










 Servicios personalizados
Servicios personalizados Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La degradación del suelo es uno de los efectos más graves de la crisis de recursos naturales a diferentes escalas. El sector agropecuario se cataloga como insostenible, debido a que el modelo ganadero convencional provoca pérdidas de la biodiversidad y sobreexplotación de los recursos naturales (Ramírez y Fernando, 2014).
Huybrechs et al. (2015), al realizar estudios en Nicaragua informaron que, en los sistemas ganaderos, la capa del suelo se degrada producto de la acción del pisoteo de los animales y por el pastoreo intensivo, que erosiona la capa superficial del suelo, disminuye la fertilidad natural y las comunidades bióticas presentes.
El funcionamiento de los agrosistemas depende de los diseños espaciales y temporales que promueven sinergias entre los componentes de la biodiversidad del suelo, que condicionan procesos ecológicos claves, como la regulación biótica, el reciclaje de nutrientes y la productividad (Montagnini y Nair, 2014).
Según Cabrera-Dávila (2012), la diversidad biológica presente en el suelo tiene diferentes funciones, que contribuyen a mejorar su composición química y física. De manera particular, la macrofauna edáfica interviene en numerosos procesos, promoviendo o afectando el crecimiento de la vegetación, así como la productividad (Masin et al., 2017).
En un estudio de Chávez-Suárez et al. (2016) se ha destacado la importancia de algunas clases y órdenes en la transformación del suelo (Annelida: Oligochaeta), la formación de poros (Insecta: Hymenoptera, e Isopoda) y la trituración de restos vegetales (Coleoptera, Diplopoda, Isopoda, Gastropoda). Entender las funciones de la macrofauna edáfica permite determinar su aporte a la sostenibilidad, lo que la convierte en un indicador importante a nivel de sistema. Chávez-Suárez et al. (2016) enfatizaron en la utilidad de la macrofauna en la evaluación del estado de conservación o perturbación del suelo.
De acuerdo con Ramírez-Suárez et al. (2018), en los sistemas ganaderos, el conocimiento de la biota edáfica reviste particular interés, debido a las funciones que esta desempeña en subsistemas que tienen que ver con la alimentación de ganado vacuno, donde participa en la descomposición de hojarasca, la mineralización de la materia orgánica y los cambios estructurales del suelo.
Los beneficios de los sistemas silvopastoriles hacia el recurso suelo han sido abordados por Murgueitio et al. (2013), en términos del incremento en el contenido de carbono, la retención de humedad, el reciclaje de nutrientes y la activación de la cobertura vegetal y la diversificación. Esto se traduce, a su vez, en el aumento de la eficiencia simbiótica de la macrofauna edáfica, que permite establecer tendencias acerca del potencial productivo del hato lechero y cárnico (Rosero et al., 2018). Este estudio tuvo como objetivo caracterizar la composición trófica de la macrofauna del suelo en dos sistemas ganaderos, pertenecientes a cuatro municipios, ubicados en el Corredor Seco de Nicaragua.
Materiales y Métodos
Localización geográfica de los sitios de estudio. La investigación se desarrolló entre junio 2017 y febrero 2018, período que representó la época óptima para el establecimiento de los bancos de proteína, al corresponderse con la etapa de lluvias (mayo a noviembre). El área de estudio comprendió a los municipios de Rivas, Belén, La Conquista y Diriamba, todos ubicados en el Corredor Seco de Nicaragua. En cada municipio se evaluaron cuatro fincas: dos con sistemas ganaderos convencionales y dos con sistemas silvopastoriles (SSP) de bancos de proteína, destinados al corte y acarreo. Se exceptuaron Rivas y Belén, donde solo hubo una finca de cada sistema (tabla 1).

Características edafoclimáticas de las áreas evaluadas. Según datos de INETER (2017), las temperaturas oscilaron entre 32 y 35 °C, y las precipitaciones anuales, entre 1 500 y 1 750 mm. Rivas y Belén fueron los municipios que registraron menores temperaturas y precipitaciones.
En cada uno de los sitios evaluados, se realizó un muestreo al azar en las parcelas a fin de tomar muestras de la heterogeneidad de condiciones en el suelo. Los sistemas silvopastoriles se encuentran ubicados en fincas ganaderas, donde se trabaja con vacas adultas lactantes, de las razas Pardo suizo y Brahman en pastoreo. Como estrato herbáceo predominan las gramíneas: Andropogon gayanus Kunth, Hyparrhenia rufa (Nees) Stapf y Dichantium aristatum (Benth). Además, se dispone de bancos de proteína de 360 m2 (12 x 30 m). De las especies Leucaena leucacephala (Lam.), Cratylia argentea (Desv.) o Kuntze, Gliricidia sepium (Jacq), Guazuma ulmifolia (Lam.) o Moringa oleifera (Lam.), al menos una, establecida a una distancia de 1 m entre plantas e hileras. Al momento de la siembra, se realizó fertilización orgánica con 0,5 kg de compost en cada postura. Esta fertilización se repitió a los 35 y 75 días después del establecimiento. Luego de la siembra, a los 60 días, se hizo control manual de las principales arvenses presentes (Cyperus rotundus L, Euphorbia graminea Jacq, Mimosa pudica L y Sida acuta Burm) y un corte de uniformidad, a una altura de 60 cm sobre el suelo.
Los SSP no recibieron irrigación. Para efectos de manejo, se consideraron plenamente establecidos ocho meses después de su siembra. Se manejaron con el enfoque de corte y acarreo. La biomasa se cortó a 60 cm sobre el suelo, cada dos meses, para suministrar el pasto a los animales en forma fresca.
Los sistemas convencionales consisten en fincas ganaderas, en las cuales la alimentación animal depende principalmente del pastoreo, en áreas con no menos de tres años de establecimiento y porcentajes de cobertura no inferiores al 80 %. Los pastos predominantes son Andropogon gayanus Kunth, Hyparrhenia rufa (Nees) Stapf y Dichantium aristatum (Benth). El hato estuvo compuesto por vacas en ordeño, de las razas Pardo Suizo y Brahman. El pastoreo siguió un calendario de ocho días de ocupación y 22 días de descanso en cada potrero. Estos sistemas convencionales no recibieron riego ni fertilización. La carga animal media en los sistemas fue de 0,7 UGM/ha. Las características del suelo en las áreas evaluadas se muestran en la tabla 2.
Tabla 2 Propiedades físico-químicas del suelo en sistemas ganaderos.
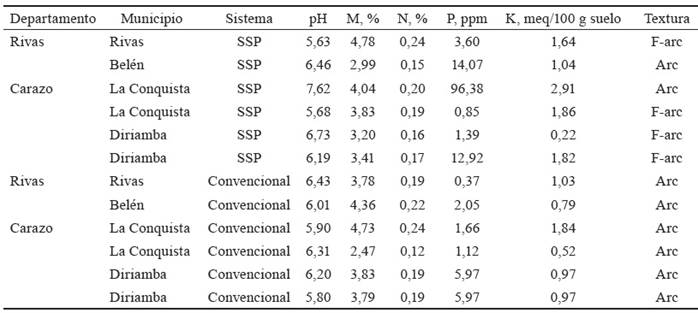
F-arc: Franco arcilloso; Arc: Arcilloso, pH; MO: materia orgánica; N: nitrógeno; P: fósforo; meq: mili equivalente; ppm: partes por millón.
Recolecta y procesamiento de la macrofauna edáfica. La recolección de muestras de la macrofauna edáfica se realizó entre el 10 de junio del 2017 y el 20 febrero del 2018. Las diferencias entre épocas climáticas no fueron objeto de estudio, sino el acumulado durante el ensayo. Los muestreos se realizaron cada 60 días. Se empleó la metodología propuesta por el Programa Internacional Biología y Fertilidad del Suelo Tropical (Lavelle et al., 2003). De cada sistema se extrajeron dos monolitos de suelo para la determinación de las propiedades físico-químicas. El muestreo de la macrofauna edáfica se realizó a tres profundidades (hojarasca, 0-10 cm y 10-30 cm), considerando que es en ellas donde existe mayor actividad biológica (Castner, 2001).
Los especímenes colectados se depositaron en viales con alcohol al 70 % para su traslado al Laboratorio de Entomología de la Facultad de Agronomía de la Universidad Nacional Agraria, donde se realizó su identificación hasta la categoría taxonómica de familia. Se aplicaron para ello las claves propuestas por Castner (2001), Cabrera-Dávila et al. (2011), Roldan (1988), y Cabrera-Dávila (2014). Las poblaciones y dominancia de la macrofauna del suelo se obtuvieron a partir de la transformación del número de individuos por monolito en número de individuos por metro cuadrado (ind/m2). Asimismo, se le asignó la función ecológica a cada familia. Los individuos se clasificaron como detritívoros, ingenieros del suelo, herbívoros y depredadores (Cabrera-Dávila et al., 2011).
Resultados y Discusión
La composición de las familias fue diferente entre sistemas, con mayor riqueza en el SSP (31 familias), en comparación con el convencional (23 familias). En ambos, la familia Formicidae, perteneciente al gremio ingenieros del suelo, fue la más representativa (tabla 3).
Tabla 3 Clasificación taxonómica, rol trófico y diversidad de la macrofauna edáfica por sistemas ganaderos.
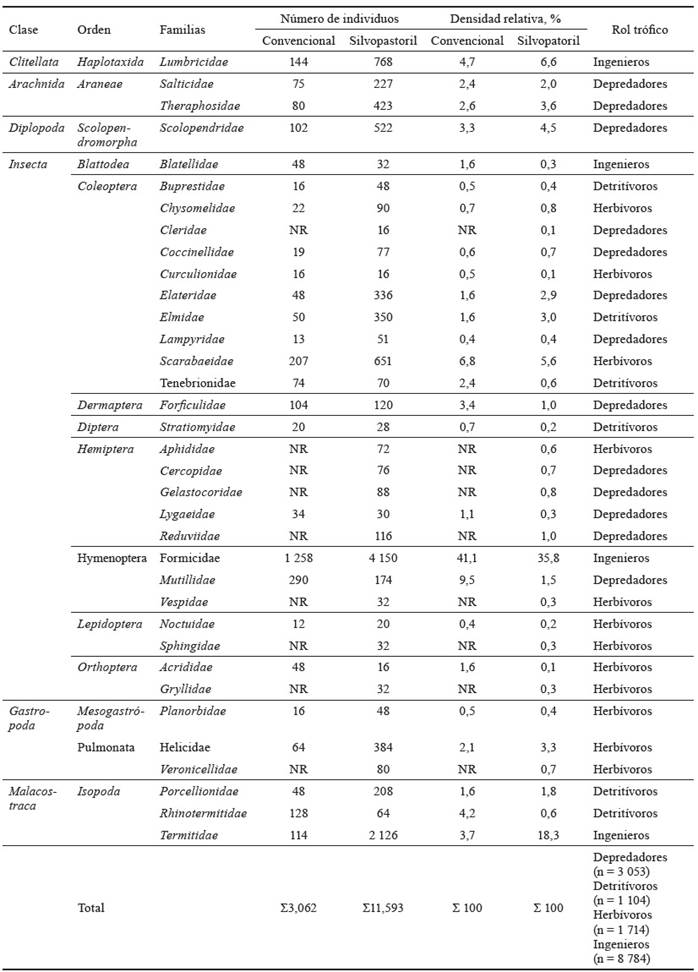
NR (No registrado)
Las familias asociadas a organismos detritívoros presentaron bajas poblaciones (ind/m2), siendo Porcellionidae común entre sistemas, con la mayor densidad con respecto a otras familias de detritívoros. La composición de familias del grupo de herbívoros fue diferente entre sistemas. Se registró mayor número en el SSP (10 familias), en comparación con las identificadas en el convencional (cinco familias).
En los SSP, la composición de familias estuvo asociada a una funcionalidad relativa a cambios en la estructura del suelo, determinada por familias del grupo de ingenieros del suelo (Formicidae, Termitidae, y Lumbricidae). Le siguieron las que forman parte del gremio de depredadores (Scolopendridae, Theraphosidae, Salticidae, entre otras), lo que sugiere una presencia importante de controladores biológicos. Asociada a la alta cantidad de ind/m2 del gremio depredadores, se registró baja cantidad de ind/m2, perteneciente a familias con funciones de herbívoro, siendo algunas de ellas Scarabaeidae, Aphididae, Noctuidae y Acrididae.
En el sistema convencional, la composición de las familias por funcionalidad registró mayor riqueza de depredadores, con nueve. Se encontraron Theraphosidae y Salticidae entre las más importantes por la cantidad de individuos. En cuanto al grupo de los detritívoros, estuvieron representados por cinco familias, entre las que hallan Stratiomyidae y Tenebrionidae. Con el mismo número de familias se identificó el grupo ingenieros del suelo, siendo Formicidae y Lumbricidae como las más importantes. El de los herbívoros, con cuatro, fue el menos representativo en la composición de familias (tabla 3).
La diferencia en la composición de las familias, en número como en la importancia de grupos funcionales entre sistemas, sugiere distintos niveles de activación biológica del suelo, con ventaja para el silvopastoril.
Se registró mayor cantidad de individuos de macrofauna en el SSP, con 11 593, que correspondió, aproximadamente, a cuatro veces la cantidad registrada en el convencional (3 062). La diferencia entre sistemas estuvo marcada por el número de individuos registrados por las familias Formicidae (4 150) y Termitidae (2 126), que representaron 54 % de los ind/m2 en el SSP (figuras 1 y 2).

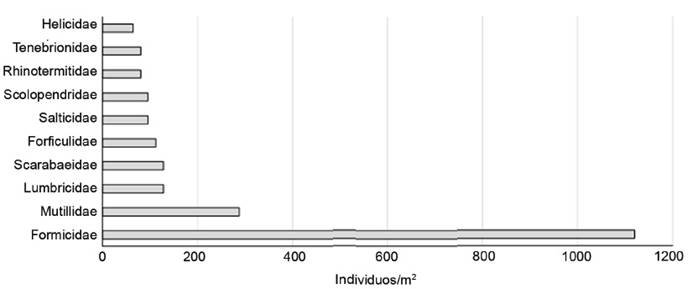
En el manejo convencional, las dos familias con mayor dominancia, Formicidae y Mutillidae (1 258 y 290 ind/m2, respectivamente), representaron 45 % del registro total de organismos, por lo que este sistema tuvo menor dominancia de familias con respecto al SSP.
Un comportamiento similar se registró en la densidad de las familias con predominancia en los sistemas evaluados. En ambos sistemas, se observó la existencia de una a tres familias con notables diferencias de ind/m2, en comparación con las familias complementarias de la composición (figuras 2 y 3). Se vio acentuada la similaridad entre sistemas, al encontrarse Formicidae y Lumbricidae entre las predominantes.

Díaz-Porres et al. (2014) manifestaron que es notoria la cantidad de ind/m2 de la macrofauna edáfica en estas familias, al ser identificadas en sistemas ganaderos diversificados, en los que la actividad biológica se incrementa con la presencia de especies arbóreas y arbustivas que incorporan materia orgánica. Cabrera-Dávila et al. (2011) informaron que estas familias realizan funciones ecológicas importantes en los sistemas ganaderos, lo que contribuye a incrementar la materia orgánica y mejorar las propiedades del suelo.
Las familias con mayor dominancia fueron Formicidae (56,9 %), Termitidae (14,5 %), Lumbricidae y Scolopendridae, que realizan funciones importantes en los ecosistemas, específicamente en lo que tiene que ver con la regulación de la dinámica del suelo y su influencia en los cultivos. Se registraron con mayor proporción en el grupo trófico ingenieros del suelo las familias Formicidae, Termitidae y Lumbricidae (tabla 3), comunes en los resultados del monitoreo de la macrofauna edáfica, con demostrada funcionalidad en procesos de mejoramiento de la estructura del suelo. Con respecto a Lumbricidae, Lavelle et al. (2003) enfatizaron en su importancia en la activación biológica del suelo, específicamente por la construcción de bioestructuras o bioagregados a cierta profundidad.
La proporción de individuos de las familias ingenieros del suelo fue diferente entre sistemas. Formicidae registró mayor porcentaje (41,0 %) en manejo convencional, en comparación con el sistema silvopastoril (35,7 %). La proporción de Lumbricidae fue mayor (7,7 %) en el sistema silvopastoril, y menor en el manejo convencional (5,8 %).
En el caso de Termitidae, las diferencias en las proporciones entre sistemas son más notables, con 13,0 % en el SSP, y 1,4 % en el manejo convencional. Los grupos funcionales herbívoros (17,5 %) y detritívoros (12,0 %) presentaron valores superiores en el sistema convencional, en comparación con el SSP. En ambos se registró igual proporción (26,0 %) del grupo trófico depredadores.
Acerca de las familias que no se registraron en el sistema convencional, se debe destacar su baja densidad (16 a 128 ind/m2), lo que sugiere potencial susceptibilidad en sus poblaciones ante cambios en las prácticas de manejo, y, por tanto, en la calidad del hábitat. Entre las 13 familias diferentes que sobresalieron en el sistema silvopastoril, se destacan las que desempeñan funciones de depredación (5), herbívoros (5), y detritívoros en menor número (3), mientras que las familias que se registraron únicamente en el sistema convencional fueron, en mayor número, herbívoros (4), y una con función de detritívoros.
El análisis de la densidad de organismos por profundidad de muestreo, como indicador de diferencias en la activación biológica del perfil del suelo, registró alta concentración de macrofauna en la capa superficial del suelo, con 5 480 ind/m2 (47,0 %) en el SSP, y 1 860 ind/m2 (60,0 %) en el convencional, y se observó una disminución a mayor profundidad. Este comportamiento ha sido referido como un patrón en diversos estudios asociados a la distribución vertical de la macrofauna (figura 4).
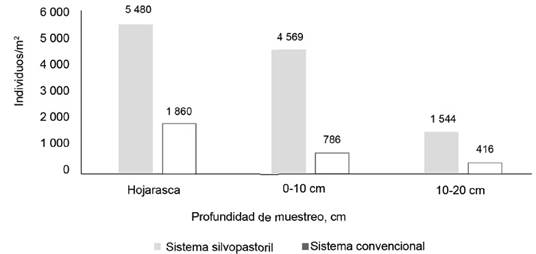
Figura 4 Cantidad de organismos (ind/m2) de la macrofauna edáfica por profundidad de muestreo y sistema ganadero.
En la figura 4, la mayor densidad de macrofauna edáfica se mostró en los primeros 10 cm del suelo. A nivel de las primeras capas, la reducción en la densidad de organismos fue marcada en el sistema convencional (con reducción superior al 50,0 %), en comparación con el SSP, donde la reducción de densidad entre la superficie y de 0 a 10 cm fue de 17,0 %.
Resultados similares a los de este estudio informaron Noguera-Talavera et al. (2017), al analizar la distribución de la macrofauna en dos sistemas de manejo del suelo. Estos autores sugirieron una mayor concentración de individuos en el sistema agroecológico, así como mayor densidad a nivel de todas las profundidades evaluadas. Ellos vincularon los resultados, en parte, a prácticas de manejo del suelo, por la adición de abonos orgánicos, que mejoran las condiciones de hábitat para la macrofauna edáfica.
Conclusiones
La mayor riqueza de familias y cantidad de individuos se obtuvo en el sistema silvopastoril, lo cual evidencia la importancia de este en las explotaciones ganaderas.
La predominancia de familias del grupo ecológico ingenieros del suelo y depredadores evidencia la existencia de procesos que transforman las características del suelo, y de mecanismos de autoregulación de poblaciones de plagas potenciales en los sistemas evaluados.
La predominancia y funcionalidad de las familias por profundidad de muestreo demuestran la hipótesis de alta concentración y mayor actividad biológica en los primeros 10 cm del perfil del suelo, así como un notable balance en las funciones de depredación y acumulación de detritos.

