










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. v.22 n.1 La Habana ene.-abr. 2007
Trabajo original
COMPETENCIA DE Diaphorina citri KUWAYAMA Y Phyllocnistes citrella STAINTON EN EL AGROECOSISTEMA CITRÍCOLA DE LA ISLA DE LA JUVENTUD, CUBA
COMPETENCE OF Diaphorina citri KUWAYAMA AND Phyllocnistes citrella STAINTON IN THE CITRUS AGROECOSYSTEM OF ISLA DE LA JUVENTUD, CUBA
Miriam Fernández*, Ileana Miranda* y María Elena Díaz**
*Grupo Plagas Agrícolas. Dirección de Protección de Plantas. Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana, Cuba. CP 32700. Corrreo electrónico: mfdez@censa.edu.cu; **Centro Universitario Jesús Montané Oropesa. Carretera aeropuerto km 31 ½. Isla de la Juventud, CP 27200
RESUMEN
Phyllocnistis citrella (Lepidoptera: Gracillariidae: Phyllocnistinae) (MHC), uno de los fitófagos más perjudiciales para los cítricos, se detectó por primera vez en la Isla de la Juventud en diciembre de 1993 y posteriormente, en febrero de 1999, a Diaphorina citri Kuwayama (Hemiptera: Psyllidae), especie de igual importancia para el cultivo. La disminución de la población del minador en el tiempo conjuntamente con la creciente población del psílido supuso la existencia de desplazamiento competitivo de esta especie sobre la otra. Para verificarlo se muestreó un campo, de dos años de edad (fomento), de naranjo dulce (Citrus sinensis (L.) Osbeck var. Valencia), desde marzo del 2002 hasta marzo del 2003. Se extrajo semanalmente un brote vegetativo foliar incipiente, de 30 plantas fijadas previamente al azar y se cuantificó la población total viva de P. citrella y de D. citri. Se empleó el modelo de Lotka _ Volterra de competencia interespecífica para su comprobación. La salida del modelo arrojó que D. citri despazó a P. citrella al determinarse que la isoclina correspondiente a la competencia intraespecífica del minador tiene menor inclinación que la isoclina correspondiente a la competencia interespecífica del psílido, lo cual fue corroborado con el valor de las densidades poblacionales de ambos fitófagos en el tiempo.
Palabras clave: Diaphorina citri; Phyllocnistis citrella; desplazamiento competitivo.
ABSTRACT
Phyllocnistis citrella Stainton (Lepidoptera: Gracillariidae: Phyllocnistinae), (MHC) is one of the most dangerous phytophages in Citrus. It first was detected in Isla de la Juventud on December 1993 and later, Diaphorina citri Kuwayama (Hemiptera: Psyllidae) invaded the crop on February 1999. This insect is as dangerous as P. citrella. The low population density of this pest and the high population density of D. citri suggested the competitive displacement between both species. In order to verify this, a two year old field was sampled weekly; and live miners and psylid populations on the buds of 30 plants previously fixed at ramdon were counted from March 2002 to March 2003. Lotka_Volterra´s interspecific competence model was used. The model out come showed that Phyllocnistes citrella was displaced by D. citri because the steepness of the interespecific line (D. citri) lied above that intraespecific line (Phyllocnistes citrella). Plotting population densities of both species were proved in the time.
Key words: Diaphorina citri; Phyllocnistis citrella; competitive displacement.
INTRODUCCIÓN
Los Ecólogos le conceden gran importancia al efecto que una especie puede ejercer sobre otra, cuando se compite por los mismos recursos del ambiente (13,14,15,23,24). Dos especies con idénticos nichos ecológicos (homólogos ecológicos) podrían estar sujetos a la misma competencia por el alimento y el espacio y la coexistencia sería imposible (25,29). Cuando dos especies, genéticamente diferentes tienen similar, pero no idénticos nichos ecológicos (no_homólogos ecológicos), y una de ellas es ligeramente superior a la otra, la selección natural discriminará contra la menos eficiente, presumiblemente menos fecunda y así eventualmente la elimina (4,21,22).
Phyllocnistis citrella Stainton (Lepidoptera: Gracillariidae: Phyllocnistinae), originaria del sudeste asiático y conocida comúnmente como el minador de la hoja de los cítricos (MHC), se detectó en Cuba, por primera vez, en el período de setiembre-noviembre de 1993, en la región occidental. En la Isla de la Juventud se halló en diciembre del mismo año, produciendo los mayores daños en plantaciones comerciales de fomento con 56,3% de infestación (González, C., comunicación personal, 2005)1. Este microlepidóptero es considerado uno de los fitófagos más perjudiciales al cultivo, porque con una afectación de más del 30% de la hoja dañada, produce pérdida en el peso de los frutos de la próxima cosecha y caída de los órganos de la planta. Las lesiones originadas favorecen las vías de acceso de Xantomona citri pv. campestri (Hasse) Dowson, agente causal del cáncer de los cítricos (12). Este fitófago, luego de su detección, rápidamente se estableció como residente en la entomofauna citrícola, tal como ha ocurrido en la mayoría de los países que ha invadido (2,3,10,35).
En febrero de 1999, fue informada por primera vez en Cuba, Diaphorina citri Kuwayama (Hemiptera: Psyllidae), el psílido asiático, plaga de igual importancia para los cítricos (17), ya que acude, al brote vegetativo foliar, produciéndole necrosis por la succión de grandes cantidades de savia (17,18). Este insecto, además de dañar por sí solo al cultivo, es un eficaz transmisor de una devastadora enfermedad exótica de origen bacteriano, el huanglongbing, greening, o likubin de los cítricos (1,2,3,17).
La preferencia mostrada por la brotación vegetativa foliar (entre 80 y 100 %), en punta de lanza por ambos fitófagos, así como una marcada tendencia a la disminución de las poblaciones del minador de los cítricos, supuso que se estaba produciendo desplazamiento competitivo de una especie por la otra. La comprobación de esta interacción ecológica, constituyó el objetivo del presente trabajo.
MATERIALES Y MÉTODOS
El experimento se realizó en un campo de fomento de naranjo dulce Citrus sinensis (L.) Osbeck var. Valencia sobre patrón Citrus volkameriana Ten., de dos años de edad, con un marco de plantación de 8 x 5 m, en suelo ferralítico amarillo lixiviado, perteneciente al lote 44 de la Empresa de Cítricos de la Isla de la Juventud, desde marzo del 2002 hasta marzo del 2003. Las observaciones se realizaron, semanalmente, en 30 plantas fijas, seleccionadas previamente al azar. En cada planta se analizó un brote vegetativo foliar incipiente, se anotó el estado fenológico (11), longitud del brote y se cuantificó, mediante el auxilio de un lente de aumento, la población total de P. citrella y de D. citri. En ambas especies se consideró la población viva.
Para describir la interacción competitiva entre ambas especies, las cuales presentan generaciones continuas, se utilizó el modelo de Lotka_Volterra de competencia interespecífica (29):

donde:
K1: densidad máxima de la población o capacidad de carga de D. citri
K2: densidad máxima de la población o capacidad de carga del P. citrella
rd1: tasa intrínseca de incremento de D. citri en estas condiciones específicas
rd2: tasa intrínseca de incremento de P. citrella en estas condiciones específicas
w1: coeficiente de competición de D. citri (representa el efecto per cápita de esta especie sobre P. citrella)
w2: coeficiente de competencia de P. citrella (representa el efecto per cápita de esta especie sobre D. citri)
Este modelo asume el efecto que se produce durante el ciclo de vida de los competidores y le adjudica mayor fuerza a la interacción porque no hay variación con la edad.
Las propiedades clásicas del modelo de Lotka Voterra de competencia entre especies están determinadas mediante el examen de las isoclinas y2 = w2 x + k2 y y1 = w1 x + k1 y2, las cuales se obtienen cuando las poblaciones dejan de crecer, entonces dN1/dt = 0 (D. citri) y dN2/dt = 0 (P. citrella). Los valores de los coeficientes de competencia empleados fueron: w1 = 2 (D. citri) y w2 = 0.25 (P. citrella). Los gráficos descritos se ajustaron a los resultados de la salida obtenida para el cálculo de las isoclinas (29). Se realizó un histograma que grafica las densidades de población en el período evaluado. Todos los análisis se realizaron con el paquete estadístico Statistica (28).
RESULTADOS Y DISCUSIÓN
Los resultados alcanzados demuestran que la incorporación de D. citri en la entomofauna cubana ha producido un desplazamiento competitivo en P. citrella, como lo indican los valores de K y rd de los competidores. En la Tabla 1 se observa que la capacidad de carga o máxima densidad de la población de D. citri (K1) es mucho mayor que la de P. citrella (K2), lo que significa que el psílido alcanza la posición de equilibrio con mayor densidad. Resultado contrario se produce en la población del minador. Las tasas neta de crecimiento instantáneo (rd1 = 0.45 y rd2 = 0.05) de las dos poblaciones, en estas condiciones, evidencian la tendencia al crecimiento de D. citri y al decrecimiento de P. citrella.

Las isoclinas representan la competencia intraespecífica del minador (N2) y la competencia interespecífica del chupador (N1), respectivamente. Nótese que la isoclina del minador queda por debajo del efecto de competencia del chupador, la cual representa que el efecto competitivo interespecífico del psílido supera al efecto competitivo intraespecífico del minador (Fig. 1).

La inclinación de las isoclinas calculadas es la medida del efecto de la competencia interespecífica ejercida por el mejor competidor y aquella especie que tenga la menor inclinación es la perdedora, y por tanto, desplazada.
La base de este mecanismo es simple (31,32). La especie ganadora es aquella que produce mayor cantidad de descendientes hembras, la cual sobrevive para reproducirse por unidad de tiempo (34). Las hembras de D. citri son capaces de ovipositar más de 800 huevos durante toda su vida (20) y P. citrella apenas un total de 120 como máximo (35). En ambas especies la relación sexual es 1:1 (26).
Este hecho, unido a que D. citri sólo usa el brote durante todas las fases fenológicas (punta de lanza, despliegue, alargamiento y crecimiento) para completar su ciclo de desarrollo, hace que acuda al brote por mecanismos de reconocimiento y hallazgo del substrato más eficientes (9,16). P. citrella acude al brote en punta de lanza cuando este tiene alrededor de 5 mm (5); mientras que D. citri lo hace con apenas un 1mm de longitud (6,7,8). Estas características, entre otras, hicieron que D. citri se convirtiera en un fuerte competidor.
Las larvas de P. citrella, durante el proceso de alimentación, requieren de espacio suficiente para elaborar sus minas en serpentina cuando consumen el tejido vegetal. Por esa razón, el recurso espacio- alimento es vital para culminar este estado, de tal manera, que se ha constatado que cuando en una hoja inician su desarrollo más de tres larvas, sólo una, a lo sumo dos, logran pasar a pupa (5,12). En virtud de disponer de mayor espacio, el minador, cuando ataca al fruto, utiliza preferentemente, la zona ecuatorial, de mayor amplitud, que favorece el desarrollo de los juveniles (5).
Dentro del patrón conductual del minador no se puede soslayar que la hoja, también, tiene importancia vital para la formación del cocón, puesto que utilizan los bordes o los repliegues del limbo para junto con los hilos de seda que excreta la prepupa, formar el pupario. Por eso, cuando la larva se desarrolla en los vástagos o en el fruto, la prepupa salta en la búsqueda de hojas adyacentes u hojarasca del suelo para transformarse en pupa (5).
Estos requerimientos y hábitos de las larvas y pupas del microlepidóptero hacen que eviten el gregarismo en la superficie de la hoja, de ahí que muestren mayor competencia intraespecífica, que las ninfas chupadoras del hemíptero, las cuales permanecen en el brote, durante su desarrollo, apiñadas. En la medida que crecen las ninfas del psílido, crece el brote y así se logra la sincronización y compensación entre el brote y el fitófago (9). De esta manera, el efecto competitivo que este ejerce supera al intraespecífico del minador.
Este comportamiento de las especies se refleja más claramente cuando a través del modelo se grafica la tendencia del crecimiento de ambas poblaciones (N1 y N2) en el tiempo (Fig. 2).
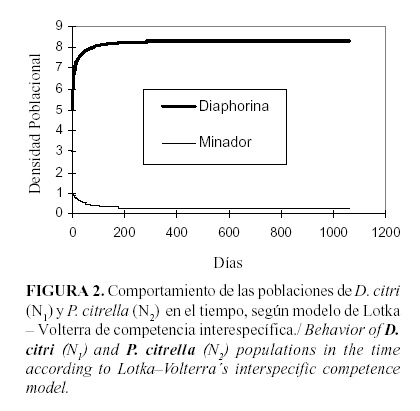
Como puede observarse, las poblaciones parten de un valor inicial de población, en el caso del invasor (N1 ) crece sostenidamente hasta llegar a su valor máximo de densidad, la densidad de equilibrio (K) y la especie desplazada (N2); prácticamente, desde el inicio decrece.
Generalmente, los efectos de la competencia interespecífica no se detectan de inmediato (22), sin embargo, en este caso particular se ha podido constatar el desplazamiento competitivo paulatino y efectivo, ejercido sobre el minador, prácticamente, en cinco años. Tal es así que muchos de los trabajos que se han tratado de iniciar con las poblaciones del minador en campo, se han visto postergado por la abundancia y constancia del psílido en las plantaciones (González C., comunicación personal, 2005).
Trabajos similares se han informado entre especies de diferentes grupos taxonómicos (no homólogos ecológicos) en los cuales se ha evidenciado el desplazamiento de una especie por otra (32).
En el desplazamiento competitivo, el ganador puede no ser siempre la misma especie porque los factores del ambiente en determinada localidad puede ser más favorable para aquella especie que frente a determinado recurso sucumbe ante el invasor. En estas interacciones pueden estar involucradas diferencias de temperatura, humedad relativa, enfermedades, pH, calidad del alimento y hasta la irradiación (30). De modo que la competencia interespecífica opera en las comunidades de los artrópodos dando lugar a numerosas salidas.
Los eventos de desplazamientos tienen importancia práctica y teórica. Desde ambos puntos de vista, el desplazamiento de una especie deja un hueco en el ambiente y altera la historia evolutiva de las especies que interactúan (19), así como de las especies asociadas como son los enemigos naturales (4). Además, la frecuencia de esos eventos de desplazamientos incrementan el grado de intervención antropogénica al ambiente, como son la introducción e invasión de especies exóticas para atenuar el desequilibrio que se produce y la aplicación de insecticidas químicos.
Reitz y Trumble (27) modificaron la definición de desplazamiento competitivo elaborada por DeBach (4), los cuales lo definen como la eliminación de una especie establecida en un hábitat por el uso, adquisición o defensa superior de los recursos por otra especie y formulan diferentes mecanismos por los cuales se producen factores que median en este tipo de competencia. Aunque la mayoría de los desplazamientos se producen por la invasión de especies exóticas pueden existir factores ambientales que predispongan a las especies a ser desplazadas como son la polifagia y la pérdida de enemigos naturales.
Se han informado ocho mecanismos generales de desplazamiento competitivo (27). En el caso que se estudia están involucrados cinco de estos mecanismos:
- Adquisición diferencial del recurso. En esta forma de explotación competitiva todos los individuos tienen acceso potencial a los recursos disponibles como ocurre con D .citri que por ser paurometábola, desarrolla el ciclo de vida desde que se inicia el crecimiento del brote (33).
- Fecundidad diferencial de la hembra. Existe fecundidad diferencial entre ambas especies, D .citri es capaz de producir hasta 800 huevos, mientras que el minador nunca ovoposita más de 120.
- Habilidad de búsqueda diferencial. Si D. citri arriba al brote cuando aún es un primordio (1mm) y es de necesidad vital para la especie esa condición, el fenómeno detectabilidad-confiabilidad es más competente que el del minador.
- Degradación del recurso. D. citri durante la alimentación ingiere grandes cantidades de savia, que debilita el substrato y, por otro lado, favorece el crecimiento de la fumagina, que coadyuva a que se produzcan daños irreversibles, dejando el recurso completamente degradado.
- Interferencia reproductiva. La degradación del brote atacado imposibilita la oviposición del minador por lo que se produce interferencia reproductiva.
En la Figura 3 se muestra la densidad de las poblaciones de ambos insectos en la misma unidad de muestreo, en ella se aprecie que con una densidad muy baja (< 5 individuos), la población de P. citrella solamente se expresa en los meses óptimos para su desarrollo.
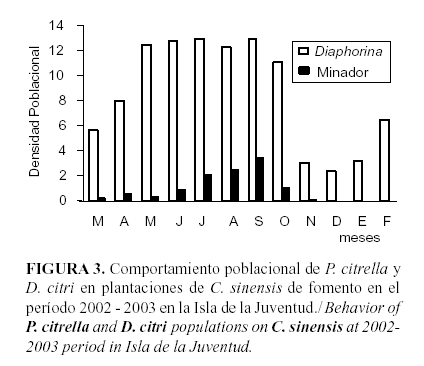
Mientras que D. citri, prácticamente, hace una gran meseta con densidades muy superiores a las del minador. Si bien, no se hace referencia a la extinción, hasta el presente, porque aún es factible la presencia en los campos del minador, sí se asegura que ha habido una significativa disminución de sus poblaciones por el efecto competitivo. Los resultados que ofrece el modelo empleado y lo observado, nacionalmente,
en los campos de cítricos así lo acreditan (Cueto, J., Comunicación personal)2
La competencia interespecífica ha acontecido en diversos taxa y se señala que las especies exóticas han desplazado en un 33% a especies nativas y en un 55% a otras especies exóticas, previa colonización, como ocurre en el caso que se estudia (27).
Desde una perspectiva ecológica, los desplazamientos proporcionan oportunidades para examinar los mecanismos de competencia interespecífica y los impactos que de ella puedan derivarse (22). El alcance e impacto de la competencia entre estos dos fitófagos en el agroecosistema citrícola, a lo largo de todo el país, requiere un análisis del comportamiento de ambas poblaciones. P. citrella exhibe una amplitud de nicho mayor que D. citri, ya que acude, preferentemente, al brote, pero puede utilizar los vástagos y el área ecuatorial de los frutos. La presión de selección ejercida por el psílido sobre el minador pudiera traer como consecuencia un proceso de especiación simpátrica (19), lo que supuestamente acarrearía variabilidad genética en sus poblaciones, fenómeno observado en el complejo Bemisia tabaci Genn.(Hemiptera: Aleurodidae) (mosca blanca) que ha traído serias dificultades en hortalizas (27). Por otra parte, su comportamiento poblacional podría devenir en metapoblaciones (subpoblaciones) dando lugar a una dinámica de caos.
D. citri, como es un fuerte competidor, y hasta el presente no se ha comprobado un efectivo control biológico, su tendencia sería la de crecer, lo que implicaría una fuerte acción antropogénica mediante la utilización del control químico que conllevaría un alto costo monetario y ambiental. Además hay que tener en cuenta, que es un eficaz vector del greening, enfermedad exótica para Cuba, que se ha informado en Florida, Estados Unidos (18).
Las invasiones de diferentes plagas en los cítricos durante los últimos años requiere un análisis por parte de los citricultores cubanos para continuar con las medidas que se han venido implementando. Conocer las relaciones que se establecen en comunidades más enriquecidas devienen en criterios más sólidos para el control de plagas.
Si bien las poblaciones de D. citri están creando alarma entre los productores y fitosanitarios no se le debe dar paso al control químico, debe profundizarse en el complejo de enemigos naturales por regiones y protegerlos, creando mecanismos que favorezca el incremento de la fauna beneficiosa y el manejo de la poda. Debe prevenirse el crecimiento excesivo mediante el empleo y manejo de su control natural (6, 18), por lo es necesario su protección y utilización.
A partir de los resultados obtenidos en este trabajo y considerando lo propuesto en el paquete de Instrucciones en Tecnología para los cítricos en la Isla de la Juventud, se propone que se tengan en cuenta estos resultados para coadyuvar en el perfeccionamiento de los procedimientos establecidos.
REFERENCIAS
1. Bellis G, Hollis D, Jacobson Sarah. Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae) and huanglugbing disease do not exist in the Stapleton station area of the Northern Territory of Australia. Austral J Entomol. 2005;44(1):68-70.
2. Bové JM, Danet JL, Bananej K, Hassanzadeh N, Taghizadeh M, Salehi M, Gamier M. Witches' broom disease of time (WBDL) in Iran. Proc 14 th Conf Int Org Citrus Virol. 2000;207-212.
3. Cermeli M, Morales P, Godoy F. Presencia del psílido asiático de los cítricos Kuwayama (Hemiptera: Psyllidae) en Venezuela. Entomol Venezolana. 2000;15(2):235-243.
4. De Bach P. The competitive displacement and coexistence principles. Ann Rev Ent. 1966;(11):183-212.
5. Díaz María Elena, Fernández Miriam, Gómez Josefina, García L. Phyllocnistis citrella Stainton (Lepidoptera: Gracillariidae). Daños en frutos de exportación en la Isla de la Juventud. Rev Protección Veg. 2006;21(1):27-30.
6. Etienne J, Quilicis S, Marival D, Franck A. Biological control of Diaphorina citri (Homoptera: Psyllidae) in Guadeloupe by imported Tamarixia radiata (Hymenoptera: Eulophidae). Fruits. 2001;(56):307-315.
7. Fernández Miriam, Miranda Ileana. Comportamiento de Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Parte I: Características morfológicas, incidencia y enemigos naturales asociados. Rev Protección Veg. 2005; 20(1):27-31.
8. Fernández Miriam, Miranda Ileana. Comportamiento de Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Parte II: Conducta preferencial con relación a la fenología del cultivo. Rev Protección Veg. 2005;20(2):22-24.
9. Fernández Miriam, Miranda Ileana. Comportamiento de Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Parte III: Relación entre el ciclo de vida y el brote foliar vegetativo. Rev Protección Veg. 2005;20(3):161-164.
10.French JV, Kahlk ECJ, da Graça JV. First Record of the Asian Citrus Psylla, Diaphorina citri Kuwayama (Homoptera:Psyllidae), in Texas. Subtrop Plant Science. 2001;(53):14-15.
11.Frómeta EM, Torres MO. Fenología Citrícola Tropical. Su importancia. Bol. Reseña. CIDA;1990.
12.Garrido A. El minador de las hojas de los cítricos: Morfología, biología, comportamiento, daños, interacción con factores foráneos. Phytoma. 1995;(72):84-92.
13.Graham M, Dayton P. On the evolution of ecological ideas: paradigms and scientific progress. Ecology. 2002; (83):1481-1489.
14.Gotelli N. Research frontiere in null model analysis. Global Ecology and Biogeography. 2002;(10):337-341.
15.Haila Y, Taylor H. The Phylosophical Diellness of Classical Ecology and a Levinsian Alternative. Biol and Phylos. 2001;(16):93-102.
16.James H, Junwang J, Ying H. Seasonal Abundance of the Asian Citrus psyllid, Diaphorina citri (Homoptera: Psyllidae) in Southern Florida. Florida Entomol. 2002;85(3):1-6.
17.Knapp JL, Halbert SE, Lee R, Hoy MA, Clark R, Kesinger M. The Asian Citrus Psyllid and Citrus Greening Disease. Florida IPM; 2002. (Consultada: 12 nov 2005). Disponible en: http://biocontrol.ifas.ufl.edu/ctgysrch/ASP-hoy.htm.
18.MacFarland C, Hoy Margorie. Survival of Diaphorina citri (Homoptera: Psyllidae) and two parasitoids Tamarixia radiata (Hymenoptera: Eulophidae) and Diaphorencirtus aligarhensis (Hymenoptera: Encyrtidae). Florida Entomologist. 2001;84(2): 227-233.
19.Mayr E. Ecological factors in speciation. Evolution. 1947;(1):163-88.
20.Mead FW. FDACS/DPI Entomology Circular No. 180; 2002. (Consultada: 12 nov 2005). Disponible en: http://emdept.ifas.ufl.edu.Mead.
21.Mendez A, Labajos L, Mesa J, de Zaldivar Ines, Mesa J. Template for producing a Camera-ready. Manuscrito para ICTE; 2003.
22.Mills NJ. Interspecific Competition in Insects. In:Huffaker CB, Gutierrez F, editors. Ecological Entomology. 2nd Ed. John Wiley & Sons Inc., Chapter 11. Part III;1999.p. 355-387.
23.Nudler O. Campos controversiales: hacia un modelo de su estructura y dinámica. Rev Patagonia Filos. 2002;3(1):9-22.
24.Nudler O. Hacia un modelo de cambio conceptual: espacios controversiales y refocalización. Rev de Filosofía Universidad Complutense de Madrid. 2004;(29):7-19.
25.Núñez M, Núñez Paula. Controversias en ecología: la competencia, de la certeza a la pregunta. Ecol Austral. 2005;(15):1-10.
26.Peña J, Duncan B. Seasonal abundance of Phyllocnistis citrella (Lepidoptera: Gracillaridae) and parasitoids in South Florida citrus. Environ Entomol. 1996;25(3):698-702.
27.Reitz S, Trumble J. Competitive Displacement Among Insects and Arachnids. Ann Rev Entomol. 2002;(47):435-465.
28.Statsoft. Statistica for Windows, versión 6.0.;1998.
29.Sharov A. Quantitative Population Ecology. (Course).11.2. Competition between Species. Dept. of Entomol.Virginia Tech, Blacksburg, VA; 1999. (Consultada: 2 sep 2005). Disponible en: http://www.gypsymoth.ento.vt.edu/~sharov/PopEcol/popecol.html.
30.Slobodkin LB. (1994): Simplicity and Complexity in the Games. Harvard University Press, Cambrige and London; 1994.
31.Slobodkin LB. The good, the bad and the reified. Evol Ecol Res. 2001;(3):1-13.
32.Torres R, Rodríguez M, Tijerina M, Flores A. Competencia de Sitophilus zeamaiz (Motschullsky) y Prostephanus truncatus (Horn) por alimento. Respyn. (Edición Especial). 2004;(6):1-2.
33.Tsai JH, Liu YH. Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plants. J Econ Entomol. 2000; 93(6):1721-1725.
34.Tsai JH, Wang JJ, Liu YH. Seasonal abundance of the Asian Citrus Psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae) in Southern Florida. Florida Entomol. 2002; 85(3):446-451.
35.Urbaneja A. Dinámica e impacto de los enemigos naturales del minador de las hojas de los cítricos, Phyllocnistis citrella Stainton. En: Actas de la VII Jornada Científica de S.E.E.A. Congreso Nacional de Entomología Aplicada. Junta de Andalucía, Consejería de Agricultura y Pesca. Congresos y Jornadas. España; 2004.
(Recibido 14-11-2005; Aceptado 10-7-2006)
1 González, Caridad. 2005. Investigador titular del Instituto de Fruticultura Tropical (IIFT). MINAG, Cuba.
2 Cueto, J. (2005). Director General del Instituto de Fruticultura Tropical. MINAG, Cuba.

