










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Ortopedia y Traumatología
versión On-line ISSN 1561-3100
Rev Cubana Ortop Traumatol v.22 n.1 Ciudad de la Habana ene.-jun. 2008
TRABAJOS DE REVISIÓN
Factores que influyen en el crecimiento endocondral: experimentos y modelos
Factors influencing on the endochondral growth: experiments and models. A review
Facteurs influençant la croissance de l'os endochondral : expériences et modèles. Une révision
Diego Alexander Garzón AlvaradoI; Máximo Alejandro Roa GarzónI; Angélica María Ramírez MartínezII
I Profesor Asociado. Universidad Nacional de Colombia. Ciudad Universitaria. Bogotá. Colombia.
II Investigadora. Universidad de Zaragoza. España.
RESUMEN
Se presentan los principales factores que influyen durante el desarrollo endocondral a nivel pre y postnatal. Los factores más importantes del desarrollo de los huesos largos se pueden clasificar en moleculares y mecánicos, los cuales a su vez se clasifican como intrínsecos y extrínsecos. En este artículo se presentan los principales avances en el conocimiento del desarrollo endocondral para lo cual se hace una revisión del estado del arte sobre los modelos experimentales, animales y algunos computacionales del conjunto de todos los factores que influyen en el crecimiento de un hueso largo. La última parte del artículo hace referencia a la aparición y desarrollo de los centros secundarios de osificación y como la mecánica y los factores moleculares, se encuentran íntimamente regulados para controlar la total formación de un hueso.
Palabras clave: Desarrollo endocondral, factores de crecimiento, biomecánica.
ABSTRACT
The main factors influencing on the endochondral development at the pre- and postnatal level are presented. The most important factors of the development of the long bones may be classified into molecular and mechanical, which at the same time, may be defined as intrinsic and extrinsic. In this paper, the fundamental advances attained in the knowledge of the endochondral development are presented, for which a review of the state of the art regarding the experimental, animal and some computer models of the set of all the factors influencing on the growth of a long bone is made. The last part of the article refers to the appearance and development of the secondary centres of ossification and to how mechanics and the molecular factors are closely regulated to control total bone formation.
Key words: Endochondral development, growth factors, biomechanics.
RÉSUMÉ
Les principaux facteurs influençant le développement de l'os endochondral aux étapes pré et postnatale sont présentés. Les facteurs les plus importants impliqués dans la formation et la croissance des os longs incluent les facteurs moléculaires et mécaniques, lesquels peuvent être classés en même temps d'intrinsèques et extrinsèques. Dans cet article, une révision de l'état de l'art des modèles expérimentaux animaux, et quelques-uns par ordinateur, de tous les facteurs influant sur la croissance d'un os long est faite afin de montrer les principaux progrès en matière de connaissances sur le développement de l'os endochondral. La dernière partie de l'article fait référence à l'apparition et au développement des centres d'ossification secondaires, ainsi qu'à l'association intime entre les facteurs biomécaniques et moléculaires dans la formation des os.
Mots clés: développement de l'os endochondral ; facteurs de croissance ; biomécanique.
INTRODUCCIÓN
El conocimiento sobre el proceso de crecimiento del hueso ha tenido un avance espectacular en los últimos 50 años,1 y se han desarrollado nuevas teorías sobre la biología molecular y celular a nivel embrionario.2 La observación ha determinado que, en conjunto con los factores moleculares y celulares, los factores epidemiológicos, nutricionales y la actividad física están incluidos en el proceso del desarrollo de los huesos.
Durante el desarrollo endocondral la placa de crecimiento es el principal motor en la elongación de los huesos.1,2 Esta se compone de varias capas que van emergiendo constantemente para producir el crecimiento endocondral de forma longitudinal y es precisamente allí donde se centra el desarrollo de esta revisión bibliográfica. La unidad funcional de la placa de crecimiento es una columna de condrocitos cuyas células se encuentran en constante proliferación, hipertrofia y están programadas para morir (apoptosis) con el correspondiente cambio en el metabolismo celular y en la matriz pericelular. Se cree que esta fisis (o placa de crecimiento) se regula principalmente por: (fig. 1).
- Factores de crecimiento intrínsecos
- Hormonas extrínsecas
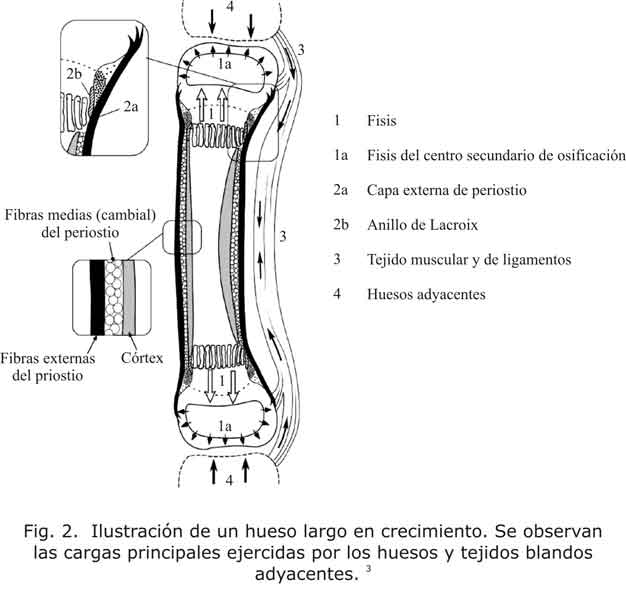
- Fuerzas mecánicas extrínsecas e intrínsecas (fig. 2).
En las siguientes secciones se detalla el funcionamiento de la placa epifisiaria y se hace especial énfasis en los nuevos hallazgos sobre el desarrollo endocondral y como los factores extrínsecos e intrínsecos, de carácter mecánico y molecular, influyen en el crecimiento.
BASES FISIOLÓGICAS
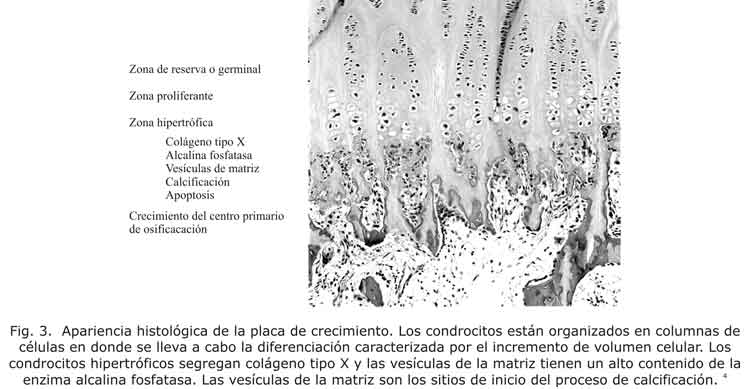
Desde el punto de vista morfológico, la placa de crecimiento se divide en células germinales, células columnares, hipertróficas y células de la zona metafisiaria. Con respecto a las funciones que desarrolla cada tipo de célula se les puede clasificar como: 1) zona germinal; 2) zona columnar con dos áreas bien definidas, la zona proliferante superior y la zona de maduración inferior; 3) zona de células hipertróficas que se divide a su vez en las 4/5 partes, superior de matriz no mineralizada y la zona restante (1/5) que es mineralizada; y 4) la parte externa de la metáfisis que alcanza la placa de crecimiento1-4 (fig. 3).
La zona mineralizada de la capa de células hipertróficas y la parte externa metafisiaria conforman la zona de calcificación provisional.3 Microscópicamente, las columnas celulares mantienen la proliferación en la parte superior y depositan matriz en la parte inferior.1,3,4 Aquellas regiones de la fisis cercana a la epífisis mantienen la proliferación de las células del cartílago, mientras que aquellas que están cerca a la metáfisis están destinadas a la mineralización de la matriz pericelular, la invasión vascular y la formación de hueso.
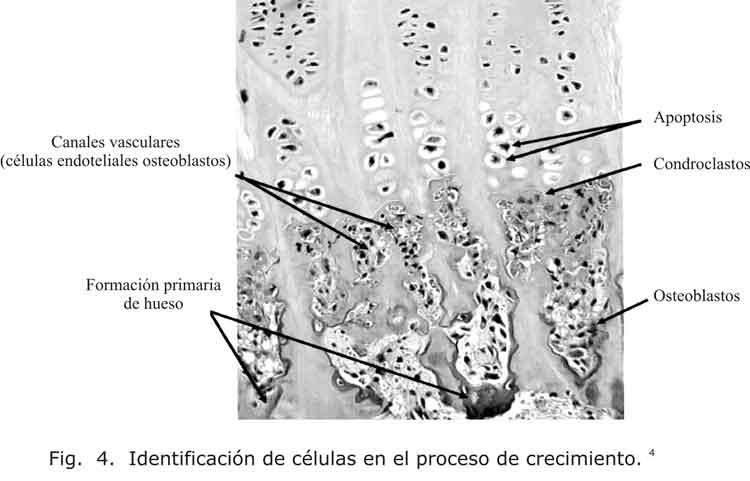
Las células hipertróficas que se encuentran cerca de las pericondriales atraen a los osteoblastos, los cuales secretan una matriz característica más dura y resistente.3 Una vez que se llega a la apoptosis por parte de los condrocitos hipertróficos, se ha construido un andamio estructural para la invasión vascular con la llegada de los osteoblastos, con lo cual se forma una matríz ósea inicial denominada esponjosa primaria (fig. 4).
FACTORES QUE INFLUYEN EN EL DESARROLLO ENDOCONDRAL
Control molecular del desarrollo endocondral
Como se ha señalado, el desarrollo endocondral viene dirigido por la placa de crecimiento lo que involucra un delicado control de proliferación, maduración y apotosis. El control exacto de la actividad celular se lleva a cabo por la síntesis de proteínas que activan procesos determinados para la acción orquestada del crecimiento endocondral. La actividad de síntesis hormonal puede ser alterado por factores genéticos 5 y por una alimentación inadecuada en los primeros meses de vida.1 La placa de crecimiento es regulada por factores moleculares y estímulos biofísicos intrínsecos y extrínsecos (fig.1). En esta sección se determinan los factores moleculares más importantes que permiten un desarrollo adecuado de los huesos largos.
Factores intrínsecos (autocrinos) moleculares del crecimiento endocondral
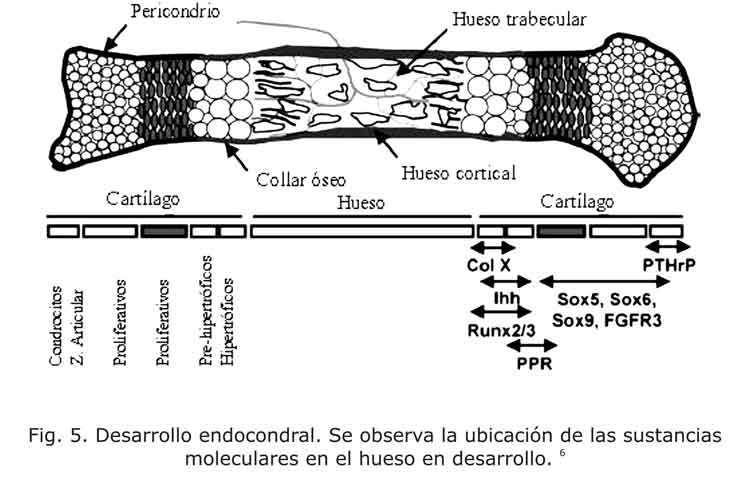
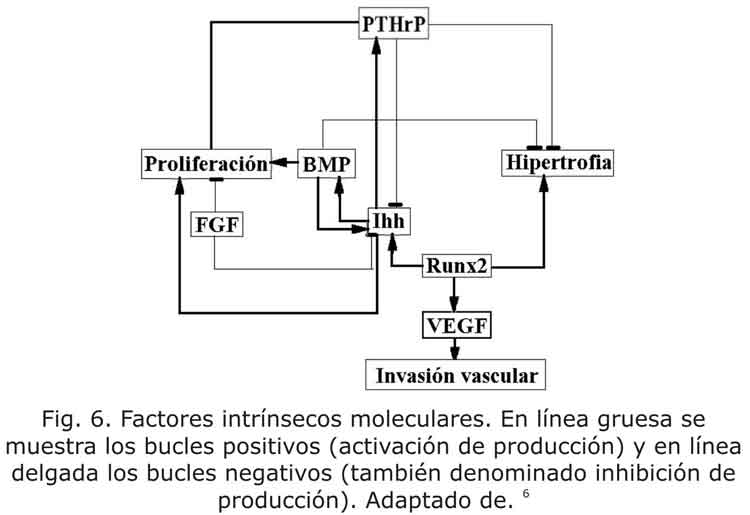
Los factores intrínsecos son aquellos que tienen una expresión localizada en la placa de crecimiento.1,4 En conjunto con las proteínas intrínsecas, las extrínsecas también actúan generando cambios localizados del proceso de proliferación, hipertrofia y apoptosis (fig. 5). En esta figura se observan los patrones moleculares y celulares que se presentan en un hueso largo.
Se han identificado numerosas hormonas locales que regulan y mantienen la proliferación de condrocitos en la placa epifisiaria. El primer paso para el proceso de desarrollo endocondral es la condrogénesis, que se regula por la familia de SOX (se observa la ubicación en la fig. 5), entre los que se cuentan las proteínas SOX9, L-SOX5 y SOX6, que son factores de transcripción.7,8 El SOX9 se requiere en las etapas secuenciales de la diferenciación, y es fundamental para que las células mesenquimales lleguen a ser condrocitos. Además, regula la formación de la matriz de colágeno, entre los que se encuentran el colágeno tipo II, el colágeno tipo IX y el tipo XI.6 Durante el crecimiento en los huesos largos el SOX9 se expresa por los condrocitos proliferantes y se inhibe por los condrocitos hipertróficos.7 Experiencias con ratones han demostrado que el SOX9 regula positivamente la proliferación y negativamente la hipertrofia.9 Análogamente el SOX9, el SOX5 y el SOX6 regulan los pasos del desarrollo endocondral. Además, se necesita la presencia de SOX9 para que exista la expresión de SOX5 y SOX6.
Una vez que se tiene el cartílago hialino primario comienza la osificación endocondral la cual está regulada por el cartílago de crecimiento. Un elevado número de proteínas han demostrado tener efectos sobre este cartílago, como son las proteínas de la matriz extracelular, las proteínas que regulan los ciclos celulares (ciclinas, Cdks), citoquinas (IL-1, IL-6, IL-10), factores de crecimiento (TGF-b, FGF, BMP), la familia hedgehog (Indian hedgehog), las metaloproteasas (MMP) y los factores angiogénicos y antiangiogénicos (VEGF, angiopoietin -2, METH-1).1,3
Principalmente, se piensa que tres moléculas regulan la proliferación de los condrocitos en la placa de crecimiento expresadas por los mismos condrocitos: 6,10
- el péptido recombinante de la hormona paratiroidea (PTHrP),
- el Indian hedgehog (Ihh),
- los factores transformantes (TGF-b).
Una gran variedad de tejidos producen la hormona PTHrP, entre los que se encuentran el cartílago, el corazón, el riñón, la placenta, los senos, en superficies epiteliales, entre otros. El PTHrP es un factor autocrino-paracrino que se une y activa al receptor PTH/PTHrP, que es un regulador de calcio/fostato, y de la remodelación ósea a través de las acciones sobre el riñón y el hueso.11,12 El PTH/PTHrP controla las acciones endocrinas del PTH y la acción auto/paracrina del PTHrP.
El receptor PTH/PTHrP es una proteína que se genera a bajos niveles por los condrocitos proliferantes columnares, y a altos niveles por los condrocitos pre-hipertróficos. A su vez, el PTHrP se expresa en el pericondrio y por células proliferativas.6 El PTHrP regula negativamente el paso del condrocito inmaduro proliferante a maduro hipertrófico y promueve la proliferación celular.
Por su parte, el Ihh es un miembro de una familia de proteínas importantes para la generación de patrones a nivel embrional, y las células proliferantes y pre-hipertróficas lo producen en grandes cantidades.1,3,6,10 El Ihh es necesario y suficiente para la expresión del PTHrP6 e inhibe, indirectamente, la diferenciación de condrocitos proliferantes a hipertróficos, gracias a que la expresión de Ihh permite generar PTHrP, el cual es el inhibidor directo de la hipertrofia.
El Ihh estimula el PTHrP el cual mantiene los condrocitos en proliferación; mientras, el PTHrP retrasa la producción de Ihh, lo cual constituye un "bucle" de retroalimentación negativo. 6,10
También al igual que el PTHrP, el Ihh es un potente estimulador de la proliferación de condrocitos.6 Alternativamente, el Ihh y el PTHrP coordinan la diferenciación de osteoblastos y determinan la longitud de la columnas proliferativas en cada hueso e individuo.1
Otro factor importante es el FGF receptor 3, que pertenece a la familia de los receptores kinasa-tirosinas que se unen a los FGF (factores de crecimiento fibroblástico). 6 A su vez los FGF son una familia de proteínas que desempeñan un papel crucial en múltiples actividades biológicas.6 Este factor FGFR3 se produce en la zona de los condrocitos proliferantes y en la zona columnar. Su importancia radica en que la alteración de la membrana del FGFR3 produce condrodisplasia e hipocondrodisplasia entre otras lesiones de retraso del crecimiento. Además, se producen miembros mucho más cortos. El 90% de los casos se atribuyen al incremento de la edad de paternidad. 4 Se han establecido modelos animales en los que el FGFR3 regula negativamente la proliferación, pero no es clara su acción sobre la hipertrofia. 13 En modelos in vitro se han establecido que el FGFR3 acelera la hipertrofia.14
Por otro lado se ha identificado que las proteínas morfogenéticas del hueso (BMP) desempeñan un papel importante en el desarrollo de diferentes partes del cuerpo6 y especialmente en el desarrollo de las extremidades. En modelos animales, especialmente en ratones, la BMP colabora en la formación de hueso ectópico, la condensación de cartílago y la formación de las articulaciones. 6 La BMP se produce en el pericondrio y en algunos casos se encuentra en los condrocitos hipertróficos (en especial la BMP2 y la BMP6). También, la BMP modula la proliferación y regula negativamente la hipertrofia.14 Las señales de BMP inducen la expresión de Ihh por los condrocitos pre-hipertróficos y aumentan tanto la proliferación de los condrocitos como la longitud de las columnas proliferativas. Las BMP y el factor de crecimiento fibroblástico (FGF) tienen efectos antagónicos. 1
Como se ha observado, el BMP, PTHrP y el Ihh retrasan la hipertrofia, pero no la regulan positivamente. Poco se conoce sobre los reguladores positivos del proceso hipertrófico.6 Sin embargo, se han realizado numerosas investigaciones sobre el RUNX2, el cual se expresa por los condrocitos hipertróficos, y regula positivamente la hipertrofia y la diferenciación de los osteoblastos. En conjunto con el VEGF, el RUNX2 desarrolla un papel importante en la invasión vascular del cartílago.
También se ha hecho referencia15 a la capacidad de regeneración y consolidación ósea después de una fractura en los adultos. Esto es el resultado de la actividad intrínseca de la reparación de los huesos largos que reside internamente en el hueso y que depende principalmente de la BMP. Específicamente la BMP2 es necesaria para controlar el inicio de la reparación de las fracturas. En15 se ha demostrado a partir del bloqueo de la producción de BMP2 en ratones, que las posibles fracturas no pueden repararse en el tiempo si no hay BMP. Efecto que, sin embargo, no se observa en la morfogénesis ósea.
Por su parte, el VEGF es el factor responsable y necesario para la vascularización de la placa epifisiaria. La ausencia de esta proteína produce importantes alteraciones en la arquitectura de la fisis, y afecta el crecimiento longitudinal del hueso. Por el contrario, la invasión de los capilares metafisarios regula la morfogénesis del cartílago de crecimiento y su remodelación.10 La invasión vascular del cartílago de crecimiento se correlaciona con la expresión en los condrocitos hipertróficos de VEGF.1,3,10
Las relaciones existentes entre las moléculas locales y la acción ejercida por cada una de ellas en la placa de crecimiento se puede observar en la tabla y en la fig. 6.
Factores extrínsecos (sistémicos) moleculares del crecimiento endocondral
En conjunto con los factores locales, los factores sistémicos como la hormona del crecimiento, la tiroidea, las gonadales, los estrógenos y los andrógenos, la vitamina D y los glucocorticoides, actúan mediando el crecimiento de los huesos largos.
La vitamina D, obtenida por la ingestión de leche, huevos y vegetales, permite la mineralización del osteoide y aumenta la actividad de la fosfatasa alcalina y de las MMP.1 Sin embargo, el efecto de la vitamina D sobre la placa de crecimiento es secundario a la luz de los efectos locales (autocrinos). Se desconoce si el efecto de la vitamina D sobre la calcificación es por acción directa del metabolismo sobre los condrocitos. 3
Otras hormonas sistémicas como la del crecimiento (GH) y la paratiroidea (PTH) actúan en la primera región de la placa epifisiaria (la zona proliferativa). Así, al extirparse la hipófisis de un animal en desarrollo, este deja de crecer porque las células de la región proliferativa dejan de dividirse, aunque sigue el proceso de calcificación en la zona metafisiaria con la disminución de trabéculas y el adelgazamiento de la placa de crecimiento.1
La hormona GH actúa sobre las células ubicadas en la zona de reserva que ayudan a la división celular y aumentan el grosor de la placa epifisiaria. Estudios en recién nacidos con insuficiencia de GH muestran tasas de crecimiento normal, lo que explica que otras moléculas son las que controlan el crecimiento durante la etapa prenatal.16 La tiroxina se encuentra íntimamente ligada al GH, y produce colágeno X por medio de la inducción local de BMP.
Por su parte, los niños hipotiroideos tienen un crecimiento defectuoso, a diferencia de los niños hipertiroideos que presentan un crecimiento acelerado del esqueleto.5 También las hormonas gonadales aumentan la vascularización en cortos espacios de tiempo lo que permite la aposición de nuevo tejido. 3
Durante el rápido crecimiento en la pubertad, hay una intensa acción del cartílago de crecimiento marcado por la acción de las hormonas sexuales y el incremento de la GH. La testosterona estimula inicialmente la división celular en la fisis. Los estrógenos parecen estimular el crecimiento del tejido óseo diferenciado y pueden frenar el crecimiento cartilaginoso afectando a la placa subcondral a cada lado de la fisis.1
Efecto de las cargas mecánicas
Los patrones iniciales durante el desarrollo embrional del esqueleto ocurren en ausencia de contracciones musculares.17 Es comúnmente aceptado que la morfogénesis en el desarrollo está únicamente controlada por los factores genéticos intrínsecos y la regulación local del ambiente químico del tejido en desarrollo.17
Se han analizado diversos efectos sobre el crecimiento de huesos largos por factores biofísicos, específicamente se ha realizado un estudio de los efectos de la carga mecánica sobre el hueso.
La mecánica estructural de los huesos en crecimiento se ha determinado mediante modelos experimentales en animales y humanos y a través de modelos computacionales de simulación (ver por ejemplo18). Estos modelos de simulación son fundamentales para evaluar los estados tensionales en los procesos de carga19 sobre los huesos en crecimiento. Así, Shefelbine y Carter19 determinaron que durante el crecimiento del hueso, el cartílago es particularmente sensible a las cargas mecánicas por su facilidad de deformación ante cargas sostenidas en el tiempo. Por lo tanto, cargas anormales sobre el hueso pueden desarrollar deformidades, como por ejemplo la displasia total de cadera. En esta afección, clínicos e investigadores están de acuerdo en que la causa de la displasia es la ubicación anormal de las cargas sobre la cabeza del fémur, debido a la posición de la extremidad, la presión de la articulación y la tensión de los ligamentos.
Shefelbine y otros20 también simularon, mediante un modelo de elementos finitos, el estado de carga durante la marcha humana en la posición de apoyo unipodal. Mediante este modelo se halló que el estado tensional en la fisis del fémur distal promueve un crecimiento más acelerado en la parte medial que en la lateral y se forma el ángulo bicondilar. Este modelo ha explicado el proceso de desarrollo del ángulo bicondilar de los niños en crecimiento.
Tanck y otros21 estudiaron las consecuencias de la mineralización sobre las propiedades mecánicas de un hueso a nivel embrionario. Para ello utilizaron modelos basados tanto en experimentos de laboratorio como en elementos finitos. Usando probetas de hueso embrional (rudimentos de hueso), se ensayaron muestras sometidas a flexión para ratones de 16 y 17 días de edad de gestación. Para la predicción del módulo de elasticidad se utilizó, de nuevo, el método de los elementos finitos usando los datos de fuerza-desplazamiento encontrados a nivel experimental. Con estos datos se constató que el módulo de elasticidad se eleva en dos órdenes de magnitud en función de la mineralización. 21
La inyección de agentes que bloquean el sistema neuromuscular en el embrión de pollo 17 ha sido utilizado para estudiar la inmovilización durante el desarrollo de las extremidades de este animal. Con esta parálisis artificial se puede separar el papel de las cargas mecánicas de la actividad biológica. Se concluyó 17 que las extremidades paralizadas tienen un sistema musculoesquelético normal que, sin embargo, requiere de las cargas mecánicas en la etapa postnatal para el correcto mantenimiento del mismo. Cabe señalar que la estructura de inserción del tendón y la rótula muestran un tamaño reducido en comparación con estructuras normales.
Sin embargo, durante el desarrollo postnatal, el ambiente mecánico controla la diferenciación del tejido a través de las deformaciones impuestas. Esta señal se traduce en un estímulo celular según el tipo y la magnitud de carga e impone un crecimiento controlado. Por lo tanto, las deformaciones generadas por el crecimiento y por la contracción muscular programan la hipertrofia y la osteogénesis. 17
Las tensiones cíclicas se han estudiado ampliamente por su similitud con las cargas fisiológicas de contracción muscular.17 Por su parte, la cargas de presión se han estudiado en huesos en desarrollo y se considera que gracias a estas se genera matriz extracelular 17 en el cartílago articular. De otro lado, Carter y otros22 simularon, mediante un modelo computacional, los patrones de deformación y tensión durante el proceso de distracción en la etapa de osteogénesis. El modelo fue capaz de predecir los patrones de diferenciación consistentes con el estudio histológico realizado en ratones. 17
El ambiente mecánico puede influir también en la dirección de crecimiento. En la etapa primaria del embrión, el ambiente químico y genético tiene una alta influencia sobre la dirección y velocidad de crecimiento. Sin embargo, con el tiempo, el citoesqueleto de las células se adapta a la dirección de la deformación y también se puede generar movimiento de las células en la matriz extracelular en dirección a las deformaciones principales. Por ello, el tejido óseo posee estructuras altamente direccionales.17,22
Se ha hecho una introducción de los efectos de las cargas mecánicas sobre el crecimiento y la placa epifisiaria. Cada uno de los efectos mecánicos afectan de forma diferente. En los siguientes apartados se lleva a cabo una recopilación y análisis de los principales factores mecánicos que regulan el crecimiento, en donde se puede anotar el ejercicio físico, la compresión excesiva de la placa epifisiaria (en personas con elevado sobrepeso), distracción, y efectos sobre el centro secundario de osificación.
Efecto del ejercicio físico
Si bien los índices de formación y remodelación de hueso se encuentran determinados por la genética, el cartílago de crecimiento es sensible a los cambios por las solicitaciones mecánicas.20 Las solicitaciones fisiológicas sobre el cartílago de crecimiento son necesarias para alcanzar un correcto desarrollo esquelético. El cartílago de crecimiento está sometido a solicitaciones provocadas por las fuerzas internas y externas. Entre las fuerzas internas hay que considerar el propio crecimiento fisario y el del núcleo de osificación secundario. Al crecer precisan espacio y colisionan con los huesos vecinos. Entre las fuerzas externas hay que considerar el periostio, el nódulo de Ranvier y los músculos, que crecen y se contraen con sus tendones insertados en el hueso.3
Dentro de los límites biológicos, el aumento de la tensión o de la compresión acelera el crecimiento, 5 mientras que por debajo de los límites fisiológicos el crecimiento puede verse disminuido.5 Estos principios son conocidos como ley de Hueter-Volkmann, propuesta inicialmente por Delpech en 1828,1 que señala la relación inversa entre las solicitaciones a compresión paralelas al eje longitudinal del cartílago epifisario y el índice de crecimiento epifisario.3 En 1984 Simon y Papierski 23 observaron que si la placa de crecimiento de ratas bípedas hipofisectomizadas se estimula con carga mecánica a compresión intermitente se prolonga el desarrollo del cartílago de crecimiento, por lo cual se concluye que el estímulo mecánico puede inducir el crecimiento biológico. Es importante notar que Henderson y Carter 17 afirman que durante el crecimiento, el desarrollo local de proliferación e hipertrofia genera deformaciones y tensiones remanentes en el hueso y el esqueleto que influyen en la morfogénesis por cualquiera de las siguientes formas:
- Al regular la acción de la velocidad de crecimiento
- Al regular la diferenciación de tejidos
- Al dirigir el crecimiento por las restricciones articulares
- Por deformar los tejidos
El rango de las cargas mecánicas o deformaciones sobre la fisis en humanos ha sido estimada en varias ocasiones. Bylski-Austrow y otros24 estimaron que la carga estática equivalente ejercida por la placa de crecimiento proximal de la tibia y la placa distal del fémur en adolescentes es aproximadamente de 500N, o que varía entre el 5 y el 25 % del peso del individuo. Por lo tanto, la tensión inducida en la placa de crecimiento es de aproximadamente 1 MPa. Para pacientes con tumores malignos Safran y otros. 25 calcularon una carga de 5,8 MPa. Cook y otros26 estimó mediante elementos finitos, la compresión de la fisis en la parte medial de una tibia para niños de 2 a 5 años y encontró que la tensión necesaria para el crecimiento es de 4 MPa.
Efecto de la compresión sobre la placa de crecimiento
Cuando se comprime la placa de crecimiento se produce una reducción de la zona de proliferación y se acelera la hipertrofia con una diferenciación de células y formación de columnas desordenadas.3 En la experimentación clínica, la fisis se estrecha al comprimirla mediante algún mecanismo externo (fijadores) o con grapas. Una vez que se anula el elemento externo se reinicia el crecimiento, aunque la tasa y el crecimiento total es impredecible.1
Sin embargo, el efecto de la compresión mecánica de baja magnitud es un tema de controversia en el que algunos autores creen que no existe inhibición del crecimiento ante pequeñas fuerzas compresivas.3 Lerner y otros27 probaron también mediante elementos finitos, que la compresión elevada en la placa de crecimiento se encuentra relacionada con menores índices de crecimiento.
Efectos de la distracción sobre el cartílago de crecimiento
Para corregir las disimetrías* y las desviaciones angulares o descompensaciones axiales se utiliza la distracción de la placa de crecimiento. Los modelos experimentales realizados en animales utilizan una distracción pequeña y a un ritmo lento han comprobado que se aumenta la longitud del hueso mediante una gran proliferación celular en la placa de crecimiento. Sin embargo, cualquier tensión sobre la fisis, de baja magnitud pero de tiempo prolongado, puede producir una separación entre la zona hipertrófica y la metáfisis.1
El aumento en la altura fisiaria originada por la tensión, antes de producirse la epifisiólisis (la separación de la metáfisis y la zona hipertrófica) puede explicarse como un problema de la irrigación sanguínea, ya que al acumularse los condrocitos en la capa hipertrófica sin aporte vascular y sin llegada de condrocitos por la isquemia metafisiaria, los vasos sanguíneos son alargados y tienen que recorrer una mayor distancia para alcanzar la zona hipertrófica de la placa de crecimiento, con lo cual se disminuye el diámetro y el flujo sanguíneo.3
Efecto de la desperiostización durante el crecimiento
El periostio tiene una función importante en la formación del hueso cortical y en la reparación de las fracturas. Este se encuentra adherido a los extremos epifisiarios del hueso en desarrollo, anclado en el anillo de Lacroix. Wolpert y otros28 afirman que el pericondrio dirige el desarrollo del cartílago de crecimiento. Además soporta las tensiones mecánicas para proveer estabilidad al hueso. Sin embargo, lo más importante es que la acción mecánica sobre el pericondrio puede activar señales de control molecular (desde y hacia el pericondrio), que causan alteraciones de la velocidad y dirección de crecimiento, y en la forma de diferenciación celular.17
La experimentación sobre la desperiostización ha sido importante para entender el comportamiento de la placa de crecimiento ante cargas mecánicas. Dimitriou y otros.29 realizaron pruebas con conejos en los cuales compararon el crecimiento del periostio en la dirección longitudinal y transversal y se concluyó que el crecimiento es solamente en dirección longitudinal, de donde se puede concluir que al desperiostizar se genera una elongación mayor del hueso. Otras pruebas de este efecto se obtuvieron con ratas, en las que se quitaron tiras de tejido perióstico y se aumentó el crecimiento un 1,5 %. También se encontró que el periostio genera deformaciones angulares importantes.1 Haasbeek y otros.30 señalan dos casos clínicos en donde se presentan deformaciones angulares al tensar el periostio cerca de la fisis. También, Rooney y Archer 31 han realizado experimentos de desperiostización en pollos y observaron un incremento de la longitud de un 16 % en comparación con los especímenes de control. Este incremento en longitud se puede deber a que la eliminación del periostio disminuye las cargas compresivas (inhibiendo el crecimiento, como se señaló al inicio de esta sección).
Como se señaló al comienzo, las cargas sobre los huesos largos determinan la dirección del crecimiento. En primera instancia, los condrocitos de la fisis (o placa de crecimiento) son orientados en formaciones columnares que crecen sobre un eje determinado. Sin embargo, Rooney y Archer 31 encontraron alteraciones en la dirección del crecimiento al quitar el pericondrio.
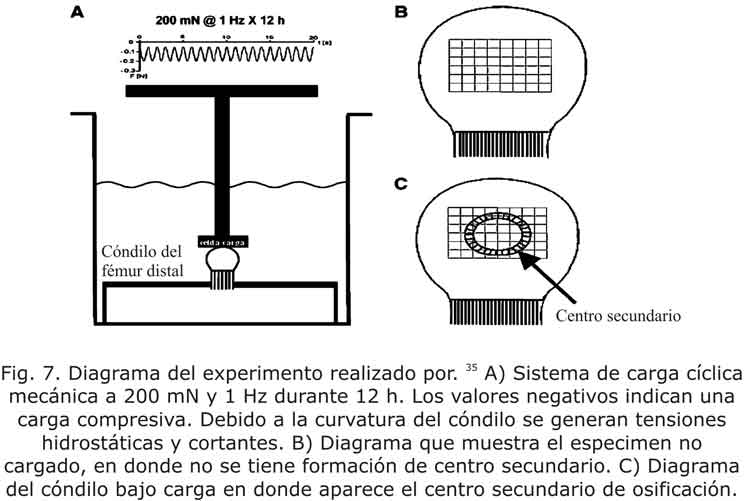
Efectos de la carga sobre el centro secundario de osificación
Los modelos teóricos propuestos por numerosos autores32-34 predicen que altas tensiones octaédricas y bajas tensiones hidrostáticas favorecen la osificación endocondral. Para validar estos modelos, Sundaramurthy y Mao35 presentan un estudio experimental sobre la osificación de la condroepífisis modulada por la carga mecánica sobre el cóndilo distal del fémur de ocho conejos (con 5 días de edad). Después de la disección, el cóndilo derecho del conejo fue extraído y puesto en una cámara de cultivo de órganos en un medio DMEM1 con 10 % de suero bovino fetal y 1 % de penicilina. Este fémur fue rígidamente fijado a la parte inferior de la cámara (fig. 7) y en la parte del cóndilo articular se aplicó una carga cíclica compresiva de 200mN y 1 Hz por un total de 12 h equivalente a 11200 ciclos. La carga fue aplicada con un sistema servohidráulico.
Durante las mismas 12 h en que el fémur derecho se encontraba bajo la acción de una carga cíclica, el fémur izquierdo fue convenientemente separado en un medio idéntico de conservación. Los dos fémures fueron puestos en un biorreactor rotatorio (20 r.p.m.) en un medio con nutrientes. 35
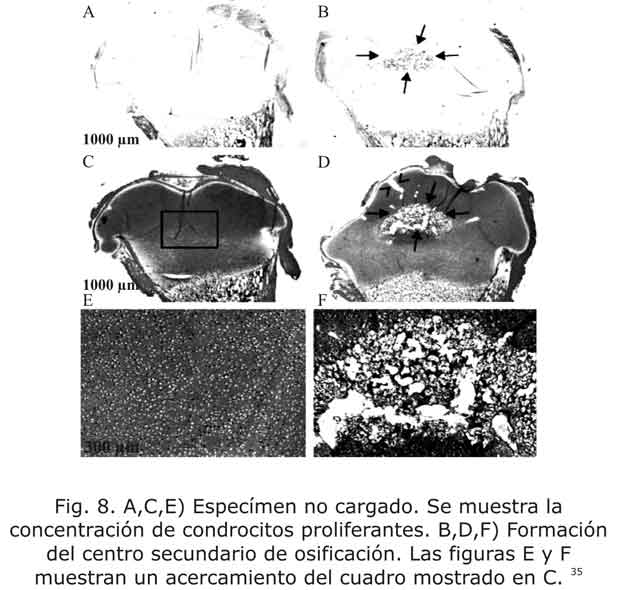
Posteriormente, los cóndilos fueron analizados microscópicamente. El análisis de todos los cóndilos no cargados (control) mostraron que el centro secundario de osificación no aparece en la epífisis. Por el contrario, en las muestras cargadas se observa la formación de una estructura de centro secundario (fig.8). Este centro presenta células hipertróficas y una pequeña proporción de hueso trabecular, en comparación con la aparición de condrocitos proliferantes en la condroepífisis no cargada.
Por su parte, la inmunolocalización reveló que la estructura donde se presenta el centro secundario tiene una alta expresión de RUNX2 solamente en el lugar donde se encuentra dicho centro y es baja la concentración en otros sitios del mismo cartílago epifisiario. Igualmente, la osteopontina fue intensamente expresada en el centro secundario. A diferencia de lo encontrado en el especímen cargado, los cóndilos de control no mostraron expresión de RUNX2 ni osteopontina. 35
También, en cuanto a la producción de colágeno tipo II y colágeno tipo X se encontraron diferencias apreciables. El colágeno tipo II (la mayor proteína estructural del cartílago) aparece en todo el fémur control, mientras que en el especímen cíclicamente cargado se encontró en la epífisis, a excepción del centro secundario. Con respecto al colágeno tipo X se vio únicamente expresado en la zona adyacente a los condrocitos hipertróficos. 35
En conclusión, las cargas mecánicas inducen la aparición del centro secundario de osificación en el fémur de conejos de pocos días de nacimiento. Una explicación alternativa para la formación de este centro establece que el linaje de las células que se encuentran en el centro de la epífisis cambian desde condrocitos hasta osteoblastos por la carga cíclica. 35
REFERENCIAS BIBLIOGRÁFÍCAS
1. Shapiro F, Forriol F. El cartílago de crecimiento: Biología y biomecánica del desarrollo. Rev Cubana Ortop Traumatol. 2005;49(1):55-67.
2. Schoenau E, Fricke O, Rauch F. The regulation of bone development as a biological system. Homo. 2003;54(2):113-8.
3. Forriol F, Shapiro F. Bone Development. Clinical Orthopaedics and Related Research. 2005;112:14-33.
4. Ballock RT, O'keefe RJ. The biology of the growth plate. J Bone Joint Surg Am. 2003;85A:715-26.
5. Ganong WF. Review of Medical Physiology (LANGE Basic Science) ed.21. USA: Hill Medical;2003.
6. Provot S, Schipani E. Molecular mechanisms of endochondral bone development. Biochemical and Biophysical Research Communications. 2005;328(3):658-65.
7. DeCrombrugghe B, Lefebvre V, Nakashima K. Regulatory mechanisms in the pathways of cartilage and bone formation. Current Opinion in Cell Biology. 2001; 6(13):721-7.
8. Lefebvre V. Toward understanding the functions of the two highly related sox5 and sox6 genes. Journal of Bone And Mineral Metabolism. 2002;20(3):121-30.
9. Akiyama H, Chaboissier MC, Martin JF, Schedl A, DeCrombrugghe B. The transcription factor sox9 has essential roles in successive steps of the chondrocyte differentiation pathway and is required for expression of sox5 and sox6. Genes and Development. 2002;16(21):2813-28.
10. Kronenberg HK. Development regulation of the growth plate. Nature. 2003; 423:332-6.
11. Potts JH, Kronenberg HM, Rosenblatt M. Parathyroid hormone: chemistry, biosynthesis, and mode of action. Advances in Protein Chemistry; 1982:35323-96.
12. Strewler GJ. The physiology of parathyroid hormone-related protein. New England Journal of Medicine. 2000;342:177-85.
13. Ornitz DM, Marie PJ. Fgf signaling pathways in endochondral and intramembranous bone development and human genetic disease. Genes and Development. 2002;16(12):1446-65.
14. Minina E, Kreschell C, Naski MC, Ornitz DM, Vortkamp A. Interaction of fgf, ihh/pthlh, and bmp signaling integrates chondrocyte proliferation and hypertrophic differentiation. Developmental Cell. 2002;3(3):119-29.
15. Tsuji K, Bandyopadhyay A, Harfe BD, Cox K, Kakar S, Gerstenfeld L, et al. BMP2 activity, although dispensable for bone formation, is required for the initiation of fracture healing. Nat Genet. 2006;38(12):1424-9.
16. Van der Eerden BCJ, Karperein M, Wit JM. Systemic and local regulation of the growth plate. Endocrine Rev. 2003;24:782-801.
17. Henderson JH, Carter DR. Mechanical induction in limb morphogenesis: the role of growth-generated strains and pressures. Bone. 2002;31(6):645-53.
18. Carter DR, Van der Meulen MCH, Beaupre GS. Mechanical factors in bone growth and development. Bone. 1996;18(1):5S-10S.
19. Shefelbine SJ, Carter DR. Mechanobiological predictions of growth front morphology in developmental hip dysplasia. Journal of Orthopaedic Research. 2004;22(2):314-52.
20. Shefelbine S, Tardieu C, Carter DR. Development of the femoral bicondylar angle in hominid bipedalism. Bone. 2002;30(5):765-70.
21. Tanck E, Van Donkelaar CC, Jepsen KJ, Goldstein SA, Weinans H, Burger EH, et al. The mechanical consequences of mineralization in embryonic bone. Bone. 2004;35(1):186-90.
22. Carter DR, Beaupre GS, Giori NJ, Helms JA. Mechanobiology of Skeletal Regeneration. Clin Orthop. 1998;355(Suppl):S41-55.
23. Simon MR, Papierski P. Effects of experimental bipedalism on the growth of the femur and tibia in normal and hypophysectomized rats. Acta Anat. 1982; 114:321-9.
24. Bylski-Austrow DI, Wall EJ, Rupert MP, Roy DR, Crawford AH. Growth plate forces in the adolescent human knee: A radiographics and mechanical study of epiphyseal staples. J Pediatr Orthop. 2001;21:817-23.
25. Safran MR, Eckardt JJ, Kabo JM. Continued growth of the proximal part of tibia after prosthetic reconstruction of the skeletally immature knee: Estimation of the minimum growth force in vivo humans. J Bone Joint Surg. 1992;74A:1172-9.
26. Cook SD, Lavernia CJ, Burke SW. A biomechanical analysis of the etiology of tibia vara. J Pediatr Orthop. 1983;3:129-34.
27. Lerner Al, Kuhn JL, Hollister SJ. Are regional variations in bone growth related to mechanical stress and strain parameters? J Biomech. 1998;31:327-35.
28. Wolpert L. Cell Behaviour-Cartilage morphogenesis in the limb. Cambridge: Cambridge University;1982.
29. Dimitriuo GA, Kapetanos CG, Symeonides PP. The effect of partial periosteal division on growth of long bones. An experimental study in rabbits. Clin Orthop. 1998;236:265-9.
30. Haasbeek JF, Rang MC, Blackburn N. Periosteal tether causing angular growth deformity: Report of two clinical cases and an experimental model. J Pediatr Orthop. 1995;15:677-81.
31. Rooney P, Archer CW. The development of the perichondrium in the avian ulna. J Anat. 1992;181:393-401.
32. Carter DR. The regulation of Skeletal biology by mechanical stress histories, In: Tissue Engineering. New York: CF FOX;1988.
33. Carter DR, Wong M. The role of mechanical loading histories in the development of diarthrodial joints. J Orthop Rev. 1998;6:804-16.
34. Stevens S, Beaupre G, Carter DR. Computer model of endochondral growth and ossification in long bones: Biological and mechanobiological influences. Journal of Orthopaedic Research. 1999;17:614-53.
35. Sundaramurthy S, Mao J. Modulation of endochondral development of the distal femoral condyle by mechanical loading. Journal of Orthopaedic Research. 2006; 24:229-41.
Recibido: 5 de enero de 2008.
Aprobado: 3 de marzo de 2008.
M.Sc. Diego Alexander Garzón Alvarado. Universidad Nacional de Colombia. Ciudad Universitaria de Colombia, Bogotá. Correo electrónico: dagarzona@unal.edu.co
*Diferencias de longitudes entre 2 huesos con simetría, ejemplo: 2 tibias, 2 fémur, etcétera.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}