










 Servicios personalizados
Servicios personalizados Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los pastos del género Urochloa (syn. Brachiaria) se cultivan ampliamente, debido a su alta palatabilidad y capacidad para producir biomasa con adecuado valor nutritivo en condiciones edáficas y climáticas adversas para otras especies (Baptistella et al., 2021). En Cuba, aunque se encuentran extendidos a lo largo del país, tienen una presencia importante en las áreas ganaderas ubicadas en la región geográfica Sabana de Manacas. Esta zona abarca parcialmente las provincias de Matanzas, Villa Clara y Cienfuegos. Sus suelos son de textura ligera, baja fertilidad natural y elevada acidez, condiciones que limitan la productividad de los pastos y su período de vida útil, dando lugar a la aparición de plantas invasoras de escaso valor nutritivo para la alimentación animal (Pereira et al., 2018).
El uso de altas dosis de fertilizantes resulta imprescindible para incrementar la oferta de forraje por unidad de superficie y tiempo y mejorar el valor nutritivo de la biomasa (Silva-Parra et al., 2021; Cabral et al., 2023). No obstante, los altos precios de los fertilizantes y la necesidad de fomentar una ganadería amigable con el medio ambiente conducen a la búsqueda de alternativas biológicas. Estas se basan en el manejo de las interacciones entre los microorganismos del suelo y las plantas, con el propósito de incrementar la productividad de los cultivos agrícolas y reducir la dependencia de insumos externos.
El manejo efectivo de las asociaciones micorrízicas constituye una vía para las intenciones referidas, debido a la capacidad de los hongos formadores de micorrizas arbusculares (HMA) para incrementar el volumen de suelo que exploran las raíces y, consecuentemente, mejorar el acceso de las plantas a los nutrientes del suelo y al agua, amén de otros beneficios como la inducción de mecanismos de respuesta ante el ataque de agentes patógenos (Chandrasekaran, 2020).
El género Urochloa se ha demostrado que la inoculación con cepas eficientes de HMA mejora los rendimientos y el valor nutritivo de la biomasa, contribuye a la recuperación de las plantas después del corte o pastoreo y reduce las dosis de fertilizantes minerales u orgánicos que requieren normalmente estos cultivos (Karti et al., 2021; Chang et al., 2023). Sin embargo, factores relacionados con las características del suelo y la comunidad de HMA residente, la capacidad de las cepas para adaptarse al nicho ecológico donde se introducen, las especies o cultivares y su manejo, pueden influir en la efectividad de la inoculación (Tshibangu-Kazadi et al., 2020; Silva-Parra et al., 2021). De ahí la importancia de evaluar el efecto de la biofertilización con estos microorganismos en los diferentes agroecosistemas.
A partir de estas premisas, este estudio tiene como objetivo evaluar la respuesta a la inoculación con cepas de hongos micorrízicos arbusculares de cuatro especies de pasto del género Urochloa, cultivadas en un suelo ácido de baja fertilidad de la región Sabana de Manacas, Cuba.
Materiales y Métodos
Localización. El experimento se realizó en la Estación de Pastos y Forrajes de Cascajal, ubicada a los 22° 39′ 44” de latitud norte y 80° 29’ 36” de longitud oeste, en la región geográfica Sabana de Manacas, provincia de Villa Clara, Cuba.
Características de suelo. El estudio se ejecutó en un suelo gley nodular ferruginoso (Hernández-Jiménez et al., 2015), cuyas características químicas principales se muestran en la tabla 1.
Tabla 1 Características químicas del suelo (profundidad 0-20 cm).
| pH H2O | MO, % | P, mg 100 g-1 | Ca2+ | Mg2+ | Na+ | K+ | CIB | H+ + Al3+ | Al3+ | V, % |
|---|---|---|---|---|---|---|---|---|---|---|
| (cmol kg-1) | ||||||||||
| 4,8 | 2,52 | 5,9 | 3,32 | 1,12 | 0,05 | 0,1 | 4,59 | 4,33 | 0,06 | 51 |
| (0,2) | (0,17) | (0,3) | (0,3) | (0,1) | (0,01) | (0,02) | (0,31) | (0,33) | (0,01) | |
MO: materia orgánica, CIB: capacidad de intercambio de bases, H+ + Al3+: acidez intercambiable, V: saturación por bases
Valores entre paréntesis indican intervalos de confianza (α = 0,05)
El suelo poseía elevada acidez, caracterizada por un pH fuertemente ácido, altos valores de acidez intercambiable (H+ + Al3+) y muy bajo porcentaje de saturación por bases (V), además de bajo contenido de materia orgánica y muy bajos niveles de fósforo asimilable y cationes intercambiables (Paneque y Calaña, 2001). Para la caracterización química del suelo en el área experimental se tomaron cinco muestras por el método del zigzag, a profundidad de 0-20 cm. Las muestras se analizaron según el Manual de Técnicas Analíticas para el análisis de suelo, foliar, abonos orgánicos y fertilizantes químicos del Instituto Nacional de Ciencias Agrícolas (INCA) (Paneque et al., 2010).
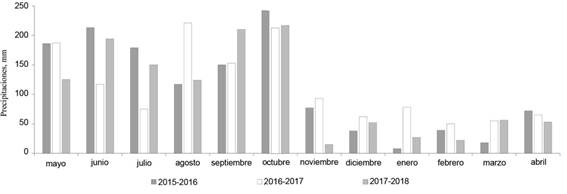
El comportamiento de las precipitaciones durante el período en que se condujo el experimento se muestra en la figura 1.
Diseño experimental y tratamientos. En un diseño de bloques al azar, con arreglo factorial 4 x 4 y cuatro réplicas, se evaluaron 12 tratamientos, resultantes de la combinación de cuatro especies de pasto del género Urochloa (U. brizantha cv. Marandú, U. decumbens cv. CIAT 606, U. híbrido cv. 36087 Mulato II y U. híbrido cv. BR02/1752 Yacaré) y la inoculación con tres cepas de HMA Funneliformis mosseae, cepa INCAM-2 (Nicol. & Gerd.) Walker & Schüßler, Glomus cubense cepa INCAM-4, (Y. Rodr. & Dalpé), Rhizoglomus irregulare cepa INCAM-11 (N. C. Schenck & G. S. Sm. Sieverd., G. A. Silva & Oehl), más un testigo sin inocular. Las parcelas constituyeron la unidad experimental, y tenían una superficie total de 21 m2 y un área de cálculo de 14 m2.
Procedimineto experimental. El suelo se preparó mediante labores de roturación (arado), grada, cruce (arado) y grada, a intervalos aproximados de 25 días entre cada una. La siembra se realizó en surcos separados a 50 cm, a chorrillo, con dosis de 1 kg de semilla pura germinable ha-1 y a profundidad de 1,5 cm. El experimento se condujo en condiciones de secano y los pastos se fertilizaron con 35 kg ha-1 de N a los 30 días después de la siembra, y luego de cada corte efectuado durante el período lluvioso de cada año con aplicaciones de (30 y 120 kg de P2O5 y K2O ha-1 año-1, respectivamente) al inicio de cada época de lluvia. Se utilizaron como portadores, urea, superfosfato triple y cloruro de potasio. El experimento tuvo una duración de tres años.
La inoculación se realizó en el momento de la siembra por el método del recubrimiento de las semillas, para lo que se sumergieron en una pasta fluida, elaborada mediante la mezcla de una cantidad de inóculo sólido, equivalente al 10 % de su peso y agua, en proporción 1:0,6 m/v. Una vez recubiertas las semillas y solidificado el inóculo, se procedió a la siembra. Los inóculos se produjeron en el departamento de biofertilizantes y nutrición de las plantas del INCA. Tenían una concentración de 35 esporas de cada especie de HMA a evaluar por gramo de inoculante, así como abundantes fragmentos de raicillas de la planta hospedera que se utilizó para su multiplicación.
El primer corte se realizó a los 120 días después de la siembra, y posteriormente a intervalos aproximados de 60 y 90 días durante el período lluvioso y poco lluvioso, respectivamente. En cada corte se pesó la masa fresca de la parte aérea de las plantas que ocupaban el área de cálculo de las parcelas. Se tomaron muestras de 200 g, que se llevaron a una estufa de circulación de aire a 70 ºC durante 72 horas, para determinar el porcentaje de masa seca (MS), estimar el rendimiento de MS y las concentraciones de N, P y K en la biomasa (Paneque et al., 2011).
Mediciones. Para la evaluación de las variables micorrízicas en cortes alternos se tomaron cinco muestras de raíces por parcela, a profundidad de 0-20 cm con la utilización de un cilindro metálico de 5 cm de diámetro y 20 cm de largo. Las muestras se unieron para formar una muestra compuesta de la que se extrajo 1 g de raicillas para su tinción y clarificación (Rodríguez-Yon et al., 2015). Se determinó la frecuencia de colonización micorrízica mediante el método de los interceptos (Giovanetti y Mosse, 1980), la densidad visual o intensidad de la colonización (Trouvelot et al., 1986) y el número de esporas en la rizosfera a partir del tamizado y decantado por vía húmeda de dichas estructuras y su observación en el microscopio (Herrera et al., 1995).
Se determinó la participación de los inoculantes en la nutrición de los pastos (Rivera et al., 2003) mediante la siguiente fórmula:
Participación, % = Concentraciones de N, P y K, g kg-1 de MS en la biomasa aérea del tratamiento inoculado-concentraciones N, P y K, g kg-1 de MS en la biomasa aérea del tratamiento no inoculado) / concentraciones de N, P y K, g kg-1 MS en la biomasa aérea del tratamiento inoculado x 100.
Análisis estadístico. Para el procesamiento estadístico, después de comprobadas la normalidad y la homogeneidad de varianza de los datos, se aplicó el análisis de varianza, de acuerdo con el diseño experimental y la prueba de rangos múltiples de Duncan (p < 0,05). Para la caracterización química del suelo, así como para evaluar las estructuras micorrízicas y la participación de los inoculantes en la nutrición de los pastos, se utilizó el intervalo de confianza a α = 0,05 (Payton et al., 2000). En todos los casos se usó el programa estadístico SPSS 25 (IBM SPSS Statistics, 2017).
Resultados y Discusión
No hubo interacción entre la inoculación entre las cepas de HMA y las especies de pasto en las estructuras micorrízicas, pero los tratamientos del factor inoculación mostraron diferencias entre sí. Como muestra la figura 2, todas las cepas incrementaron la frecuencia e intensidad de la colonización y el número de esporas en la rizosfera con respecto al testigo sin inocular, que reflejó el nivel de ocupación radical de los HMA residentes. Sin embargo, los mayores valores de estas variables se alcanzaron con F. mosseae, cuyo efecto se mantuvo hasta el período poco lluvioso del segundo año.
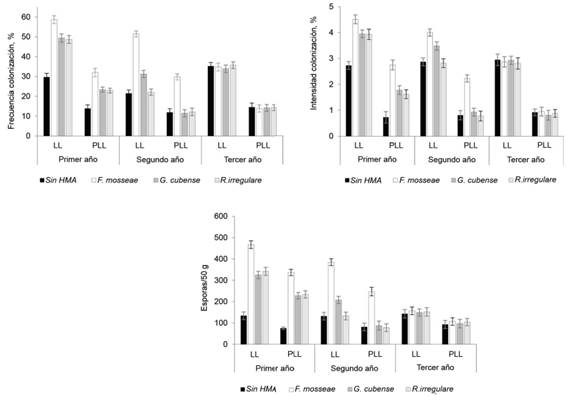
Figura 2 Efecto de inoculación con cepas de HMA en la frecuencia e intensidad de la colonización y el número de esporas en la rizosfera.
Los inóculos con G. cubense y R. irregulare no solo produjeron menores frecuencia e intensidad de colonización y número de esporas que F. mosseae, sino que sus efectos tuvieron menor permanencia, pues sólo se observaron hasta el período lluvioso y poco lluvioso del segundo y el primer año, respectivamente. En el tercer año no se observó efecto de la inoculación en las variables micorrízicas.
No se encontraron diferencias significativas entre las especies de pasto para las variables fúngicas, las que también mostraron los mayores valores en el período lluvioso de cada año (figura 3).
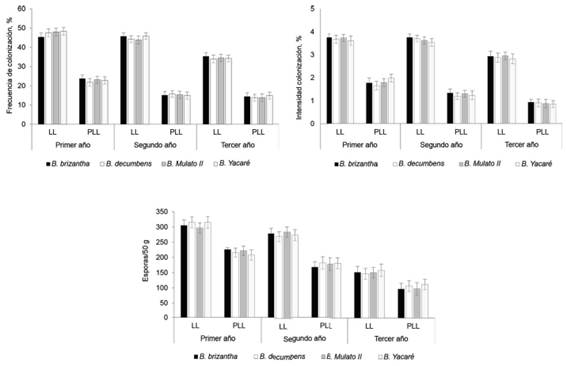
Figura 3 Efecto de las especies de pasto en la frecuencia e intensidad de la colonización y el numero de esporas de HMA en la rizosfera.
Estos resultados corroboran los encontrados por González-Cañizares (2014), quien al evaluar el efecto de la inoculación con cepas de HMA en pastos del género Brachiaria cultivados en dos tipos de suelo, concluyó que para cada condición edáfica hubo, al menos, una cepa altamente efectiva para incrementar los valores de las variables micorrízicas en todas las especies de pasto. Esto indica, de acuerdo con el autor citado, alta compatibilidad entre la cepa eficiente de HMA y el tipo de suelo y baja especificidad entre la cepa eficiente de HMA y las especies de pasto. Estas regularidades se han encontrado también en otros cultivos con diferentes patrones de crecimiento y requerimientos nutricionales en un grupo importante de suelos (Rivera et al., 2020; Simó et al., 2020).
Existen evidencias consistentes de que las propiedades del suelo, específicamente el pH, influyen marcadamente en la efectividad de la inoculación con cepas de HMA (Tshibangu-Kazadi et al., 2020). De hecho, la inoculación con F. mosseae ha sido más efectiva en ambientes edáficos caracterizados por su elevada acidez y baja fertilidad (Rivera-Espinosa et al., 2015), condiciones similares a las del suelo donde se realizó este estudio.
Los mayores valores de frecuencia e intensidad de la colonización y el número de esporas en la rizosfera, registrados en los períodos lluviosos con respecto a los secos, se pueden relacionar con la variación estacional de la producción de biomasa de los pastos. Su mayor crecimiento en la época de más precipitaciones, temperatura y luminosidad, propicia más demanda de nutrientes y, de hecho, mayor formación de estructuras micorrízicas para acceder a los recursos del suelo (Cera et al., 2021).
No hubo interacción entre la inoculación con cepas de HMA y las especies de pasto para las concentraciones (promedios anuales) de macronutrientes en la biomasa, pero el factor inoculación sí mostró diferencias significativas entre tratamientos (tabla 2). Al igual que en las variables fúngicas, todas las cepas incrementaron las concentraciones de N, P y K con relación al testigo sin inocular. Sin embargo, los mayores valores se lograron con F. mosseae, cuyo efecto, a diferencia de los inoculantes formulados con G. cubense y R. irregulare, se prolongó hasta el segundo año. Las concentraciones de estos nutrientes no mostraron diferencias entre las especies de pasto.
Tabla 2 Efecto de la inoculación de las cepas de HMA y las especies de pasto en las concentraciones (promedios anuales) de N, P y K en la biomasa aérea, g kg-1 .
| Tratamiento | Primer año | Segundo año | Tercer año | ||||||
|---|---|---|---|---|---|---|---|---|---|
| N | P | K | N | P | K | N | P | K | |
| Efecto inoculación | |||||||||
| Sin HMA | 12,2c | 1,6c | 11,1b | 11,9b | 1,7b | 11,9b | 11,8 | 1,7 | 11,7 |
| 15,5a | 2,2a | 14,5a | 14,3a | 2,2a | 12,5a | 12,3 | 1,5 | 12,3 | |
| 13,3b | 1,9b | 13,2b | 12,1b | 1,8b | 10,1b | 12,1 | 1,6 | 11,9 | |
| 13,8b | 1,8b | 13,5b | 12,0b | 1,7b | 10,5b | 12,2 | 1,5 | 12,5 | |
| EE ± | 0,311 | 0,102 | 0,421 | 0,315 | 0,103 | 0,415 | 0,500 | 0,101 | 0,432 |
| Valor- P | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,322 | 0,489 | 0,377 |
| Efecto especies de pasto | |||||||||
| 13,7 | 1,9 | 12,7 | 12,6 | 1,9 | 11,6 | 12,1 | 1,6 | 12,5 | |
| 12,8 | 2,1 | 12,9 | 12,4 | 2,0 | 10,9 | 11,8 | 1,5 | 12,3 | |
|
|
13,9 | 1,8 | 13,0 | 12,7 | 1,7 | 11,5 | 12,3 | 1,7 | 11,7 |
|
|
13,7 | 1,7 | 12,8 | 12,7 | 1,8 | 10,7 | 11,8 | 1,5 | 11,5 |
| EE ± | 0,411 | 0,104 | 0,371 | 0,387 | 0,110 | 0,502 | 0,425 | 0,112 | 0,507 |
| Valor - P | 0,436 | 0,252 | 0,327 | 0,391 | 0,222 | 0,411 | 0,415 | 0,332 | 0,451 |
Letras no comunes en la misma columna difieren significativamente a p < 0,05, según dócima de Duncan
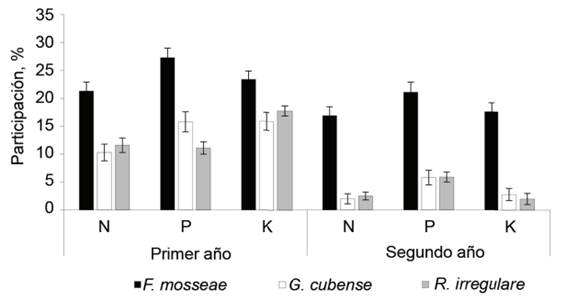
La participación de los inoculantes micorrízicos en la nutrición de los pastos se evaluó mediante el incremento porcentual de las concentraciones de N, P y K en la biomasa aérea con respecto al tratamiento no inoculado (figura 3). Solo se muestran los resultados de los dos primeros años, ya que como deja ver la tabla 2, en el tercero no hubo efecto de la inoculación en las concentraciones de estos nutrientes en la biomasa.
Al igual que en las concentraciones de nutrientes en la biomasa aérea, la mayor participación correspondió al inoculante formulado con F. mosseae, cuyo efecto se mantuvo hasta el segundo año, aunque con valores inferiores al primero, en los tres elementos. G. cubense y R. irregulare, además de tener menor participación que F. mosseae durante el primer año, tuvieron en el segundo efecto muy bajo, pues su contribución al incremento de los nutrientes en la biomasa apenas superó 5 %, solo en el fósforo.
En cultivos forrajeros inoculados con HMA se ha podido constatar que mayores niveles de colonización micorrízica se asocian, generalmente, con aumento en la absorción de nutrientes (Tran et al., 2019; Cavagnaro et al., 2021). Esto pudiera explicar las mayores concentraciones de N, P y K en la biomasa aérea de los pastos inoculados con F. mosseae, así como su mayor participación en la nutrición de los tres elementos durante los dos años en que se encontró respuesta a la inoculación. Como muestra la figura 2, durante ese período, los niveles de ocupación radical alcanzados con el inoculante que contenía esta cepa, fueron significativamente mayores que con el resto.
En la figura 4 se muestra que, durante el primero y el segundo año, los inoculantes micorrízicos tuvieron mayor contribución a la nutrición fosfórica que a la nitrogenada y potásica. No obstante, de acuerdo con los valores alcanzados, fue más notable con F. mosseae. Este comportamiento pudo estar relacionado con la función de las micorrizas en el incremento de la absorción de fosfatos del suelo, incluso de formas poco asimilables (Teutscherova et al., 2019, Chandrasekaran, 2020) y con el aumento del coeficiente de aprovechamiento del fertilizante fosfatado que se aplicó de fondo, sobre todo en el suelo donde se condujo este experimento. En ellos, los contenidos de fósforo asimilable fueron muy bajos de por sí y, al parecer, no fueron suficientes para satisfacer las necesidades de este nutriente en los pastos inoculados.
No hubo interacción entre la inoculación con las cepas de HMA y las especies de pasto para el rendimiento de masa seca. No obstante, el factor inoculación y las especies mostraron diferencias entre sí (tabla 3).
Tabla 3 Efecto de la inoculación con cepas de HMA y las especies de pasto en el rendimiento de MS durante los tres años en que se condujo el experimento (t ha-1).
| Tratamiento | Primer año | Segundo año | Tercer año | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MS LL | MS PLL | MST | MS LL | MS PLL | MST | MS LL | MS PLL | MST | |
| Efecto inoculación | |||||||||
| Sin HMA | 8,3c | 3,2c | 11,5c | 9,3c | 3,2b | 12,5c | 8,6 | 3,5 | 12,1 |
| 10,5a | 3,9a | 14,4a | 11,0a | 4,1a | 15,1a | 8,5 | 3,8 | 12,3 | |
| 9,6b | 3,3b | 12,9b | 10,0b | 3,2b | 13,2b | 8,7 | 3,7 | 12,4 | |
| 9,4b | 3,4b | 12,8b | 9,6c | 3,3b | 12,8 | 8,7 | 3,6 | 12,2 | |
| EE ± | 0,210 | 0,102 | 0,231 | 0,223 | 0,112 | 0,231 | 0,180 | 0,109 | 0,198 |
| Valor - P | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,113 | 0,143 | 0,115 |
| Efecto especies de pasto | |||||||||
| 9,1b | 3,4b | 12,4b | 9,9b | 3,1b | 13,0b | 8,4b | 3,4b | 11,8b | |
| 8,9b | 3,2b | 12,1b | 9,9b | 3,0b | 13,0b | 8,5b | 3,4b | 11,9b | |
|
|
9,1b | 3,3b | 12,4b | 9,9b | 3,2b | 13,1b | 8,3b | 3,3b | 11,6b |
|
|
10,6a | 4,1a | 14,7a | 11,0 a | 4,5a | 15,5a | 9,5a | 4,5a | 14,0a |
| EE ± | 0,231 | 0,122 | 0,267 | 0,224 | 0,108 | 0,251 | 0,207 | 0,111 | 0,243 |
| Valor - P | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 |
Letras distintas en la misma columna difieren significativamente según Duncan (p < 0,05).
MS LL: rendimiento de masa seca durante el período lluvioso, MS PLL: rendimiento de masa seca durante el período poco lluvioso, MST: rendimiento de masa seca total (período lluvioso + poco lluvioso).
En el primer año, en el período lluvioso como en el poco lluvioso, la inoculación con las cepas de HMA incrementó el rendimiento con respecto al tratamiento no inoculado. No obstante, los mayores valores se alcanzaron con F. mosseae. En el segundo, con el inoculante formulado con F. mosseae también se obtuvo el mayor rendimiento durante ambas épocas. G. cubense volvió a incrementar el rendimiento con respecto al tratamiento sin inocular, pero su efecto permaneció sólo hasta el período lluvioso. La influencia del inóculo con R. irregulare en esta variable no fue más allá del primer año. En el tercero, no se observó efecto de la inoculación en el rendimiento. Entre las especies de pasto, los mayores rendimientos durante los tres años se alcanzaron con B. Yacaré.
Al analizar de forma integral los resultados obtenidos en este experimento, se concluyó que el inoculante micorrízico formulado con F. mosseae fue más efectivo para mejorar el rendimiento de los pastos B. brizantha, B. decumbens, B. Mulato II y B. Yacaré, cultivados en un suelo gley nodular ferruginoso de baja fertilidad y elevada acidez. Esto ratificó, en las condiciones en que se condujo el ensayo, la baja especificidad entre las cepas de HMA inoculadas y las especies de pasto, tal como se observó antes al evaluar las estructuras micorrízicas. Este efecto permaneció hasta el período poco lluvioso del segundo año.
Aunque generalmente no se manifiesta una especificidad estricta hongo-planta hospedera, no todas las especies de HMA colonizan con la misma intensidad y eficiencia las distintas especies vegetales, por lo que queda demostrada la existencia de distintos grados de compatibilidad en la simbiosis, como resultado de las influencias del ambiente en la expresión genotípica de ambos simbiontes (Morales-Londoño et al., 2019; Rosales-Jenqui et al., 2021).
Trabajos realizados en Cuba, relacionados con la inoculación con HMA en diferentes cultivos agrícolas y tipos de suelos, han demostrado gran relación entre factores edáficos, fundamentalmente el pH, y la efectividad de la cepa inoculada en el aumento de los rendimientos. Se destacan los inoculantes formulados con F. mosseae como los más eficientes para las plantas cultivadas en suelos de baja fertilidad (Martín-Alonso et al., 2017).
El efecto del inoculante micorrízico que contenía la cepa F. mosseae en el incremento del rendimiento de los pastos se relacionó con la mejora del estado nutricional de estos últimos, lo que se hizo evidente por las mayores concentraciones de N, P, K en la biomasa aérea alcanzada con la inoculación, como por su mayor participación en la nutrición de los pastos en relación con G. cubense y R. irregulare. Este hecho se relaciona con el aumento del aprovechamiento de los nutrientes del suelo y de los fertilizantes aplicados a los pastos a partir de la formación de suficientes cantidades de estructuras micorrízicas que facilitaron el acceso de las plantas a estos recursos (Huang et al., 2020, Bouskout et al., 2022).
No menos interesante resultó el mejor comportamiento del pasto Yacaré con respecto a las demás especies evaluadas. Aunque en algunas regiones de Cuba este cultivar ha mostrado mayores rendimientos y niveles de persistencia que otras especies de Urochloa (Pentón-Fernández et al., 2018), el hecho de que en suelos ácidos y de baja fertilidad haya alcanzado mayor productividad, indica que además de la inoculación con HMA, la inclusión del pasto Yacaré en la estructura varietal de las especies forrajeras también puede ser una opción para incrementar la producción de biomasa en estas condiciones edáficas.
Conclusiones
La biofertilización con inoculante micorrízico formulado con F. mosseae constituye una vía efectiva para mejorar el estado nutricional y los rendimientos de pastos del género Urochlola, cultivados en suelos ácidos y de baja fertilidad, y que el efecto del inoculante permanece durante los dos primeros años posteriores a su aplicación.

