para determinar la probabilidad de similitud de familias por sistema de manejo y el índice de Jaccard. Este índice expresa el grado en que dos o más muestras son semejantes por las especies presentes en ellas, y se utilizó para el nivel de familia.
RESULTADOS Y DISCUSIÓN
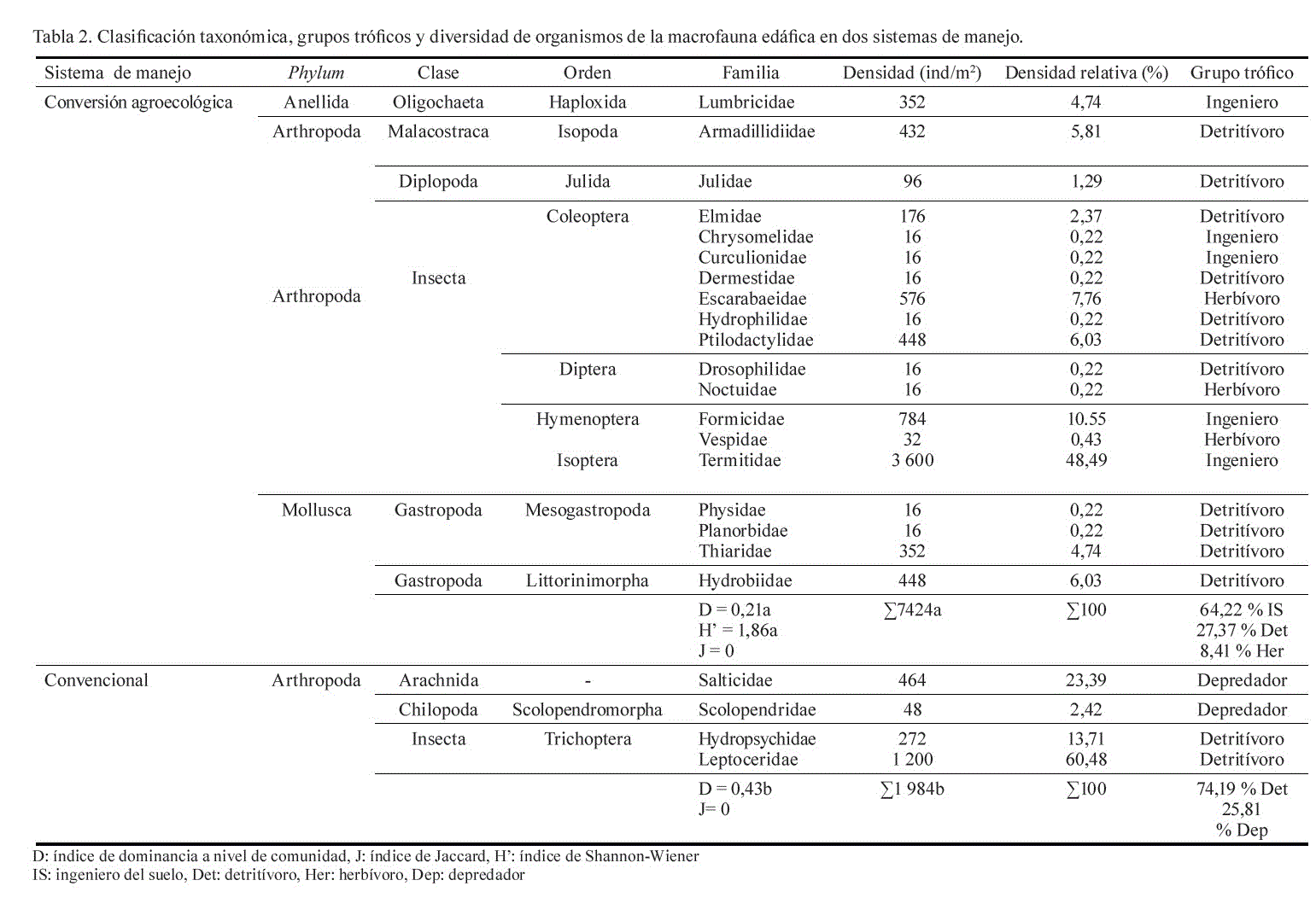
El sistema conversión agroecológica registró la mayor diversidad taxonómica de la macrofauna edáfica, con tres phylum, cinco clases, nueve órdenes y 19 familias; en comparación con un phylum, tres clases, dos órdenes y cuatro familias identificadas en el sistema convencional (tabla 2).
La densidad de individuos fue estadísticamente diferente (p < 0,05) entre sistemas de manejo; se registraron 7 424 ind/m2 en conversión agroecológica, en comparación con 1 984 ind/m2 en manejo convencional (tabla 2). Este resultado coincide con los de Díaz-Porres et al. (2014) y Matienzo-Brito et al. (2015), quienes reportaron diferencias en la densidad y diversidad de la macrofauna edáfica entre sistemas diversos y sistemas ganaderos simplificados, respecto a sistemas intensivos de cultivo, como consecuencia de la complejidad y manejo de los sistemas. Estos autores hallaron que a mayor diversificación mayor actividad biológica, y ello se aplica en este estudio al sistema conversión agroecológica.
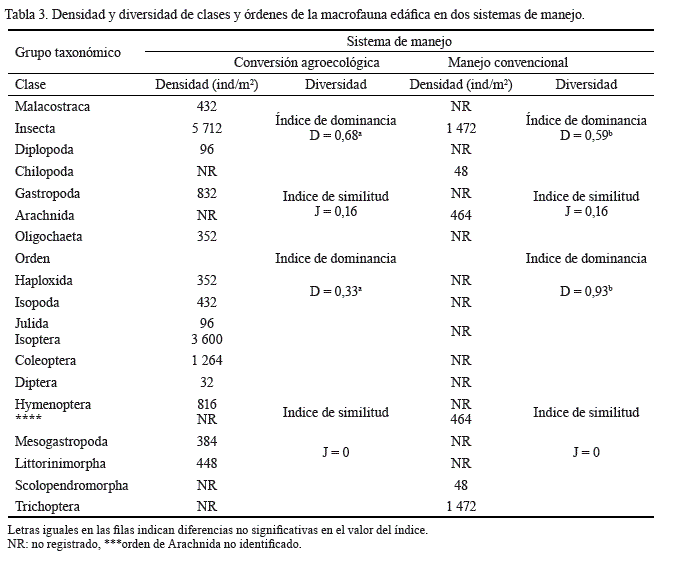
A nivel de clases, los componentes de la diversidad mostraron mayor densidad total en conversión agroecológica, asociada a la dominancia de especímenes del grupo taxonómico Insecta, que se expresó en diferencias significativas (p < 0,05) en el índice de dominancia de la comunidad (D = 0,68); el dominio de la clase Insecta se observó también en manejo convencional (D = 0,59, tabla 3).
Rendón-Pareja et al. (2011), al analizar la dominancia del phylum Artropoda, y dentro de este de la clase Insecta, argumentaron que sus hábitos reproductivos, alimenticios y su distribución e intervención ecológica a nivel del suelo lo hacen útil como indicador biológico del estado de dicho recurso.
La riqueza de clases fue mayor en conversión agroecológica (cinco clases), en comparación con manejo convencional y, por tanto, se determinó baja probabilidad de similitud (J = 0,16) entre los sistemas de manejo.
Sheibani y Gholamalizadeh (2013) informaron que el volteo del suelo durante la labranza tiene efectos sobre los indicadores fisicoquímicos que promueven la diversidad funcional de la macrofauna; mientras que Ayuke et al. (2009) y Díaz-Porres et al. (2014) asociaron la baja diversidad de la macrofauna edáfica a prácticas de manejo tales como el uso de agroquímicos y la modificación (simplificación) del hábitat al establecerse M. oleifera en monocultivo, elementos que permiten explicar los bajos valores de equitatividad registrados.
La menor diversidad en conversión agroecológica, en comparación con los resultados de Ayuke et al. (2009), se atribuyó a la edad del sistema. Al respecto, Nicholls et al. (2016) señalaron que la productividad basada en la diversidad funcional tiende a ser baja durante los primeros tres a cinco años en esquemas de diversificación, en comparación con el manejo convencional, para después incrementarse por efecto de diseños eficientes en cuanto a relaciones de facilitación entre cultivos, lo que aporta no solo al incremento de la diversidad, sino también a su funcionalidad en pro del sistema.
Lo anterior es un indicador del progresivo incremento en los valores de diversidad como parte de la evolución del sistema, según Nicholls et al. (2016), quienes registraron disminución de la diversidad después de algunos años de iniciadas las prácticas basadas en insumos orgánicos, rotación de cultivos e incorporación de leguminosas.
En este sentido, se plantea que la biodiversidad en la agricultura difiere entre agroecosistemas, que a la vez se diferencian en indicadores relativos al tiempo de establecimiento, la constitución de las especies y las prácticas sostenibles.
La clase insecta registró los órdenes con mayor representatividad, en la que se destacaron Isoptera (3 600 ind/m2), Coleoptera (1 264 ind/m2) e Hymenoptera (816 ind/m2) por su alta densidad en el manejo conversión agroecológica. Estos órdenes, según Ayuke et al. (2009), ocupan una importante proporción a nivel de la macrofauna edáfica, lo que confiere una buena dinámica de transformación en las propiedades del suelo en este sistema de manejo; en contraste, en manejo convencional Trichoptera (1 472 ind/m2) presentó la mayor densidad.
Debido a la mayor densidad de Trichoptera, el índice de dominancia de la comunidad registró un valor más alto (D = 0,93) en manejo convencional, en comparación con conversión agroecológica (D = 0,33), con diferencias significativas (p < 0,05) entre los sistemas; no hubo ninguna probabilidad de similitud entre prácticas de manejo, ya que no presentaron órdenes en común (tabla 3). El bajo valor del índice de dominancia en conversión agroecológica demostró que la estructura con diferentes estratos, manejo de bajo impacto y, por tanto, diversidad y calidad de hábitat promueven la equitatividad en la distribución de la macrofauna edáfica y un alto potencial de interacciones dentro del sistema.
La diversidad de familias resultó diferente entre sistemas. En tal sentido, la dominancia de la comunidad (D = 0,43) fue aproximadamente el doble en manejo convencional, en comparación con conversión agroecológica (D = 0,21); por lo que, en este último sistema de manejo, la equitatividad en la densidad de individuos por familia fue mayor (H' = 1,86 vs. 1,0; p < 0,05).
De las 19 familias identificadas en conversión agroecológica, ocho representaron el 94,15 % de la población de la macrofauna edáfica (tabla 2); entre las que sobresalieron Termitidae (48,49 %), Formicidae (10,55 %) y Escarabaeidae (7,76 %). La dominancia de estas familias fue reportada por Ayuke et al. (2009) y Cabrera-Dávila et al. (2011), con representantes que realizan funciones importantes dentro de los agrosistemas, tanto desde el punto de vista de participación en la regulación de la dinámica físico-biológica del suelo, como en su interacción con los cultivos, principalmente como organismos plagas.
En manejo convencional, de las cuatro familias identificadas, Leptoceridae representó el 60,48 % de la población de macrofauna, seguida por Salticidae (23,39 %) e Hydropsychidae (13,71 %); mientras Scolopendridae fue la familia con menor proporción de individuos.
Diferentes autores, entre ellos Díaz-Porres et al. (2014), demostraron que la incorporación de residuos de cosechas, sobre todo cuando estos tienen alto contenido de nitrógeno (leguminosas), promueve un incremento en la densidad de individuos. Por el contrario, cuando en el sistema hay alta cobertura de pastos, que frecuentemente tienen alta relación C/N (Díaz-Porres et al., 2014), la densidad es menor. Los resultados del presente estudio coinciden con lo antes expresado, ya que en conversion agroecológica se realizaron incorporaciones periódicas al suelo de leguminosas y residuos de la poda de moringa, cuyas hojas tienen alto contenido de nitrogeno (2 g en 100 g de materia fresca); mientras que en manejo convencional, la cobertura del suelo fue principalmente pasto estrella (C. nlemfuensis).
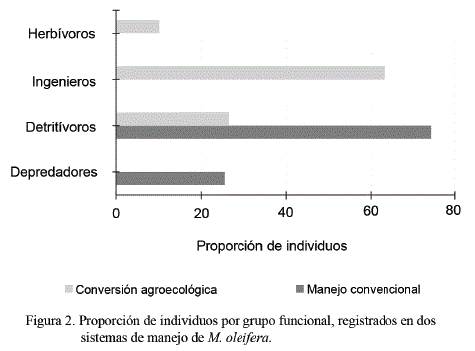
La distribución de familias por grupo funcional fue diferente entre sistemas de manejo, con predominancia de ingenieros del suelo (64,22 %) y detritívoros (27,37 %) en conversión agroecológica; y de detritívoros (74,19 %) y depredadores (25,81 %) en manejo convencional (fig. 2).
Matienzo-Brito et al. (2015) señalaron que el número de grupos funcionales difiere debido a la complejidad en la composición de los ecosistemas, con ventaja para los sistemas diversificados con manejo de biota auxiliar; ello permite explicar la presencia de grupos con funciones de acumulación y transformación de la biomasa, como son los ingenieros del suelo y los detritívoros. En este mismo sentido, Díaz-Porres et al. (2014) concluyeron que las condiciones que más influyen en la diversidad y funcionalidad de los grupos de la macrofauna edáfica son el contenido de materia orgánica y la relación carbono/nitrógeno que contiene el suelo.
Se ha reportado asociación entre la predominancia de detritívoros con el uso poco intensivo del suelo y, por tanto, con un buen contenido de materia orgánica (fig. 2), lo que contribuye complementariamente a la función de los ingenieros del suelo, y confirma que la diferencia en la composición de los grupos funcionales estuvo asociada a las condiciones del hábitat.
La proporción de detritívoros confirmó el contenido de materia orgánica registrado en el análisis de suelo (tabla 1) para ambos sistemas, así como su calidad en cuanto al contenido de N y el estado de descomposición, ya que especies representantes de familias Hydrobiidae, Hydrophilidae y Physidae son indicadores de sitios con sedimentos en descomposición (estiércol, excretas de caracoles y materia vegetal en descomposición).
La humedad del suelo (tabla 1) fue una condición que, según Cabrera-Dávila et al. (2011) y Matienzo-Brito et al. (2015), influyó en la presencia de ciertos organismos. Este factor se asumió por la presencia de familias de caracoles, como Hydrophilidae, Planorbidae y Leptoceridae, en manejo convencional (tabla 2); y de coleópteros acuáticos, como Elmidae, en conversión agroecológica (tabla 2). Algunas especies presentan una distribución asociada a sitios inundados o en agua sin corriente y a ambientes lóticos; por lo que son útiles como indicadores de la humedad del suelo, del grado de descomposición de la materia orgánica y, por tanto, de los nutrientes disponibles en el sistema.
El efecto de una alta presencia de detritívoros se relaciona con sus actividades de alimentación, ya que, como parte de la red trófica en los agrosistemas, este grupo funcional hace más eficiente la movilidad y adquisición de nutrientes por las plantas; ello indirectamente influye en la presencia de insectos defoliadores y plagas de cultivos (Altieri y Nicholls, 2003), condición que no es deseable desde el punto de vista de la sanidad y productividad de los cultivos.
En sentido práctico, los detritívoros hacen eficiente la concentración de recursos expresados en biomasa, materia orgánica y, por tanto, nutrientes disponibles para las plantas, por lo que la concentración de recursos atrae más organismos potencialmente plagas a los sistemas; sin embargo, las actividades de diversificación, rotación de cultivos y promoción de enemigos naturales contribuyen a la disminución de plagas y, por tanto, a la sostenibilidad productiva de los sistemas.
Silva et al. (2012) concluyeron que la diversidad de depredadores, que en este estudio estuvo compuesta principalmente por las clases Arachnida (Fam. Salticidae, 23,39 %), Arthropoda (familia. Formicidae, 10, 55 %) y Chilopoda (familia. Scolopendridae, 2,42 %), indica disponibilidad de presas. Debido a su hábito epigeo, funcionalmente aportan a la regulación de poblaciones de especies potencialmente plagas, condición que también fue reportada por Díaz-Porres et al. (2014) en sistemas agrícolas, en comparación con sistemas naturalizados.
Se registró una importante población de arañas (Tailless whip scorpions) en el sistema conversión agroecológica, así como depredadores de la familia Formicidae, lo que influyó en una baja proporción de herbívoros, a pesar de haber una alta concentración de recursos (diversidad de leguminosas y arvenses, entre otras). Esto permite comprobar que la reducción de las poblaciones de plagas en sistemas agroecológicos es una consecuencia tanto de los cambios nutricionales inducidos en el cultivo por la fertilización orgánica, como del incremento de los controles naturales de las plagas (Altieri y Nicholls, 2003).
La notable incidencia de individuos de la familia Formicidae en conversión agroecológica se debió a la presencia de moringa, leguminosas y arvenses en floración; este grupo, denominado ingenieros del suelo, indica condiciones particulares, como el grado de perturbación del ecosistema y, a la vez, la interacción potencial entre los cultivos y los organismos.
Chávez-Suárez et al. (2016) plantearon que los individuos de Formicidae son organismos de notable diversidad específica, porque comúnmente se encuentran en alta densidad y constituyen indicadores útiles, debido a que experimentan respuestas rápidas frente a diferentes prácticas agrícolas.
Por su parte, Castro-Delgado et al. (2008) informaron que elevadas densidades de Formicidae (hormigas forrajeras) pueden mover la misma cantidad de suelo que las lombrices de tierra; ello explica el hecho de que, en los sistemas de manejo, Solenopsis sp. y Camponotus sp. transportaron detritos originados por la descomposición de la moringa y de las leguminosas al interior del suelo.
En cuanto a la interacción de la moringa con organismos representantes de Formicidae, la fenología de la planta, manifestada en dos profusas floraciones anuales, garantizó la presencia y funcionalidad de esta familia; lo que se evidenció en actividades como la defoliación, que presentó mayor incidencia sobre plantas jóvenes, y el forrajeo de flores en plantas adultas, principalmente en la época seca. También se observó la depredación de individuos de la familia Formicidae por arañas pequeñas de la familia Salticideae, con lo cual se identificó uno de los mecanismos de regulación de las poblaciones.
Como parte del grupo funcional ingenieros del suelo, la densidad de Termitidae en conversión agroecológica es muestra de las condiciones en las que se originó el sistema, en cuanto a la predominancia de vegetación arbustiva de barbecho forestal, lo que conllevó concentración de restos de ramas y otro material orgánico, propicio para la proliferación de estos organismos.
La interacción de organismos de la familia Termitidae con M. oleifera fue negativa, ya que Termitidae constituyó una plaga que rápidamente debilitó las plantas de diferentes edades y que conllevó la pudrición seca de ramas y tallos. Asociado al parasitismo de Termitidae sobre moringa, aparecieron larvas de Lepidoptera que ejercieron herbivorismo, además de Hydrobiidae que actuó como detritívoro.
La incorporación y existencia continua de hojarasca en el suelo en forma de residuos de cosecha incrementó la densidad de algunos órdenes de Formicidae, que los transportaron al interior del suelo; y en el proceso ocurrió depredación de otros organismos de la macrofauna, como Termitidae y larvas de Coleoptera. Otros depredadores, como Julidae y Scolopendridae, utilizaron las galerías para ejercer su función, los primeros en la trituración de restos vegetales (Chávez-Suárez et al., 2016).
Contrario a lo propuesto por Leyva-Rodríguez et al. (2012) y Cabrera-Dávila (2012), la humedad no influyó en la presencia de Oligochaeta, aun cuando se registró una baja abundancia de lombriz de tierra en conversión agroecológica; ello coincide con los valores reportados por Matienzo-Brito et al. (2015) en parcelas diversificadas, en sistemas de conversión de uso del suelo de convencional a agroforestal, que presentaron baja densidad al inicio y un evidente incremento a los 10 años de conversión. Individuos de este grupo funcional no se registraron en el sistema de manejo convencional.
Chávez-Suárez et al. (2016) argumentaron que la presencia de Coleoptera es importante, debido a que interviene en la trituración de los restos vegetales, por lo que es un indicador de acumulación de biomasa y materia orgánica. La presencia de herbívoros, principalmente larvas de Coleoptera (familia Escarabaeidae, Phyllophaga ssp.), estuvo asociada a anteriores usos agrícolas del suelo y, según Leyva-Rodríguez et al. (2012), a la incorporación de residuos y a la calidad de la hojarasca, como producto de la presencia de árboles y arbustos con altos niveles de proteína en los sistemas.
La interacción entre las larvas y adultos de Escarabaeidae con moringa en manejo convencional, y con moringa más leguminosas en conversión agroecológica, fue diferente entre los estados de desarrollo, ya que en estado larvario se alimentaron de las raíces de plantas jóvenes; mientras que en estado adulto la especie Cotinis mutabilis se alimentó del néctar de moringa y de las leguminosas, lo cual contribuyó al proceso de polinización.
Noctuidae, por estar representada por larvas u orugas que viven en el suelo y se alimentan de hojas, flores, frutos y en ocasiones de semillas, representó riesgos de afectación para el cultivo de moringa en la fase inicial de crecimiento; pero, por otro lado, algunas especies en estado adulto participan en los procesos de polinización de especies arbóreas acompañantes y, por tanto, atraen polinizadores al sistema.
CONCLUSIONES
El número de unidades taxonómicas, la densidad y la diversidad de la macrofauna edáfica constituyeron indicadores que permitieron distinguir entre el manejo agroecológico y la agricultura convencional de M. oleifera.
Asimismo, la alta proporción de individuos de los grupos funcionales ingenieros del suelo y detritívoros en conversión agroecológica resultó un indicador de la buena salud del suelo, así como de una notable dinámica de transformación físico-biológica.
En sistemas con manejo convencional, la carencia de individuos del grupo funcional ingenieros del suelo originó una lenta transformación físico-biológica, a pesar de la alta descomposición de materia orgánica realizada por la dominancia de detritívoros.
REFERENCIAS BIBLIOGRÁFICAS
1. Altieri, M. A. & Nicholls, Clara I. Soil fertility management and insect pests: harmonizing soil and plant health in agroecosystem. Soil Till. Res. 72 (2):203-211, 2003.
2. Altieri, M. A.; Nicholls, Clara I.; Henao, A. & Lana, M. A. Agroecology and design of climate change-resilient farming systems. Agron. Sustain. Dev. 35 (3):869-890, 2015.
3. Altieri, M. A.; Nicholls, Clara I. & Montalba, R. Technological approaches to sustainable agriculture at a crossroads: an agroecological perspective. Sustainability. 9 (3):349, 2017.
4. AOAC. Official methods of analysis. 13th ed. Washington: Association of Official Analytical Chemists. https://archive.org/strem/gov.law.aoac.methods. 1980/aoac.methods.1980#page/n1/mode/2up. [22/08/2015], 1980.
5. Ayuke, F. O.; Karanja, N. K.; Muya, E. M.; Musombi, B. K.; Mungatu, J. & Nyamasyo, G. H. N. Macrofauna diversity and abundance across different land use systems in Embu, Kenya. Trop. Subtrop. Agroecosyst. 11 (2):371-384, 2009.
6. Blanco-Lobaina, J.; Contino-Esquijerosa, Y.; Iglesias-Gómez, J. M.; Caballero-Grande, R.; Perera, C. E. & Funes, F. Indicadores para evaluar la reconversión agroecológica en unidades básicas de producción cooperativa. Agricultura orgánica. 19 (1):27-29, 2013.
7. Cabrera-Dávila, Grisel. La macrofauna edáfica como indicador biológico del estado de conservación/perturbación del suelo. Resultados obtenidos en Cuba. Pastos y Forrajes. 35 (4):349-363. 2012.
8. Cabrera-Dávila, Grisel. Manual práctico la macrofauna edáfica como indicador biológico de la calidad del suelo según resultados en Cuba. La Habana: Fundación Rufford, 2014.
9. Cabrera-Dávila, Grisel; Robaina, Nayla & Ponce De León, D. Composición funcional de la macrofauna edáfica en cuatro usos de la tierra en las provincias de Artemisa y Mayabeque, Cuba. Pastos y Forrajes. 34 (3):331-346, 2011.
10. Castner, J. L. Photographic atlas of entomology and guide to Insect Identification. Gainesville, USA: Feline Press, 2001.
11. Castro-Delgado, Silvia; Vergara-Cobian, Clorinda & Arellano-Ugarte, Consuelo. Distribución de la riqueza, composición taxonómica y grupos funcionales de hormigas del suelo a lo largo de un gradiente altitudinal en el Refugio de Vida Silvestre Laquipampa, Lambayeque-Perú. Ecología Aplicada. 7 (1-2):90-103, 2008.
12. Chávez-Suárez, Licet; Labrada-Hernández, Yakelín & Álvarez-Fonseca, A. Macrofauna del suelo en ecosistemas ganaderos de montaña en Guisa, Granma, Cuba. Pastos y Forrajes. 39 (3):111-115, 2016.
13. Cunha-Neto, F. V. da; Correia, Maria E. F.; Pereira, G. H. A.; Pereira, M. G. & Leles, P. S. dos S. Soil fauna as an indicator of soil quality in forest stands, pasture and secondary forest. Rev. Bras. Cienc. Solo. 36 (5):1407-1417, 2012.
14. Díaz-Porres, Mónica; Rionda, Macarena H.; Duhour, A. E. & Momo, F. R. Artrópodos del suelo: Relaciones entre la composición faunística y la intensificación agropecuaria. Ecologia Austral. 24 (3):327-334, 2014.
15. Gliessman, S. R. Agroecologia: plantando las raíces de la resistencia. Agroecologia. 8:19-26, 2013.
16. Gómez-Pamies, D. F.; Godoy, María C. & Coronel, J. M. Macrofauna edáfica en ecosistemas naturales y agroecosistemas de la eco-región Esteros del Iberá (Corrientes, Argentina). Cienc. Suelo, Argentina. 34 (1):43-56, 2016.
17. Hatt, S.; Artru, Sidonie; Brédart, D; Lassois, Ludivine; Francis, F.; Haubruge, É. et al. Towards sustainable food systems: the concept of agroecology and how it questions current research practices. A review. BASE. 20 (S1):215-224, 2016.
18. INETER. Boletín climático. Managua: Dirección General de Meteorología, Instituto Nicaragüense de Estudios Territoriales, 2015.
19. Lavelle, P.; Senapati, P. & Barros, E. Soil macrofauna. In: G. Schroth and F. L. Sinclair, eds. Trees, crops and soil fertility. Concepts and research methods. United Kingdom: CABI Publishing. p. 303-322, 2003.
20. Leyva-Rodríguez, S. L.; Baldoquin-Pagan, A.; Reyno-Manduley, Y. & Ayala-Yera, J. R. Influencia del suelo en la macrofauna edáfica en áreas de la región norte del municipio de Las Tunas. Innovación Tecnológica. 18 (4):1-13, 2012.
21. Matienzo-Brito, Y.; Alfonso-Simonetti, Janet; Vásquez-Moreno, L. L.; De-la-Masa-Arias, A. R.; Matamoros-Torres, M.; Díaz-Finalé, Yunaisy et al. Diversidad de grupos funcionales de la fauna edáfica y su relación con el diseño y manejo de tres sistemas de cultivos. Fitosanidad. 19 (1):45-55, 2015.
22. Navarrete-Segueda, A.; Vela-Correa. G.; López-Blanco, J. & Rodríguez-Gamiño. Ma. de Lourdes. Naturaleza y utilidad de los indicadores del suelo. ContactoS. 80:29-37, 2011.
23. Nicholls, Clara I. & Altieri, M. A. Suelos saludables, plantas saludables: la evidencia agroecológica. LEISA. 24 (2):6-8, 2008.
24. Nicholls, Clara I.; Altieri, M. A. & Vásquez, L. L. Agroecology: principles for the conversion and redesign of farming systems. J. Ecosys. Ecograph. S5:010, 2016.
25. Nieto-Gómez, Libia E.; Valencia-Trujillo, Francis L. & Giraldo-Díaz, R. Bases pluriepistemológicas de los estudios en agroecología. Entramado. 9 (1):204-211, 2013.
26. Rendón-Pareja, S.; Artunduaga-Lemus, F.; Ramírez-Pisco, R.; Quiroz-Gambia, J. A. & Leyva-Rojas, Edna I. Los macroinvertebrados como indicadores de la calidad del suelo en cultivos de mora, pasto y aguacate. Rev. Fac. Nac. Agron. Medellín. 64 (1):5793-5802, 2011.
27. Roldán-Pérez, G. Guía para el estudio de macroinvertebrados acuáticos del Departamento de Antioquia. Bogotá: FEN, Colciencias, Universidad de Antioquia, 1988.
28. Sheibani, S. & Gholamalizadeh, A. Effect of tillage on soil biodiversity. JNASCI. 2 (8):273-281, 2013.
29. Silva, J.; Jucksch, I. & Tavares, R. C. Invertebrados edáficos em diferentes sistemas de manejo do cafeeiro na Zona da Mata de Minas Gerais. Rev. Bras. Agroecol. 7 (2):112-125, 2012.
30. Socarrás, Ana & Izquierdo, I. Evaluation of agroecological systems through biological indicators of the soil quality: edaphic mesofauna. Pastos y Forrajes. 37 (1):109-114. https://payfo.ihatuey.cu/ index.php?journal=pasto&page=article&op=view&path%5B%5D=1782&path%5B%5D=2930, 2014.
31. Turner, Monica G. & Gardner, R. H., eds. Quantitative methods in landscapes ecology. The analysis and interpretation of landscape heterogeneity. New York: Springer-Verlag, 1991.
Recibido el 4 de octubre del 2016
Aceptado el 28 de septiembre del 2017


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}