










 Servicios personalizados
Servicios personalizados Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los pastos y los forrajes son la principal fuente de alimento para los rumiantes y representan el mayor volumen de la dieta, ya que son más baratos, tienen una gran capacidad de producción y crecen fácilmente (Lee et al., 2017). Por otro lado, el consumo de nutrientes es uno de los principales factores que restringe la producción animal en el trópico y solo se puede controlar si el valor nutricional de los forrajes no constituye un factor limitante (Olafadehan y Okunade, 2018).
La producción ganadera depende, en gran medida, de los factores climáticos y meteorológicos; por ello, la transición anual de las condiciones climáticas, fundamentalmente en cuanto a la temperatura, la humedad relativa y las precipitaciones, genera la correspondiente variabilidad en la producción y el valor nutritivo de los forrajes. En tal sentido, la ganadería tropical basada en el pastoreo de gramíneas en monocultivo enfrenta grandes desafíos, especialmente en la época de sequía, durante la cual disminuyen de manera drástica la disponibilidad y la calidad de los forrajes (Cuartas-Cardona et al., 2014).
Por otra parte, la determinación de la capacidad de producción actual de los pastos, junto a la presión en relación con el uso de la tierra para la producción ganadera, se ha convertido en uno de los principales desafíos para desarrollar la planificación integrada y la toma de decisiones en los sistemas tropicales de pastoreo (Crestani et al., 2013).
Ello explica la necesidad de adoptar sistemas sostenibles de producción ganadera que aprovechen las ventajas del manejo integrado en el contexto biofísico del neotrópico, donde los paisajes naturales y los bosques mixtos se usan incorrectamente como sistemas de pastoreo extensivo. Por tanto, la conversión ambiental basada en el silvopastoreo constituye una alternativa promisoria para enfrentar estos problemas (Murgueitio-Restrepo et al., 2011), ya que los sistemas con árboles son capaces de conservar la biodiversidad (Schindler et al., 2016), aportar servicios ambientales a los ecosistemas (Martínez-Pastur et al., 2017), incrementar tanto cualitativa como cuantitativamente la oferta de forraje al ganado (Carvalho et al., 2017), así como minimizar el desbalance en la producción de alimentos que caracteriza a los sistemas sin árboles (Murgueitio-Restrepo et al., 2016).
La composición química y la morfología de los forrajes determinan la palatabilidad y el valor nutricional para el ganado, por lo cual influyen en la cantidad de alimento que se consume, la eficiencia de la rumia, la tasa de ganancia de peso, el volumen y la calidad de la leche que se produce, y en el éxito reproductivo (Herrero et al., 2015). De ahí que, el objetivo del estudio fue evaluar el efecto de la época del año en la producción y el valor nutritivo de los forrajes, en una asociación de Megathyrsus maximus con Leucaena leucocephala.
Materiales y Métodos
Ubicación del área experimental. La investigación se desarrolló durante cinco años en condiciones de producción, en el área de pastoreo de la vaquería 66, perteneciente a la Empresa Pecuaria Genética de Matanzas -Cuba-, la cual se ubica geográficamente en los 22º 58ʹ 39ʺ de latitud norte y los 81º 29ʹ 55,66ʺ de longitud oeste, a 100 msnm (Academia de Ciencias de Cuba, 1989).
Características edáfoclimáticas. El suelo se clasifica como Pardo con Carbonatos (Hernández-Jiménez et al., 2015), con relieve ondulado. El clima es cálido tropical (Centro del Clima, 2018). Los valores de las variables climatológicas en la etapa experimental se presentan en la tabla 1.
Tabla 1 Comportamiento de las variables climatológicas en el período experimental.
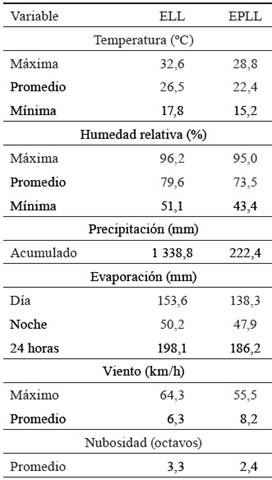
ELL: época lluviosa, EPLL: época poco lluviosa.
Descripción del área de pastoreo y el manejo.
El área de pastoreo (42 ha) fue dividida en 36 cuartones de aproximadamente 1,2 ha cada uno. Como gramínea mejorada, predominó M. maximus cv. Likoni; mientras que la leguminosa arbórea del sistema fue L. leucocephala cv. Cunningham, que tenía 10 años de establecida y se sembró a una distancia entre surcos de 5 m, con densidad promedio de 553 plantas/hectárea. Para el manejo con los animales, la superficie de pastoreo se fraccionó en tres áreas de 12 cuartones cada una; el tiempo de ocupación fue de 3,0-3,5 días en la ELL y de 4,5-5,0 días en la EPLL, lo que permitió garantizar tiempos de reposo al pasto de 33-39 días y de 50-55 días para la ELL y la EPLL, respectivamente. La carga promedio utilizada fue de 2,0 UGM/ha.
Las plantas de L. leucocephala se podaron en el período de febrero-abril, a 1,7 m de altura, en una cantidad aproximada de 15 árboles por día de ocupación en cada rotación comprendida en el período de poda.
Mediciones en la comunidad vegetal
Composición florística. Se realizó semestralmente en mayo y noviembre, por el método de los pasos descrito por EEPFIH (1980).
Densidad de plantas de L. leucocephala. Se estimó con una frecuencia semestral (mayo y noviembre). Para ello, se contó la cantidad de plantas de L. leucocephala en tres surcos característicos de cada cuartón, se halló el promedio entre ellos, el resultado se multiplicó por la cantidad de surcos en el cuartón y este valor se dividió entre el área.
Estimación de la disponibilidad para ramoneo en L. leucocephala. Con frecuencia mensual, se recolectaron manualmente las hojas y los tallos tiernos comestibles (hasta aproximadamente 3 mm de diámetro) en 10 de los árboles establecidos en cada cuartón, simulando el ramoneo que realizan los animales hasta una altura aproximada de 2 m.
Disponibilidad del pasto. Se realizó mensualmente, a través del método alternativo propuesto por Martínez et al. (1990) a la entrada y a la salida de los animales del cuartón; y se efectuaron, como promedio, 80 observaciones por hectárea.
Análisis químico proximal de los alimentos. Con frecuencia mensual, se envió al laboratorio una muestra homogénea de 300 g de follaje de L. leucocephala, 300 g de la biomasa comestible de M. maximus y 300 g de forraje de Neonotonia wightii, cosechados según la metodología propuesta por Herrera (2006). Se midieron los siguientes indicadores: materia seca (MS), ceniza, materia orgánica (MO), proteína bruta (PB) y extracto etéreo (EE), según las técnicas descritas por la AOAC (1990). Las fracciones fibrosas -fibra en detergente neutro (FDN), fibra en detergente ácido (FDA)- y la lignina se analizaron mediante los procedimientos referidos por Van Soest et al. (1991), sin corrección por cenizas, usando bolsas de filtro ANKOM F-57 en un analizador de fibras ANKOM200 (ANKOM Technology Corporation, Fairport, NY, USA). La hemicelulosa y la celulosa se calcularon mediante la diferencia entre la FDN, la FDA y la lignina. El nitrógeno ligado a la FDN (N-FDN) se estimó de acuerdo con los procedimientos propuestos por Licitra et al. (1996).
Degradabilidad in vitro de la MS de los alimentos. Se estimó a las 96 h de la fermentación, a través de la técnica de producción de gas (PG) in vitro propuesta por Theodorou et al. (1994).
Estimación de la energía metabolizable (EM). La EM (MJ/kg MS) se estimó de acuerdo con Menke y Steingass (1988):
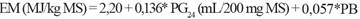
Procedimiento estadístico. Para las variables correspondientes a la composición florística se comprobó, mediante el programa Statistica, el cumplimiento de los supuestos teóricos del análisis de varianza: la homogeneidad de la varianza por la dócima de Levene (1960) y la normalidad de los errores por la dócima de Shapiro y Wilk (1965). Al evidenciarse que no se cumplieron dichos supuestos, se aplicó la transformación ArcoSeno √%, que tampoco posibilitó su cumplimiento.
Por ello, con el programa estadístico InfoStat (Di Rienzo et al., 2012) se realizó un análisis no paramétrico (Kruskal-Wallis), y se utilizó la dócima de Conover (1999) para la comparación de los rangos medios. Para las variables propias de la disponibilidad de MS y PB de la gramínea y de la leucaena, así como para los datos correspondientes al valor nutricional de los alimentos de la dieta, se realizó análisis de varianza según modelo lineal general. En el modelo se tuvo en cuenta el efecto época, y se aplicó la dócima de LSD-Fisher para p < 0,05.
Resultados y Discusión
En la tabla 2 se presentan los resultados de la composición florística del pastizal por época del año. Se constató que, en ambas épocas, aproximadamente el 70 % del área estuvo cubierta por pastos mejorados (M. maximus, Cynodon nlemfuensis Vanderyst y Urochloa mutica (Forssk.) T.Q.Nguyen), los cuales a su vez estuvieron representados en más de un 70 % por la especie M. maximus; mientras que los pastos naturales ocuparon entre el 14,0 y el 17,8 % del área de pastoreo, sin diferencias entre las épocas del año, y las principales especies estuvieron representadas por el complejo Dichanthium annulatum (Forssk.) Stapf - Dichanthium caricosum (L.) A. Camus, Paspalum notatum Alain ex Flüggé, Cynodon dactylon (L.) Pers., Hyparrhenia rufa (Nees) Stapf, Sporobolus indicus (L.) R. Br. y Phylla strigillosa.
Tabla 2 Composición florística del pastizal por época del año*.

* Rango medio ± desviación estándar (media aritmética expresada en %), ELL: época lluviosa, EPLL: época poco lluviosa, PM: pastos mejorados, PA: plantas arvenses, LV: leguminosas volubles.
Las leguminosas volubles, entre las cuales N. wightii representó más del 80 %, surgieron de forma natural en el sistema y tuvieron una presencia significativamente superior (p = 0,009) en la ELL respecto a la EPLL. Además, se encontró que los valores fueron ligeramente superiores a los informados por Lamela et al. (2009) en un sistema silvopastoril (asociación de L. leucocephala con C. nlemfuensis), en el cual las leguminosas volubles igualmente aparecieron de forma espontánea (3,0-4,0 %).
Las plantas arvenses (Acacia farnesiana (L.) Willd., Dichrostachys cinerea (L.) Wight & Arn., Mimosa pudica L. y Ricinus communis L., entre otras) tuvieron una presencia relativamente baja en el sistema, con valores similares para ambas épocas (6,4 y 5,6 % para la ELL y la EPLL, respectivamente); de igual forma, las zonas despobladas fueron escasas (0,01-0,49 %) y no manifestaron diferencias en cuanto a la representación en el área entre las épocas del año.
Los pastos mejorados más las leguminosas volubles, que constituyen la mayor fracción de la dieta y presentan el mejor valor nutricional con respecto al resto de los alimentos que consumen los animales en pastoreo, representaron de conjunto entre el 76,1 y el 79,6 % del total del área de pasto; mientras que la superficie que ocuparon las plantas arvenses más las zonas despobladas, que constituye el área que no produce alimento de forma directa para los animales, significó entre el 6,1 y el 6,4 % del área de pastizal.
La disponibilidad de las gramíneas (tabla 3) fue alta en ambas épocas del año, aunque manifestó diferencias significativas (p < 0,001) a favor de la ELL. Por el contrario, la disponibilidad de follaje en L. leucocephala fue relativamente baja, pero similar en las dos épocas (85,7 y 72,1 kg de MS/ha/rotación para la ELL y la EPLL, respectivamente). No obstante, estos valores son superiores al informado por Sánchez-Santana et al. (2016) para una asociación de M. maximus y L. leucocephala, donde la disponibilidad de forraje de la leñosa, con densidad de 236-364 plantas/ha, fue solo de 13,3 kg de MS/ha/rotación.
Tabla 3 Disponibilidad de MS (kg MS/ha/rotación) y PB (kg PB/ha/rotación) de los forrajes del SSP.
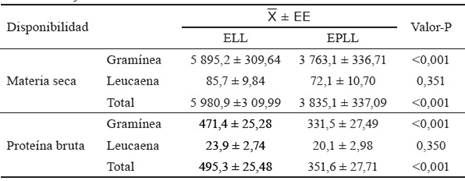
ELL: época lluviosa, EPLL: época poco lluviosa.
Por otra parte, la disponibilidad de biomasa total del sistema resultó elevada en ambas épocas del año; no obstante, presentó el mismo comportamiento que la disponibilidad de la gramínea, con diferencias significativas (p < 0,001) a favor de la ELL.
Es necesario destacar que, aun cuando las precipitaciones durante la EPLL representaron solo el 14,3 % del total anual, la disponibilidad de MS en esta época significó aproximadamente el 31 % de la disponibilidad anual, por lo que el sistema contribuyó a lograr un mejor balance en la producción de alimentos entre ambas épocas del año al compararse con los sistemas sin árboles, donde la biomasa disponible en la EPLL puede llegar a representar menos del 20 % con respecto a la disponibilidad de MS anual. Estos resultados evidencian la importancia de asociar las gramíneas mejoradas con plantas arbóreas, principalmente leguminosas (L. leucocephala), ya que permiten no solo incrementar la producción de biomasa comestible del sistema, sino también garantizar una mayor estabilidad productiva entre las épocas del año (Sánchez-Santana et al., 2018).
La disponibilidad de PB de la gramínea fue alta en ambas épocas del año, pero significativamente superior (p < 0,001) en la ELL debido a la mayor disponibilidad de MS que manifestó el pasto en esa época. En cambio, la disponibilidad de PB de L. leucocephala fue baja y similar para las dos épocas. Ello determinó que la disponibilidad de proteína bruta total siguiera la misma tendencia que la disponibilidad de PB de la gramínea, con diferencias significativas (p < 0,001) favorables a la ELL.
Los valores de disponibilidad de PB del sistema en ambas épocas del año propiciaron que se alcanzaran 3 501,6 kg de PB/ha/año, resultado satisfactorio para las condiciones de producción de Cuba, aunque inferior (4 100,0 kg) al informado por Bacab et al.( 2013) en un sistema silvopastoril intensivo constituido por una asociación de L. leucocephala (10 000 plantas/ha) y Cynodon plectostachyus.
Los indicadores del valor nutricional de M. maximus por época del año se presentan en la tabla 4. El contenido de MO fue óptimo y presentó valores superiores (p = 0,044) en la ELL. La DIVMS a las 96 h fue elevada y mostró el mismo comportamiento en ambas épocas del año, con valores promedio de 655,1 y 649,5 g/kg de MS para la ELL y la EPLL, respectivamente, que están en el rango recomendado por Arango et al. (2016) para esta especie (600-700 g/kg de MS). Además, dichos valores son similares al informado por Santiago-Hernández et al. (2016) en un sistema de M. maximus vcs. Mombaza y Tanzania (657,0 g/kg de MS) asociados con Melia azedarach L., sembrada con una densidad de 1 000 plantas por hectárea. Sin embargo, son ligeramente superiores a los obtenidos por Frota et al. (2017) en M. maximus vc. Mombaza en asociación con Attalea speciosa Mart. (629,9 y 609,8 g/kg de MS para la ELL y la EPLL, respectivamente).
Tabla 4 Indicadores del valor nutricional de M. maximus.

ELL: época lluviosa, EPLL: época poco lluviosa.
La EM en el forraje de M. maximus fue relativamente baja en las dos épocas del año y mostró un valor superior (p < 0,001) en la ELL, similar al informado por Heuzé et al. (2015), estimado mediante la técnica de producción de gas in vitro (7,9 MJ/kg de MS). El contenido de PB fue significativamente superior (p = 0,004) en la EPLL con respecto a la ELL, lo que pudo estar asociado a la disminución en la intensidad de luz solar en esa época del año y a la capacidad de M. maximus de tolerar condiciones limitantes de luz. Según Santiago-Hernández et al. (2016), en tales condiciones esta especie cambia su fisiología, con lo cual disminuye la producción de biomasa, pero mejora su calidad nutricional.
El contenido de PB en la biomasa comestible de M. maximus, en ambas épocas del año, fue superior al descrito para las gramíneas tropicales en monocultivo (Gaviria et al., 2015), lo que se debe a la asociación de M. maximus con los árboles (Silva-Parra et al., 2018), en este caso L. leucocephala, leguminosa capaz de fijar nitrógeno atmosférico al suelo (Conrad et al., 2018) y con ello contribuir a mejorar la calidad nutricional de los pastos que se encuentran asociados a ella. Los valores para la ELL (11,5 %) fueron similares a los informados para esta gramínea, tanto en un SSP con igual densidad de L. leucocephala y equivalente régimen de precipitaciones (Sánchez-Santana et al., 2016), como en otro con alta densidad de esta leñosa y un régimen de precipitaciones bimodal (Molina et al., 2015).
En el caso del N-FDN se encontraron valores de 43,7-45,6 %, similares a lo informado por Montalvão-Lima et al. (2018) para la biomasa comestible de M. maximus (45,0 %) en monocultivo, con fertilización (20 kg de P2O5/ha, 60 kg de K2O/ha, y 70 kg de N/ha), en un suelo clasificado como Oxisol, y a una altitud de 1 014 msnm.
El contenido de FDN y FDA en la biomasa comestible de M. maximus no difirió entre las épocas del año, lo que implica que la edad fenológica de las plantas era análoga aun cuando la edad cronológica era diferente (33-39 y 55-60 días para la ELL y la EPLL, respectivamente). Estos valores son inferiores a los informados por Rodríguez y Lara (2018) en M. maximus cv. Tanzania (74,9-80,7 y 41,9-57,6 % para FDN y FDA, respectivamente) en un sistema silvopastoril intensivo constituido por una asociación de L. leucocephala (34 000 plantas/ha) y M. maximus.
El contenido de hemicelulosa fue similar al informado por Patiño-Pardo et al. (2018) para el forraje de M. maximus cv. Tanzania, cosechado entre 35 y 45 días en la subregión de Sabana en el Departamento de Sucre -Colombia.
Con respecto a L. leucocephala (tabla 5), el contenido de MO fue adecuado y con valores similares en las dos épocas del año. Por otra parte, la DIVMS96 mostró un valor superior (p = 0,003) en la EPLL, lo que pudo deberse al menor contenido de lignina en el follaje durante esa época del año. Este indicador estuvo en el rango informado para el follaje de esta planta por Pal et al. (2015) y Rivera-Herrera et al. (2017); sin embargo, fue superior al obtenido tanto por Hernández-Morales et al. (2018) en hojas de L. leucocephala (489,7 g/kg de MS) en México, como por Vivas-Arturo et al. (2017) en la biomasa comestible de esta planta (412,0 g/kg de MS) en Colombia. En este último caso, el bajo valor de la digestibilidad podría estar asociado a la baja proporción de hojas con respecto a los tallos en la biomasa comestible total.
Tabla 5 Indicadores del valor nutricional de L. leucocephala.
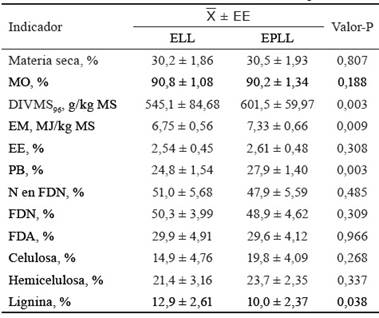
ELL: época lluviosa, EPLL: época poco lluviosa.
El valor de EM fue superior (p = 0,009) en la EPLL en relación con la ELL. Estos datos superan el valor informado por Rodríguez et al. (2014) en la harina del follaje de L. leucocephala (6,3 MJ/kg de MS); sin embargo, son inferiores a los 8,4 MJ/kg de MS informados por Heuzé y Tran (2015) para la biomasa comestible de esta planta, en ambos casos estimados mediante producción de gas in vitro.
El contenido de PB en el follaje de L. leucocephala fue superior (p = 0,003) en la EPLL con respecto a la ELL; sin embargo, dicho indicador presentó valores adecuados en ambas épocas del año (24,8 y 27,9 % para la ELL y la EPLL, respectivamente), los cuales son similares a los informados por Singh et al. (2014) y Cuartas-Cardona et al. (2015). Estos resultados demuestran la importancia de L. leucocephala para incrementar el contenido de PB de la dieta ofrecida a los animales en pastoreo, debido a que en tales condiciones las leguminosas aportan más proteína que las gramíneas (Carvalho et al., 2017).
Por otra parte, el follaje de las leguminosas arbóreas (como L. leucocephala) no solo es importante por el aporte directo de PB, sino también por la presencia de taninos condensados; estos se unen a la proteína de la dieta para formar un complejo tanino-proteína que puede insolubilizar las proteínas y de esa forma evitar su degradación en el rumen, lo que aumenta el sobrepaso de proteína hacia el tracto gastrointestinal inferior. Ello trae consigo un elevado contenido de proteínas en el intestino delgado, que, cuando se digieren incrementan la absorción de aminoácidos (Wanapat et al., 2015).
El contenido de FDN y FDA en el follaje de L. leucocephala fue similar en ambas épocas del año, y los valores fueron inferiores a los informados por Piñeiro-Vázquez et al. (2017) para la biomasa comestible de esta planta en Yucatán, México (58,1 y 42,0 % para la FDN y la FDA, respectivamente). No obstante, el contenido de FDN fue superior al informado por Worknesh y Getachew (2018) en la biomasa comestible de L. leucocephala (34 %), cosechada con una frecuencia de corte de 45 días, en Etiopía.
El contenido de lignina en la biomasa comestible de L. leucocephala fue superior (p = 0,038) en la ELL, quizás como consecuencia de una mayor edad de los rebrotes, debido a un menor consumo por los animales durante la época del año en la que el follaje muestra el menor valor nutricional. El contenido de lignina en el follaje de L. leucocephala en la ELL fue similar al informado por Carvalho et al. (2017) en la biomasa comestible de esta leguminosa arbórea cultivada sobre un suelo latosólico rojo-amarillento, durante la ELL, en un sistema en asociación con pastos mejorados para la alimentación de cabras, en el Estado de Piauí -Brasil.
Por el contrario, el contenido de lignina que mostró el follaje de esta planta en la EPLL fue similar al informado por Worknesh y Getachew (2018) en plantas de L. leucocephala, cosechadas cada 45 días, en Etiopía (10,6 %).
El análisis de los indicadores del valor nutricional del forraje de N. wightii (tabla 6) mostró que en todos los casos hubo un comportamiento estadísticamente similar en las dos épocas del año.
Tabla 6 Indicadores del valor nutricional de N. wightii.

ELL: época lluviosa, EPLL: época poco lluviosa.
La DIVMS a las 96 h fue elevada, con valores superiores a los 640 g/kg de MS en las dos épocas; ello resulta similar a lo informado por Valarini y Possenti (2006) para la biomasa comestible de N. wightii (640 g/kg de MS), cosechada en Brasil en un sistema con precipitación anual de 1 185 mm.
El contenido de PB (18,1-20,6 %) y la concentración de FDN (48,8-50,1 %) fueron similares a los valores informados por Carter et al. (2015) para el forraje de N. wightii (19,3 y 49,7 %, respectivamente) en Uganda. Sin embargo, la EM para rumiantes (7,40-7,67 MJ/kg de MS) -calculada mediante la técnica de producción de gas in vitro- fue inferior a la informada por Heuzé et al. (2015) para el forraje de N. wightii (8,10 MJ/kg de MS), estimada mediante ecuaciones.
Los valores de FDN, FDA y lignina fueron similares a los informados por Verdecia et al. (2017) en el forraje de N. wightii, cosechado con una frecuencia entre 30 y 45 días en la ELL (49,7-52,9; 27,5-29,4 y 5,8-7,2 %, respectivamente), en un sistema de monocultivo sobre suelo Pardo con Carbonatos, en la provincia Granma -Cuba.
Conclusiones
La asociación de gramíneas mejoradas con L. leucocephala, en condiciones de producción, mostró una elevada disponibilidad de biomasa total y de PB por hectárea; la cual, aun cuando fue superior en la época lluviosa, demostró el potencial de estos sistemas para garantizar una adecuada producción de alimento durante todo el año.
Además, la biomasa comestible de M. maximus presentó, en las dos épocas, un adecuado valor nutricional, con mayor concentración de EM en la ELL y un contenido de PB más elevado en la EPLL. Sin embargo, el follaje de L. leucocephala tuvo el mejor valor nutricional en la EPLL, fundamentalmente en cuanto a EM y DIVMS96; mientras que el forraje de N. wightii mostró excelentes indicadores del valor nutricional durante todo el año, por lo que la biomasa comestible de ambas leguminosas contribuye a complementar apropiadamente la dieta potencial que representa el pasto en ambas épocas.

