










 Servicios personalizados
Servicios personalizados Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La pérdida de especies autóctonas crece de manera desmedida, y una de las principales razones es el desconocimiento de sus bondades y las características que favorecen su persistencia (Oscanoa-Lagunas, 2005). El género Desmodium Desv. pertenece a la familia Fabaceae (Leguminosas), subfamilia Faboideae, tribu Desmodieae. Desmodium incanum DC., (sin. Desmodium canum Schinz & Thell.). Desmodium incanum DC, que se conoce comúnmente como pega-pega, se distribuye a lo largo de América, desde Canadá hasta Argentina central y Uruguay (Burkart, 1987). En Argentina está presente en diez provincias (Vanni, 2008).
Se trata de una especie forrajera perenne, de ciclo primavero-estival, con período de floración en noviembre y fructificación en diciembre, y otro de floración en febrero y fructificación en marzo (Fernández et al., 1988). Es apetecible para los animales, y por sus propiedades nutricionales resulta ser un alimento de buena calidad. Es muy común en pastizales naturales. Hasta el momento no se conocen problemas de toxicidad asociados a ella (Scandaliaris et al., 2013). Estudios preliminares de la digestibilidad de sus hojas, realizados en una población de esta especie, ubicada en áreas de los alrededores de Paraná, informan 14,8 y 19,5 % de proteína bruta en los estados vegetativo y reproductivo, y una energía metabólica de 2,31 y 2,44 Mcal/kg de MS, respectivamente (Galussi, A. y Gillij, Y., com. pers., 2017; 2018).
Para determinar la densidad de siembra de esta especie, es necesario conocer el peso y las distintas categorías de semillas y plántulas halladas en el análisis de germinación. Aun cuando se cuenta con metodologías estipuladas por las normas de la Asociación Internacional para Análisis de Semillas (ISTA, 2016), no se dispone de información acerca de la calidad de las semillas de esta especie producidas en la región de Entre Ríos. Al respecto, se conoce que el contenido de humedad de las semillas, así como la temperatura, son factores determinantes en su conservación (Marcos Filho, 2005). En semillas de Medicago sativa L., por ejemplo, para un almacenamiento adecuado es preciso disminuir el contenido de humedad de 25-20 % (cosecha) a valores inferiores al 12 % (Rivas-Jacobo et al., 2007).
En la semilla, el agua se encuentra en distintas formas de asociación: agua tipo 1, 2, 3 y 4, con contenido menor de 7,5 %, entre 7,5 y 20 %, entre 20-33 % y de 33 a 41 %, respectivamente (Villela et al., 2003). Según el contenido de agua de la semilla, será su grado de deterioro a través del tiempo (Villela et al., 2003; Marcos-Filho, 2005).
En semillas de D. incanum, cosechadas en Entre Ríos, no se conoce el contenido de humedad en el momento de la cosecha. Sin embargo, este es un indicador de calidad que es importante determinar para establecer su relación con la longevidad de las semillas.
Con respecto a la germinación, aun cuando la semilla sea viable, una germinación lenta y pobre en condiciones naturales y apropiadas (disponibilidad de temperatura, luz, agua y oxígeno) se explica como producto del fenómeno fisiológico conocido como dormición de la semilla (Quinlivan, 1971). Según Bewley y Black (1986), la dormición de las semillas es un factor importante en la dinámica de las poblaciones naturales, y está relacionada con la adaptación de las plantas a la heterogeneidad ambiental. En ensayos preliminares de germinación con semillas de D. incanum, obtenidas en Entre Ríos, durante el período de 2016 a 2018 (Galussi, A., com. pers., 2016; 2018), se observaron bajos valores de plántulas normales y elevada cantidad de semillas no germinadas, categorizadas como semillas duras.
En el análisis de germinación en fabáceas y otras familias botánicas se menciona la presencia de semillas duras como manifestación de dormición física, carácter que ha sido referido por diferentes autores (Galussi et al., 2013a; Orsenigo et al., 2019; Paul et al., 2019).
El término semilla dura se aplica a la semilla que presenta una cubierta impermeable al agua, y no logra germinar porque su embrión no puede embeber (ISTA, 2016). La dormición física se relaciona con diversas características de las semillas, como la anatomía y la composición de sustancias químicas en el tegumento seminal, que ocasionan diferentes grados en la profundidad de dormición (Galussi et al., 2019). Los grados de profundidad de la dormición física originan distintos niveles de permeabilidad en las semillas, que se pueden analizar mediante modelos matemáticos. Estos modelos permiten una descripción adecuada del proceso de imbibición (Chapra y Canale, 2004). Para la ruptura de la dormición física se citan varias metodologías (ISTA, 2016; Zapata et al., 2017; Castillejo-Jiménez, 2019).
Teniendo en cuenta la escasa información acerca de los atributos de calidad de las semillas de D. incanum, producidas en la región de Entre Ríos, en Argentina, este estudio tiene como objetivo determinar la calidad y dormición física de dichas semillas.
MATERIALES Y MÉTODOS
Localización. Se cosecharon frutos (lomentos) de plantas de dos áreas naturales cercanas: Colonia Ensayo, Diamante (31°51’14.0’’S y 60º 33’02.0’’W) y Paraná, Santuario La loma (31°45’44.0”S y 60°31’57.0”W), situadas en la provincia de Entre Ríos, Argentina.
La colecta se realizó entre diciembre y febrero de 2017-2018 en una superficie de 20 m2 para cada área, cuando los artejos con semillas se encontraron maduros con una coloración marrón. La determinación del color del pericarpio y la cubierta seminal se realizó mediante la comparación con la carta de colores de Munsell (2000). Los frutos se almacenaron en el laboratorio, en recipientes plásticos, a temperatura ambiente (20 ± 1 °C) hasta el inicio de los ensayos. Del conjunto homogéneo de artejos cosechados (650 g) se extrajeron las semillas manualmente para realizar los ensayos evaluativos de calidad.
Contenido de humedad. Este análisis se realizó según el método de alta temperatura constante, establecido según las reglas del ISTA (2016), con leves modificaciones en el peso de semillas. Para esto, una vez extraídas las semillas del fruto, se colocaron en recipientes metálicos dos repeticiones de 1,5 g de semillas por ensayo y por cosecha, a 130-133 °C, durante una hora. Luego se dejaron enfriar en el desecador durante una hora. La tolerancia entre repeticiones estuvo entre la aceptada (≤ 0,2%). El contenido de humedad se calculó, como porcentaje en peso, con tres cifras decimales para cada repetición y el resultado se informó redondeado al 0,1 % más cercano.
Peso de 1 000 semillas. Se determinó a partir de la muestra homogeneizada, con ocho repeticiones de 100 semillas (ISTA, 2016).
Germinación y viabilidad. Las evaluaciones se realizaron con cuatro repeticiones de 100 semillas. Las semillas se colocaron entre papel humedecido para germinación (en rollo) y se pusieron en bolsas de polietileno para evitar la evaporación. Se llevaron a cámara a 25 °C, con un fotoperíodo de 8/16 horas, durante 21 días. Se evaluaron las plántulas según las normas del ISTA (2016). A las semillas no germinadas se les evaluó la viabilidad mediante el ensayo topográfico por tetrazolio. Para ello se realizó un corte en el tegumento seminal en el extremo distal de los cotiledones (zona del antirrafe). Después las semillas se colocaron en agua hasta completar la imbibición. Luego, se sumergieron en una solución acuosa al 0,5 % de 2, 3, 5-trifenil cloruro de tetrazolio a 20 °C durante 12 horas. Se evaluó la topografía de la tinción en los embriones y se determinó la viabilidad (ISTA, 2016).
Velocidad de hidratación. Se realizaron ensayos de permeabilidad de las semillas al agua, según Galussi et al. (2013a; 2013b). Se tomaron de la muestra tres repeticiones, de 400 semillas cada una, previamente observadas bajo lupa (× 10). Se descartaron aquellas semillas que estaban agrietadas, fisuradas o carentes de parte de la testa. Las semillas se sumergieron en agua y se consideraron completamente hidratadas cuando se observó un aumento de tamaño máximo (80 % de su peso fresco). De esta manera se determinó su tiempo de permeabilidad. Durante el período de inmersión en agua, se observaron todos los días por una semana. Posteriormente, dos veces a la semana durante un mes, y una vez a la semana hasta los 269 días de estar en agua y finalizar el ensayo. Se registró a su vez, el número de semillas permeables (embebidas) y el tiempo en días hasta observar aumento de tamaño y ruptura del tegumento seminal. Posteriormente, para evaluar germinación, se colocaron entre papel húmedo, según la metodología antes explicada.
Ensayo de longevidad. La evaluación de la longevidad (germinación a través del tiempo) se realizó mediante ensayos de germinación (PG), según la metodología antes explicada. Para ello se establecieron tres momentos: a) siembra inmediatamente de realizada la cosecha, b) a los 12 meses y c) a los 22 meses. De cada fracción se sembraron cuatro repeticiones, de 100 semillas puras, colocadas entre papel húmedo para geminación, dispuestas en una estufa a 25 °C, con un fotoperíodo de 8 /16 h luz y oscuridad.
Análisis estadístico. Se modelaron los resultados de porcentaje de semillas embebidas acumuladas (y) con el tiempo (días) como única variable independiente (x) y se seleccionó el modelo que mejor representó la liberación de latencia física. El método de estimación utilizado fue el de mínimos cuadrados, y el criterio para la selección del modelo fue el mayor coeficiente de determinación, R2. Se utilizó el programa InfoStat (Di Rienzo et al., 2019).
RESULTADOS Y DISCUSIÓN
Calidad de las semillas. La tabla 1 muestra las características medias de las semillas de D. incanum, correspondientes al peso, humedad, germinación y viabilidad (semillas duras).
Tabla 1 Calidad de semillas de D. incanum.
| Peso de 1000 semillas, g | Porcentaje, % | |||||
|---|---|---|---|---|---|---|
| Humedad | Plántulas | Semillas no germinadas | ||||
| Normales | Anormales | Muertas | Duras | |||
| Viables | No viables | |||||
| 4,12 ± 0,04 | 8,2 | 3,5 ± 1,0 | 1,0 ± 0,96 | 0,5 ± 0,5 | 89,5 ± 1,3 | 5,5± 2,0 |
Con relación al peso de las semillas, se pudo establecer que el valor registrado fue de 4,1 g, que se aproxima al peso de Medicago polymorpha L., que fue de 4,5 g (Ovalle et al., 2005), y es superior al de M. sativa (2,21-2,49 g). Según Galussi et al. (2013a), la semilla de D. incanum es de forma similar a la de estas especies, pero resulta de mayor tamaño. El peso también podría estar condicionado por el mecanismo de la dormición, específicamente si se inicia durante el período de acumulación de materia seca o deshidratación temprana (Bewley y Black, 1986).
El contenido de humedad de las semillas (8,2 %) fue ligeramente mayor que el citado para Desmodiumovalifolium (7,6 %) y sin presencia de semillas duras (Muñoz et al., 2009). No obstante, fue menor al que informan Rivas-Jacobo et al. (2007) y Galussi (2013a) en muestras de semillas de M. sativa almacenadas en la región (9-12 %).
Esto se puede explicar porque las semillas duras de M. sativa y Trifolium repens L., presentaban bajo contenido de agua (6-7 %), según afirman Galussi et al. (2013a; 2013b). Probablemente, el elevado número de semillas duras presentes en la muestra de D. incanum (95 %) explicaría el bajo contenido de agua encontrado en la muestra.
La humedad indicó la presencia de agua tipo 2 (cantidad muy cercana al tipo 1), lo que evidencia muy baja actividad metabólica en la semilla, y favorece la conservación de la viabilidad de las semillas ortodoxas (Marcos-Filho, 2005).
Si se relaciona con la regla de Harrington (1972), válida para contenidos de humedad en el rango de 4 a 14 %, que sostiene que por cada punto que se reduce el contenido de humedad de la semilla se duplica su potencial de almacenamiento, las semillas duras tendrían elevado potencial de conservación.
Meyer et al. (2007) determinaron que en semillas duras de soja la conductividad hidráulica en los tegumentos es menor que en semillas no duras. En correspondencia con ello, las semillas duras no están expuestas a un rápido intercambio con la humedad ambiente y, como consecuencia, se verían menos afectadas por el mecanismo de sorción (adsorción-desorción), manteniendo su mínima actividad respiratoria, lo que favorece que el deterioro sea menor.
El ensayo de germinación mostró bajo porcentaje de plántulas normales (3,5 %) y elevada cantidad de semillas duras (95,0 %). Sin embargo, según el ensayo topográfico por tetrazolio, 89,5 % de los embriones se observaron viables (tabla 1).
Los embriones no viables mostraron tejido dañado en la radícula y en el eje hipocótilo-radícula. La ocurrencia de plántulas anormales en el ensayo de germinación fue baja. Correspondió a plántulas con la raíz primaria atrapada en la cubierta seminal y plántulas con raíz primaria muerta. Estos resultados concuerdan con lo informado por Rojas y Herrera (1988) para D. ovalifolium, quienes señalaron que la baja germinación es un problema común en los lotes de semillas de esta especie y responde a la dormición física.
En la muestra de D. incanum analizada en este ensayo se encontró alto porcentaje de semillas duras (95 %). Es por ello que, en la muestra estudiada, el peso, la humedad y la germinación/viabilidad manifestaron, mayormente, resultados que responden a los atributos de la categoría semillas duras. La baja germinación coincide con lo expresado por Guerrero y Alfandy (2019) para Desmodium heterocarpon (L.) DC. (20 % de germinación). El elevado porcentaje de semillas duras puede expresar la adaptación de la especie al ambiente en su reproducción sexual, de acuerdo con lo expresado por Bewley y Black (1986).
Velocidad de hidratación. En cuanto al carácter de permeabilidad al agua de las semillas, el análisis matemático del porcentaje acumulativo de semillas permeables (variable respuesta) en función del tiempo de inmersión, expresado como días hasta la imbibición (variable explicativa), mostró diferentes profundidades de dormición física. La curva de la dormición física de las semillas de la muestra fue de tipo sigmoide (fig. 1), lo que mostró que la profundidad de la dormición física fue gradual. La curva respondió a un modelo polinómico de tercer grado, que se expresa como:
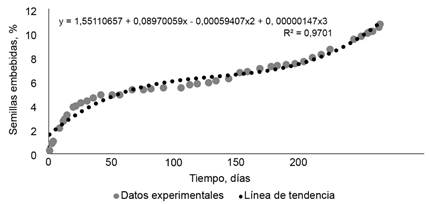
La variable y representa el porcentaje de semillas completamente embebidas. La x corresponde a los días de inmersión hasta estar completamente embebidas.
Este modelo presentó un coeficiente de ajuste de 97,1 %. En cuanto al análisis de residuos, 1,08 % de semillas embebidas (13 individuos de 1 200) en los primeros cuatro días expresaron una rápida permeabilidad al agua. Se lograron establecer tres categorías de dormición física: a) media, semillas que requirieron hasta 42 días de inmersión; b) profunda, semillas de muy lenta imbibición, con más de 42 días de inmersión y hasta 250 días; c) muy profunda, semillas que se hidrataron después de 250 días de permanecer en agua.
En la muestra, las categorías a y b evidenciaron 5 y 10 % de semillas permeables, respectivamente. Las semillas que al ponerlas a germinar rompieron la dormición física, desarrollaron plántulas normales. El resto, 90 % de las semillas duras (categoría c), permaneció con dormición física hasta finalizado el ensayo, a los 269 días.
Los resultados confirmaron la presencia de una elevada cantidad de semillas duras con diferentes categorías de dormición física. El comportamiento hallado en la muestra fue diferente a lo informado para M. sativa y el trébol blanco (Galussi et al., 2013a; 2013b). Los distintos grados de dormición física en D. incanum se pueden relacionar con los componentes químicos encontrados en las semillas duras (Galussi et al., 2019).
Ensayo de longevidad. Los valores acumulados de plántulas normales respondieron a las semillas con distinto período de almacenamiento en seco, que permanecieron en el ensayo hasta su germinación. Durante el tiempo de almacenamiento hasta el momento del ensayo de germinación (0 mes, 12 meses y 22 meses), las semillas presentaron una ruptura natural de la dormición, que se manifestó en el primer conteo de plántulas.
Las semillas cosechadas y sembradas (0 mes) expresaron 0,25 % a los 12 meses 0,5 %. En tanto, a los 22 meses 8,5 % de semillas permeables dieron plántulas normales. Las semillas de 0 mes, cuando alcanzaron 269 días de permanencia en el ensayo de germinación, originaron 11 % de plántulas normales (fig. 1). Las de 12 y 22 meses de edad, a los 300 días de sembradas, lograron 65 y 76 % de germinación, respectivamente (figura 2).
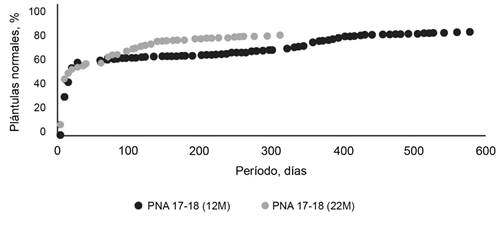
En el ensayo de longevidad se observó que el ambiente húmedo del sustrato de germinación favoreció más rápidamente la ruptura natural de la dormición en las semillas de mayor edad que el ambiente seco de almacenamiento (temperatura ambiente en laboratorio). Las semillas que rompieron la dormición física originaron plántulas normales, por lo que se puede afirmar que la dormición física protege la viabilidad del embrión mientras dura dicha característica, lo que coincide con el comportamiento de otras leguminosas forrajeras (Zimmermann et al., 1998; Galussi et al., 2013a; 2013b).
Para asegurar una implantación exitosa del cultivo, con semillas producidas en la provincia Entre Ríos, se requiere de la ruptura de la dormición física, que debe estar acorde con las categorías de dormición que se presenten. No obstante, los diversos tratamientos para la ruptura de la dormición física (ISTA, 2016; Zapata et al., 2017; Castillejo-Jiménez, 2019) tuvieron en común una característica: al superar la dormición, eliminan parcial o totalmente el mecanismo o los mecanismos de protección de las semillas contra la adversidad del ambiente. Por ello es conveniente que las semillas se utilicen inmediatamente después de recibir el tratamiento (Marcos-Filho, 2005).
A partir de los resultados, es recomendable continuar con investigaciones en semillas de una segunda cosecha, y realizar estudios anatómicos del tegumento seminal y los componentes químicos, asociados a las categorías de dormición física en las semillas de D. incanum.
Conclusión
Las semillas de la especie producida en la región presentaron elevada cantidad de embriones viables, aunque la mayoría presentó dormición física profunda y predominio de la fracción con gran profundidad de dormición.
El contenido de humedad en el momento de la cosecha permite el almacenamiento seguro de las semillas, sin necesidad de aireado o secado previo. Las semillas duras, evaluadas en el tiempo, mostraron lenta ruptura de la dormición física, por lo que originaron plántulas normales, aun después de 22 meses de almacenamiento.
El modelo propuesto dejó ver el comportamiento de los individuos de la muestra, que mostraron diferente profundidad de dureza física en el tegumento seminal. El almacenamiento en ambiente húmedo favoreció la ruptura de la dormición, principalmente en la semilla de mayor edad

