










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. v.22 n.1 La Habana ene.-abr. 2007
Trabajo original
ANTAGONISMO DE CEPAS DE Pseudomonas fluorescens y Burkholderia cepacia CONTRA HONGOS FITOPATÓGENOS DEL ARROZ Y EL MAÍZ
ANTAGONIST ACTIVITY OF Pseudomonas fluorescens and Burkholderia cepacia STRAINS AGAINST RICE AND MAIZE PHYTOPATHOGENIC FUNGI
I. Trujillo, Acela Díaz, Annia Hernández y Mayra Heydrich
Facultad de Biología. Universidad de La Habana. Calle 25, #455, e/I y J. Plaza. Código Postal 10400. Ciudad de La Habana, Cuba. Correos electrónicos: acela@fbio.uh.cu, mayra@fbio.uh.cu
RESUMEN
Las bacterias promotoras del crecimiento vegetal (PGPB) influyen de forma directa o indirecta en el desarrollo de los cultivos. Este trabajo tiene como objetivos determinar la actividad antagónica y el efecto inhibitorio de los metabolitos activos producidos por rizobacterias ante fitopatógenos que afectan a los cultivos del maíz y el arroz. Para ello se desarrollaron bioensayos in vitro en el medio King B. Se utilizaron cepas de referencia y autóctonas de Pseudomonas fluorescens y Burkholderia cepacia, previamente aisladas de la rizosfera del maíz variedad Francisco mejorado, para determinar sus potencialidades como antagonistas de los patógenos fúngicos Fusarium oxysporum, Alternaria alternata, Curvularia sp. y Fusarium sp. Los resultados mostraron que la rizobacterias estudiadas tienen actividad antagónica y sus metabolitos efecto inhibitorio ante los hongos fitopatógenos enfrentados, destacándose con el mejor comportamiento las cepas de P. fluorescens MPf2, MPf3, J-143 y B. cepacia MPp8. Se observó un mayor efecto de las rizobacterias frente Curvularia sp. y A. alternata. Estos resultados poseen valor práctico, pues se selecciona un conjunto de cepas potencialmente eficientes en el control biológico de patógenos que podrían ser utilizadas en la agricultura en beneficio de los cultivos del arroz y el maíz.
Palabras clave: Pseudomonas fluorescens; Burkholderia cepacia; antagonistas; fitopatógenos.
ABSTRACT
Plant Growth Promoting Bacteria (PGPB) has been proved to exert a beneficial effect on crop yield, directly or indirectly. This work was aimed to determine the antagonistic activity and inhibitory effect exerted on different plant pathogens by rhizobacteria. A number of bioassays was carried in vitro in King B medium. Previously isolated bacterial strains of Pseudomonas fluorescens and Burkholderia cepacia, from corn plants var. Francisco mejorado, were assayed to determine their potential as antagonists of the phytopathogenic fungi Fusarium oxysporum, Alternaria alternata, Curvularia sp. and Fusarium sp. The results showed the capability of inhibiting the pathogen growth by the rhizobacterial strains, proving the best behaviour the strains P. fluorescens MPf2, MPf3, J-143 and B. cepacia MPp8. It was shown that there was a greater inhibitory effect on the fungi strains Curvularia sp. A1 and A. alternata M2. Considering these results, the bacterial strains were competent for pathogen biocontrol, and could be used in agriculture as valuable growth promoting agents in maize and rice crops.
Key words: Pseudomonas fluorescens; Burkholderia cepacia; antagonists; phytopathogens.
INTRODUCCIÓN
La aplicación de bacterias promotoras del crecimiento vegetal (PGPB) es una alternativa viable tanto en países con una agricultura subdesarrollada, que carecen de fertilizantes, como en los que practican la agricultura moderna, ya que permitiría reducir la cantidad de fertilizantes y plaguicidas químicos que contaminan el medio ambiente y deterioran el suelo, despojándolo de la materia orgánica y erosionándolo. Dentro de esta clasificación se agrupan múltiples géneros bacterianos, como Pseudomonas, Bacillus, Azotobacter, Acetobacter, Azospirillum, Burkholderia, Herbaspirillum y Klebsiella (9,18).
Las PGPB pueden promover el crecimiento por vías directas o indirectas, cuyos elementos específicos no han sido debidamente caracterizados. Los efectos directos pueden evidenciarse en ausencia de otros microorganismos, es decir, la planta solo interactúa con el microorganismo en estudio; mientras que los mecanismos indirectos se pueden observar en la interacción del microorganismo de interés con un fitopatógeno, mediante la cual se reducen los efectos dañinos en el vegetal (10,15).
Entre los mecanismos de control biológico mediados por rizobacterias ampliamente reconocidos se encuentran: la competencia por un nicho ecológico o sustrato, la síntesis de compuestos inhibitorios como sideróforos, antibióticos, enzimas líticas y detoxificadoras (1), así como la inducción de resistencia sistémica en la planta (7,11).
Las bacterias de los géneros Pseudomonas y Burkholderia, son de los grupos más estudiados porque tienen la capacidad de producir reguladores del crecimiento vegetal y otros metabolitos con efecto antagónico y represivo del crecimiento de patógenos en la rizosfera (6), por lo que resulta de gran importancia la caracterización y selección de cepas autóctonas de estos géneros, los cuales muestran efecto antagónico ante patógenos que atacan a cultivos de importancia económica, de forma tal que se puedan aprovechar sus potencialidades como agentes de control biológico.
En este sentido, el objetivo de este trabajo es determinar la actividad antagónica y el efecto inhibitorio de los metabolitos producidos por las cepas estudiadas ante hongos fitopatógenos de los cultivos del maíz y el arroz.
MATERIALES Y MÉTODOS
Cultivos microbianos: Se utilizaron nueve cepas autóctonas de las especies Pseudomonas fluorescens (MPf2, MPf3, MPf4, MPf5, MPf6 y MPf19) y Burkholderia cepacia (MPp8, MPp16, MPp20), previamente aisladas de la rizosfera del cultivo del maíz variedad Francisco mejorado en el año 2002 (5). Como controles fueron utilizadas las cepas de referencia de B. cepacia 0057 y P. fluorescens J-143, pertenecientes a la Colección de Cultivos de la Facultad de Biología de la Universidad de La Habana. Todas las cepas se encontraban conservadas en Caldo LB (DUCHEFA BIOCHEME) con glicerol al 40%, a -200C.
Se utilizaron cepas de los hongos fitopatógenos Curvularia sp. A1, Fusarium sp. A2, Fusarium oxysporum M1 y Alternaria alternata M2, todas gentilmente donadas por el Instituto de Investigaciones de Sanidad Vegetal (INISAV) y conservadas en tubos de cultivo con medio Papa Dextrosa Agar (PDA) (DIFCO), a 4oC.
Determinación del efecto antagónico de las rizobacterias ante fitopatógenos de los cultivos del maíz y el arroz: Se desarrollaron bioensayos in vitro para determinar el efecto antagónico de las cepas bacterianas ante cepas de hongos fitopatógenos de los cultivos del arroz (Curvularia sp. A1, Fusarium sp. A2) y el maíz (Fusarium oxysporum M1, Alternaria alternata M2). En todos los casos se siguió la metodología descrita por Bashan et al. (3).
Para este estudio fue utilizado el medio King B, compuesto por: 8.7 mL de glicerol, 1.5 g de peptona, 1.5 g de K2HPO4 y 1.5 g de MgSO4 (todo por L de H2O mili Q), teniendo en cuenta resultados precedentes que Demostraban que en este medio de cultivo ambos grupos microbianos mostraban un buen crecimiento (15) y que el mismo favorece la producción de metabolitos secundarios por parte de las bacterias utilizadas como antagonistas (12).
Para ello los hongos se sembraron previamente en el medio de cultivo seleccionado y se incubaron a 30oC±2 oC durante 7 días. Posteriormente, se realizaron perforaciones en las placas previamente sembradas con el hongo, en la parte central de la colonia, utilizando un obturador con diámetro de 5 mm. Los parches fueron colocados en el centro de las placas sembradas con la bacteria, utilizándose un parche para cada placa. Se sembraron ambos microorganismos al mismo tiempo y se realizó la incubación a 30oC durante 7 días. El efecto antagónico de las bacterias fue determinado a través de mediciones del diámetro de crecimiento del hongo (mm) en comparación con un tratamiento control donde no se aplicó el antagonista bacteriano. Las placas Petri utilizadas tenían un diámetro de 90mm.
Determinación del efecto inhibitorio de metabolitos producidos por las rizobacterias ante fitopatógenos de los cultivos del maíz y el arroz: Para determinar el efecto inhibitorio de los metabolitos producidos por las cepas se realizaron bioensayos in vitro (3). Para ello primeramente las placas Petri con el medio de cultivo King B se sembraron con las diferentes cepas de rizobacterias por diseminación con espátula de Drigalsky y se incubaron durante 48 horas a 30oC±2oC y 37oC±2oC, en dependencia de la especie bacteriana. Una vez crecidas las bacterias, las placas se sometieron a vapores de cloroformo durante 30 minutos (5). Posteriormente fueron cola dos discos de 5 mm de diámetro que contenían los hongos patógenos en el centro de las placas libres de células.
Los discos de hongos fueron previamente obtenidos como se describe en el acápite anterior. El porcentaje de inhibición se determinó a través del diámetro de crecimiento del hongo (mm) en comparación con el control.
En ambos casos se estableció un diseño Completamente Aleatorizado estableciéndose 10 réplicas por tratamiento. Los bioensayos fueron repetidos 3 veces cada uno.
Análisis biométricos: Como punto de partida a los porcentajes de inhibición se les realizó la prueba de normalidad de Shapiro y Wilk, haciendo uso de la versión simplificada (13), y la prueba de homogeneidad de varianza, a partir de las cuales se decidió realizar análisis no paramétricos.
Los datos fueron analizados utilizando la prueba no paramétrica de comparación múltiple de Kruskal Wallis seguido de la prueba de comparación múltiple de medias no paramétrica de Student-Newman-Keuls (SNK), con el paquete estadístico STATISTIC versión 6.0.
RESULTADOS
Efecto antagónico de las cepas bacterianas en estudio
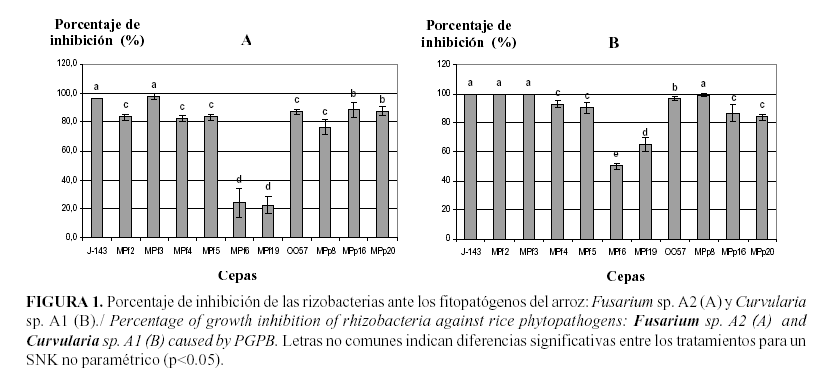
Los resultados mostraron que las cepas de rizobacterias ejercen efecto antagónico ante los patógenos Estudiados. Al establecerse la interacción entre los antagonistas bacterianos y las cepas de Fusarium sp. A2 y Curvularia sp. A1 se observaron diferentes niveles en los porcentajes de inhibición.
Al enfrentar las rizobacterias a la cepa de Fusarium sp. A2, las cepas de P. fluorescens MPf3 y J-143 se destacaron por presentar los mayores porcentajes de inhibición, mostrando diferencias significativas con el resto de los tratamientos (Fig. 1). Asimismo todas las cepas estudiadas mostraron efecto antagónico frente a la cepa de Curvularia sp. A1. En este caso, las cepas P. fluorescens MPf2, MPf3 y J-143 se destacaron por lograr una inhibición total del crecimiento del hongo, no mostrando diferencias significativas con la cepa MPp8 (Fig. 1).
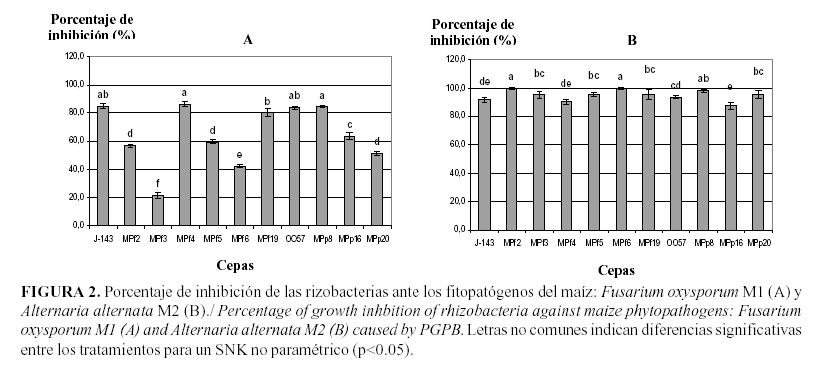
Por otro lado, al analizar el efecto antagónico de las cepas bacterianas en estudio ante los hongos fitopatógenos F. oxysporum M1 y A. alternata M2, también se observaron altos porcentajes de inhibición del crecimiento.
Con relación a F. oxysporum M1, en la Figura 2 se muestra que las cepas que presentaron los mayores índices de inhibición fueron las de P. fluorescens J-143, MPf4, B. cepacia 0057 y MPp8.
Al interactuar las rizobacterias con A. alternata M2, se observaron altos índices de inhibición del crecimiento del hongo, al compararse los tratamientos con el control (Figura 2). Las cepas más destacadas fueron las de P. fluorescens MPf2, MPf6 y B. cepacia MPp8, mostrando porcentajes de inhibición superiores al 97%.
Efecto inhibitorio de los metabolitos producidos por las rizobacterias en estudio
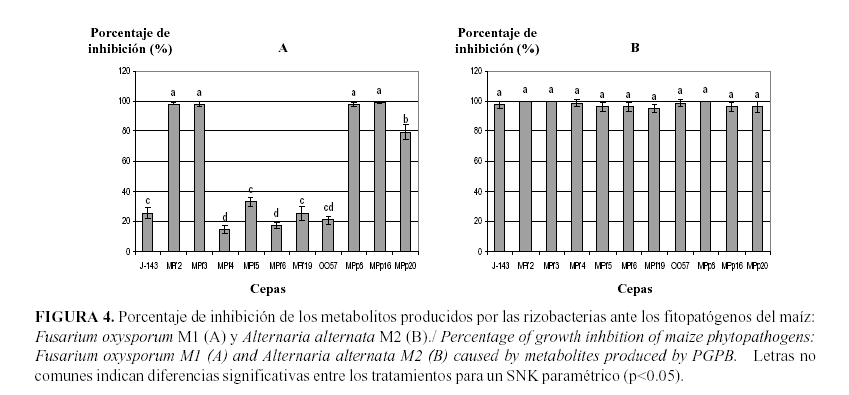
Resultados similares a los anteriores fueron encontrados al enfrentar los metabolitos activos producidos por las rizobacterias a los hongos fitopatógenos en estudio, lo que indica el efecto inhibitorio de los mismos.
Los metabolitos producidos por las cepas MPf2, MPf3 y MPp8 lograron inhibir totalmente el crecimiento del hongo Fusarium sp. A2, mostrando diferencias significativas con el resto de los tratamientos (Fig. 3). Los metabolitos que menor inhibición del crecimiento fúngico evidenciaron fueron los producidos por P. fluorescens MPf5 y MPf19, con diferencias significativas entre sí.
Los metabolitos producidos por las cepas bacterianas también inhibieron el crecimiento de Curvularia sp. A1 (Figura 3), al igual que las cepas MPf2, MPf3 y MPp8 inhibieron un 100% del crecimiento del hongo. En este caso las cepas que menor efecto inhibitorio mostraron fueron las de B. cepacia 0057 y P. fluorescens MPf19.
Al enfrentar los metabolitos producidos por las rizobacterias a los hongos fitopatógenos A. alternata M2 y F. oxysporum M1 también observaron diferentes niveles de inhibición. Las cepas que mostraron mayores porcentajes de inhibición frente a Fusarium oxysporum M1 fueron MPf2, MPf3, MPp8 y MPp16.
se mostró un alto nivel de inhibición al enfrentarse los metabolitos producidos por las cepas al fitopatógeno A. alternata M2 (Fig. 4). En todos los tratamientos se encontraron porcentajes de inhibición del crecimiento fúngico con valores superiores al 94%, sin diferencias significativas entre los tratamientos. De este modo se corrobora la sensibilidad del hongo ante los productos del metabolismo de las rizobacterias en estudio.
DISCUSIÓN
En este trabajo se demostró que las cepas de rizobacterias utilizadas tienen efecto antagónico e inhibitorio ante los hongos fitopatógenos Curvularia sp. A1, Fusarium sp. A2, Fusarium oxysporum M1 y Alternaria alternata M2, lo que corrobora resultados obtenidos por otros autores que han señalado el efecto de las bacterias asociadas a las plantas en el control biológico de patógenos (6, 19).
Las cepas de P. fluorescens y B. cepacia estudiadas y sus metabolitos activos ejercen efecto antagónico e inhibitorio ante los hongos Fusarium sp. A2, Curvularia sp. A1, F. oxysporum M1 y A. alternata M2, destacándose por el mejor comportamiento las cepas MPf2, MPf3, J-143 y MPp8.
Se observó un mayor efecto de las rizobacterias antagonistas estudiadas frente a los hongos fitopatógenos Curvularia sp. A1 y A. alternata M2.
Las respuestas obtenidas podrían estar relacionadas con la producción de metabolitos de naturaleza antibiótica. De hecho, Velázquez et al. (17) demostraron que la cepa de Burkholderia cepacia 0057, utilizada en este estudio, producía alcaloides quinolisidínicos de naturaleza antibiótica. También se ha señalado la capacidad que tienen las cepas de Pseudomonas de suprimir patógenos presentes en la rizosfera, a través de la producción de 2,4 dimetyl-phloroglucinol y pirrolnitrina (16).
En este caso, también podrían haber influido los sideróforos, si se tiene en cuenta que el medio de cultivo utilizado para el desarrollo de los bioensayos posee bajo contenido de hierro, lo que favorece la producción de este tipo de metabolito. En este sentido, diferentes autores (6, 12), señalaron que cepas autóctonas cubanas pertenecientes a las especies en estudio al crecer en medios de cultivo con concentraciones limitadas de hierro, tales como el King B y el Sirope fructosa (Sp.), producían sideróforos a partir de las 10 horas de crecimiento de la bacteria.
Asimismo, se demostró que cepas de las especies B. cepacia y P. fluorescens producían diferentes tipos de sideróforos (2, 6, 8), los que podrían estar involucrados en el antagonismo microbiano y en la resistencia sistémica inducida (RSI) en diferentes sistemas planta-patógeno.
Nuestros resultados evidencian que los metabolitos activos de origen bacteriano desempeñan una función vital en las interacciones microbianas. Sin embargo, se ha señalado que los antagonistas microbianos no poseen un único modo de acción, y que no es fácil determinar con precisión los mecanismos que intervienen en las interacciones entre los antagonistas y los fitopatógenos (4, 14).
Estos resultados poseen valor práctico pues se evalúa un conjunto de cepas con efecto antagónico ante patógenos que afectan a los cultivos del arroz y el maíz, las que podrían ser utilizadas en la biotecnología agrícola con vistas a obtener biopreparados en beneficio de cultivos de interés económico, una vez que se estudie su efecto en interacción con la planta.
REFERENCIAS
1. Bais HP, Park SW, Weir TL, Callaway RM, Vivanco JM. How plants communicate using the underground information superhighway. Trends Plant Sci. 2004;(9):26-32.
2. Barelman I, Meyer JM, Taraz K, Budzikiewicz H. Cepaciachelin, a new catecholate siderophore from Burkholderia (Pseudomonas) cepacia. Z Natarfosch. 1996;(51):627-630.
3. Bashan Y, Holguín G, Ferrera R. Interacciones entre plantas y microorganismos benéficos. Terra. 1996;14(2):159-192.
4. Fernández-Larrea Orieta. Microorganismos antagonistas para el control fitosanitario. Manejo Integrado de Plagas. (Costa Rica). 2001;(62):96-100.
5. Hernández Annia. Obtención de un nuevo producto bacteriano a partir de rizobacterias asociadas al cultivo del maíz (Zea mayz L.). Tesis en opción al Grado Científico de Doctor en Ciencias Biológicas; 2002.
6. Hernández Annia, Rives N, Caballero A, Hernández AN, Heydrich M. Caracterización de rizobacterias asociados al cultivo del maíz en la producción de metabolitos del tipo AIA, sideroforos y ácido salicilico. Rev Col Biotec. 2004;6(1):6-13.
7. Matiru VN, Dakora F. Potential use of rhizobial bacteria as promoters of plant growth for increased yield in landraces of African cereal crops. African J Biotech. 2004;(3):1-7.
8. Meyer JM, Trân Van V, Stinzi A, Berge O, Winkelmann G. Ornibactin production and transport properties in strains of Burkholderia vietnamiensis and Burkholderia cepacia (formally Pseudomonas cepacia). BioMetals. 1995;309-317.
9. Nelson LM. Plant growth promoting rhizobacteria (PGPR): Prospects for new inoculants. Online. Crop Management, Plant Management Network; 2004.
10.Ping L, Boland W. Signals from the underground: bacterial volatiles promote growth in Arabidopsis. Trends Plant Sci.2004; 9(6):262-266.
11.Ryu CM, Murphy JF, Mysore KS, Kloepper JW. Plant growth-promoting rhizobacterial systemically protect Arabidopsis thaliana against Cucumber mosaic virus by a salicylic acid and NPR1-independent and jasmonic acid-dependent signaling pathway. The Plant J. 2004;(39):381-392.
12.Sánchez L, Ortiz S, Hernández A. Evaluation of two culture media for producing siderophore from Burkholderia cepacia through stirred-tank fermentation. Rev Protección Veg. 2004;19(2):97-101.
13.Shapiro SS, Francia RS. An approximate Analysis of Variance Test for Normality. J American Statistical Association. 1972;(67):215-216.
14.Trujillo ID, Hernández A, Díaz A, Heydrich M. Selección de cepas de P. fluorescens y B. cepacia para el biocontrol de patógenos asociados al cultivo del arroz. En: Actas del III Encuentro Internacional del arroz. La Habana. Cuba. ISBN 959-7164-91-4; 2005.
15.Unno Y, Okubo K, Wasaki J, Shinano T, Osaki M. Plant growth promotion abilities and microscale bacterial dynamics in the rhizosphere of Lupin analysed by phytate utilization ability. Env Microb. 2005;7(3):396-404.
16.Validov S, Mavrodi O, De La Fuente L, Boronin A, Weller D, Thomashow L, Mavrodi D. Antagonistic activity among 2,4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. FEMS Microb. Letters. 2005;(242):249-256.
17.Velázquez M, Hernández A, Heydrich M, Hernández AN. Estudio de la interacción maíz-Burkholderia cepacia. Rev Lat Microb. 1999;(41):17-23.
18.Vessey JK. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil. 2003;(255):571-586.
19.Zehnder GW, Murphy JF, Sikora EJ, Kloepper JW. Application of rhizobacteria for induced resistance. Eur J Plant Path. 2001;107(1):39-50.
(Recibido 20-3-2006; Aceptado 14-9-2006)


{kind=link}
{kind=link}
{kind=link}
{kind=link}