










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752
Rev. Protección Veg. vol.28 no.1 La Habana ene.-abr. 2013
ORIGINAL ARTICLE
Characterization of Citrus tristeza virus isolates from Colombia
Caracterización de aislamientos de Citrus tristeza virus de Colombia
Juan MoralesI*, Orlando AcostaII, Pablo TamayoIII, José PeñarandaII,†
IUniversidad Nacional de Colombia sede Medellín, Departamento de Ciencias Agronómicas. Calle 59A No 63-20,
Núcleo el Volador, Bloque 11. Medellín-Colombia. Tel: 0057 4 4309099, fax: 0057 4 4309118. E-mail: jgmoraleso@unal.edu.co.
IIUniversidad Nacional de Colombia sede Bogotá, Colombia. Departamento de Ciencias Fisiológicas.
IIICorporación Colombiana de Investigación Agropecuaria, CORPOICA. Regional 4 "La Selva", Rionegro-Antioquia, Colombia.
ABSTRACT
Isolates of Citrus tristeza virus around the world differ widely in their biological properties. These properties may depend on strain mixtures within individual trees. Monoclonal Antibody 13 (MCA13) is an antibody that detects almost all the severe strains causing Quick Decline and Stem Pitting CTV syndromes. The objectives of this work were: I) to determine the percentage of CTV isolates reactive to the MCA13 antibody in each Colombian region evaluated and II) to analyze the genetic diversity of isolates from these regions. In almost all the regions evaluated, CTV MCA13 reactive isolates were identified; however, only one was found in Mompox area. The coat protein gene was amplified by reverse transcription-polymerase chain reaction (RT-PCR) and analyzed by single strain conformational polymorphisms (SSCP) patterns. In this work, at least twelve different dsRNA electrophoretic profiles and seven SSCP different electrophoretic patterns were identified. At least two different isolates were shown to be a mixture of strains. Non reactive strains to MAC13 from the Mompox region could be the best candidates for a Tristeza disease cross-protection program.
Key words: virus cross-protection, citrus tristeza disease, Citrus spp., RT-PCR, SSCP.
RESUMEN
Los aislamientos de Citrus tristeza virus de diferentes regiones del mundo difieren en sus propiedades biológicas. Esas propiedades pueden depender de mezclas de cepas en el interior de árboles individuales. El anticuerpo monoclonal MCA13 detecta casi todas las cepas severas que causan los síndromes de declinamiento rápido y acanalamiento de tallos. Este trabajo tuvo como objetivos: I) determinar el porcentaje de aislamientos de CTV reactivos al anticuerpo monoclonal MCA13, en cada región Colombiana evaluada y II) analizar la diversidad genética de los aislamientos provenientes de esas regiones. En este trabajo identificamos aislamientos de CTV reactivos al MCA13 en casi todas las regiones de Colombia; sin embargo en Mompox, solamente se encontró uno. La proteína de la cápside fue amplificada por transcripción reversa seguida de la reacción en cadena de la polimerasa y los patrones de polimorfismo conformacional de una sola hebra (SSCP) fueron analizados. Se identificaron al menos 12 perfiles electroforéticos de RNA de doble hebra y siete perfiles SSCP. Se encontró que al menos dos aislamientos diferentes están compuestos de mezclas de cepas. Las cepas no reactivas al MCA13 podrían ser buenas candidatas para un programa de protección cruzada.Palabras clave: protección cruzada en virus, enfermedad de la tristeza de los cítricos, Citrus spp., RT-PCR, SSCP.
INTRODUCTION
Citrus tristeza virus (CTV) is a member of the genus Closterovirus, family Closteroviridae of (+) sense, single-stranded RNA plant viruses. CTV is a phloem virus associated with a host range primarily in the genus Citrus (1). Virus strains induce various economically important diseases worldwide and are a significant threat to citrus production (1). CTV produces several symptoms depending on the host and the bud and scion combination on graft (1). Main symptoms are quick decline, seedling yellowing and stem pitting (2).
Virus particles spread through aphid transmission and vegetative propagation (1); Toxoptera citricida is the most efficient vector within the aphids and is spreading around the world from South Africa to South America reaching Florida-USA, Madeira Island, and Mainland Portugal (3). The complete viral genome sequence is available (1), allowing for extensive molecular analysis, detection and strain characterization in several countries around the world (1).
CTV detection is possible either by polyclonal or monoclonal antibodies (1). The monoclonal antibody MCA13 can detect 95% of severe strains causing quick decline and stem pitting in sweet orange grafted onto sour orange seedlings (4). In a previous survey made in Colombia, a CTV incidence of 95% was quantified using polyclonal and monoclonal antibodies by the ELISA test (Enzyme linked immunoabsorbent assay) (5, 6), excluding the Mompox-Bolívar area, where the incidence was 70%. According to the detection by the monoclonal antibody MCA13, CTV strains detected in almost all Colombia were severe excluding Mompox-Bolívar, where MCA13-severe strains were not found (5).
From the point of view of disease management, genetic engineering and mild strain-mediated cross- protection are the only known means for the disease control (1, 7). Selection for potential effective cross-protection mild strains is an expensive and time consuming process (7, 8). Molecular tools may help to select mild strains in a short period at a low cost (1, 6, 9, 10). Understanding the variation at the molecular level is of crucial importance to design fast and maybe cheap means of strain differentiation and selection.
In this research, we measured CTV incidence in several Colombian Citrus-growing areas and determined the double stranded ribonucleic acid (dsRNA) and single strand conformational polymorphism (SSCP) patterns of the CTV p25 gene from Colombian isolates.
The objectives of this work were: I) to determine the incidence of the CTV isolates reactive and not reactive to the MCA13 antibody en each Colombian region evaluated and II) to analyze the CTV dsRNA patterns and the coat protein gene p25 diversity by single strand conformational polymorphism (SSCP) of isolates from these regions.
MATERIALS AND METHODS
Sampling
Ninety-five isolates were collected in the main citrus producing areas in Colombia in the departments of Antioquia (northwest), the region of Mompox-Bolívar (comprising the departments of Bolivar and Magdalena, north) and Meta (east). A zig-zag sampling strategy was followed inside each field. Department, farm, host (scion-rootstock), age, and visual symptoms such as healthy or deficient development, vein clearing, quick decline and stem pitting, were recorded. Green bark tissue of tree young shoots was removed and ground in liquid nitrogen until a fine powder was obtained, it was stored at -60°C for further use (11).
For virus detection, 4ml of extraction buffer were added to 1g of sample powder, carefully mixed and incubated for 15 min at room temperature. The mixture was centrifuged at 5000 g for 10 min and the supernatant transferred to a clean tube for being used in the serological tests.
Virus Detection
The virus was detected using the Enzyme Linked Immunosorbent Assay (ELISA), Agdia Kit test (5). To detect severe strains, the MCA13 monoclonal antibody was used (4) in an I-DAS-ELISA test; this antibody detects almost all the severe strains and has been widely used around the world (4, 5). The measurements were carried out in an ELISA reader at 405 nm wavelength. Positive results were those samples showing an optical density (O.D./405nm) twice or more times higher than that of the mean value of the negative control (5, 12).
DsRNA electrophoretic pattern
Seven grams of bark of infected tissue were collected in a centrifuge tube, and 12 ml of STE double-strength buffer (2X STE= 0,2M NaCl; 0,1M Tris; 0,002M EDTA, pH 6.8) were added together with 15ml of STE-saturated phenol and 1,5 ml of 10% SDS. The mixture was shaken for 30min at room temperature and then centrifuged at 8000g for 20min, the aqueous phase recovered and adjusted to 16,5% ethanol.
The solution was percolated through a CF-11 cellulose powder column in the presence of STE buffered with 16,5% ethanol (11). The column was washed with 100ml of STE-buffered with 16,5% ethanol, and the dsRNA contained in the washed column was eluted with 15 ml of ethanol-free STE. The dsRNA in the collected eluate was precipitated with 2,5 volumes of 95% ethanol and 1/20 volumes of sodium acetate 3M pH 5.5 and stored at -20°C for one hour or indefinitely.
Precipitated dsRNA was collected by centrifugation at 8000g for 30min and resuspended in 1 ml of STE, transferred to an eppendorf tube and ethanol and sodium acetate precipitated as described, centrifuged at 3000g for 30min and resuspended in TAE buffer (40mM Tris, 20mM Sodium acetate, 1mM EDTA) (11, 13). The samples were run on polyacrylamide gel electrophoresis (PAGE) (6%) for 3h at 100v, and silver nitrate- stained as described before (14).
cDNA synthesis and cloning.
DsRNA was used as a template for reverse transcription-polymerase chain reaction (RT-PCR). To denature the template, a reaction tube containing: 5,0 ml dsRNA template, 500ng of each of two primers CN150 (5'ATA TAT TTA CTC TAG ATC TAC CAT GGA CGA CGA AAC AAA) and CN151 (5'GAA TCG GAA CGC GAA TTC TCA ACG TGT GTT AAA TTT CC) (15); 22,5 ml of HPLC water and 25ml mineral oil overlain, were heated at 98°C for 5min and chilled on ice. To the denatured template tube reaction, 10ml of RT 5X GibcoÒ Buffer, 5ml 0,1M ditiothreitol; 0,5ml Rnasin (40U/ml); 2,5ml DNT`s mix (10mM); 2,0ml M-V (200U/ml) were added in a final volume reaction of 50ml and incubated at 37°C for 60min for retrotranscription.
The cDNA was amplified by the polimerase chain reaction as follows: 2,5 ml of PCR buffer 10X GibcoÒ; 2,5ml of cDNA (50ng); 1,25ml MgCl2; 1ml DNTP's (10mM), 300ng of each of two primers CN150 and CN 151; 0,625 U Taq DNA polymerase (5U/ml) and 16,875ml of HPLC water in a final volume reaction of 25ml. Then the following thermocycling conditions were used: 94°C for 1min, 55°C for 1min, 72°C for 2min (35 cycles) and a final extension at 72°C for 10min. After amplification, the PCR products were separated from the mineral oil overlain by chloroform (1,5 volumes) extraction.
ThePCR products were analyzed on agarose gel electrophoresis (1% agarose gel in TAE buffer at 100v for 40 minutes) stained with ethidium bromide (0,5mg/) and visualized under UV light at 300 nm (15).
PCR amplified fragments were cloned into pGEM3zf+ plasmid by preparing a T-overhang PGEM3zf+ plasmid. 10mg of Sma I-digested vector were incubated in PCR buffer (GibcoÒ) with 2mM dTTP and 5U Taq DNA polymerase at 72°C for 2h. After phenol-chloroform extraction and ethanol precipitation, the pellet containing the T-overhang vector was re-suspended in sterile distilled water and ligated for 16h at room temperature in 50ml reaction containing 2U of T4 ligase and ligation buffer as recommended by the manufacturer (GibcoÒ). Escherichia coli cells were transformed with the plasmid-insert construct and colonies containing plasmid-insert were selected with X-gal, following the standard procedures described by Sambrook et al. (16). The insert was verified by agarose (1%) gel electrophoresis as described.
Single strand conformational polymorphism, (SSCP)
SSCP analysis was performed directly for the PCR products of the coat protein cDNA gene (cp) of Colombian isolates following the procedure described by Rubio et al. (9) modified by Morales (17). One microliter of PCR product was mixed with 9ml of denaturing solution (950ml/L formamide, 20mM EDTA, 500 mg/L bromophenol blue), heated for 10min at 99°C and chilled on ice. Denatured DNA was electrophoresed in a non-denaturing 80g/L polyacrylamide gel, using TBE (Tris-borate 89mM, 2mM EDTA, pH 8.0) as electrophoresis buffer and a constant voltage (200v) for 2,5h at room temperature (17). The gels were silver nitrate- stained following the procedures described by Rubio et al. (9), modified by Morales (17).
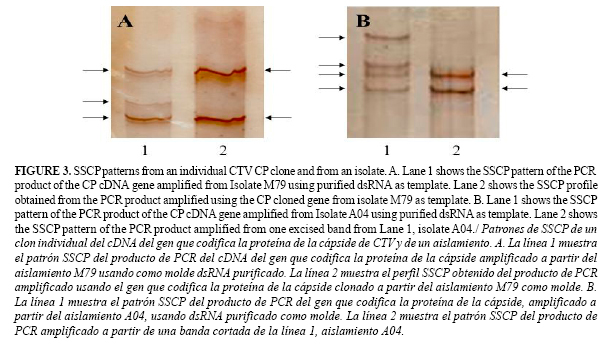
To determine if a band pattern from one isolate corresponded to a single or a mixture of sequences, the cp gene from isolates M79 (MCA13 non-reactive) and A01 (MCA13 reactive) were cloned and the individual clones tested. PCR products were separated by PAGE, and the band was excised from the polyacrilamide gel and cloned in plasmid pGEM3zf+, amplified by PCR and electrophoresed as described before. Alternatively, a band was excised and directly amplified by PCR and electrophoresed for SSCP pattern analysis as described (17).
Data evaluation and statistical analysis
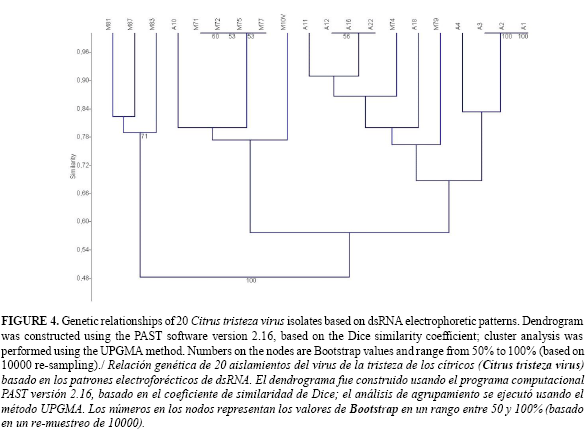
A binary character matrix was constructed where the presence of a band in a particular position was recorded as 1 and absence as 0 (18, 19). The PAST software version 2.16 (19), was used to calculate the DICE similarity coefficient between all samples and the cophenetic correlation coefficient. A cluster analysis was performed using the unweighted pair group method with arithmetic mean (UPGMA) based on the Dice index (19). Bootstrap values (based on 10000 re-samplings) were used to estimate the reliability of the clustering pattern.
RESULTS AND DISCUSSION
CTV incidence
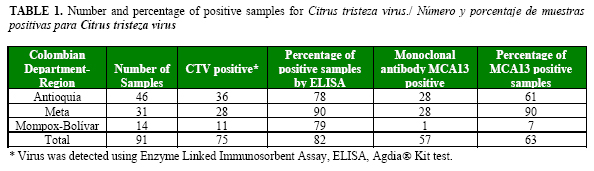
CTV was detected in all the regions surveyed. From 91 samples evaluated by DAS-ELISA test, 75 (82%) were positive (Table 1). Previous studies showed similar results, indicating that this disease was well established in Colombian citrus crops (5). The highest incidence was found in Meta, followed by the Mompox-Bolívar region and the department of Antioquia. More important, from the point of view of disease losses and management, was the distribution of mild and severe strains. In Meta, most isolates were positive to the severe strain detecting with antibody MCA13, followed by Antioquia and Mompox-Bolívar (1 sample) (Table 1).
Most isolates collected in Mompox-Bolívar were mild, confirming previous results and pointing out the importance of this region as a source of potential mild strains for cross protection (5) (Table 1). In a previous survey, Peñaranda and collaborators (unpublished results) collected 18 seedlings pre-infected with MCA13-non-reactive strains in the region of Mompox-Bolívar and planted them in Meta, in a farm previously identified with 100% incidence of CTV and Toxoptera citricida as the most frequent vector (5). Non-infected seedlings were used as negative controls. In this study, samples from this experiment were collected and evaluated by using monoclonal and polyclonal antibodies as described in Materials and methods.
After six years, the trees pre-infected with mild strains were growing without external symptoms and all the controls died except one. The standing control tree showed external symptoms of quick decline. ELISA tests were positive for both monoclonal and polyclonal antibodies confirming infection of trees by a severe strain. All the trees pre-infected with mild strains from the region of Mompox were positive to MCA13 suggesting that they had acquired severe strains in the department of Meta by natural mechanisms of infection. Although all the samples were positive by DAS-ELISA using MCA13, the trees pre-infected with mild strains did not showed external symptoms at the time the samples were taken.
DsRNA electrophoretic pattern
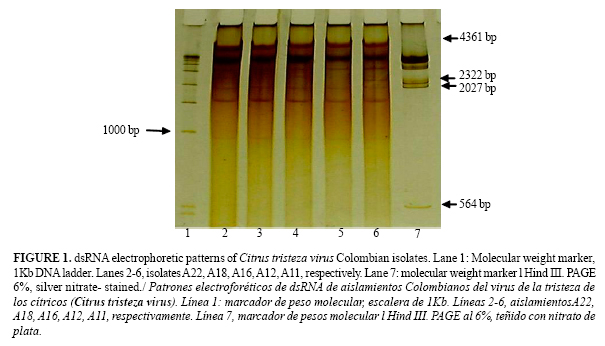
Twelve different dsRNA electrophoretic patterns were found indicating variability between isolates. All the isolates tested showed two characteristic dsRNA bands of 13,3 x 106 Daltons (Da) (replicative form of the viral genome) and 0,8x106 Da (replicative form of the coat protein gene p25) respectively (11, 13) (Figure 1).
Six different patterns were present in the isolates from Antioquia. A band of 0,5 x 106 Da was detected in all of them except one. Furthermore, all isolate having this band was MCA13-positive. In previous studies, the presence of this band (0,5 x 106 Da) was correlated with MCA13-reactive strains (4, 11) exhibiting seedling yellowing and stem pitting symptoms in grapefruit (11, 13). Seven different patterns were detected from the Mompox-Bolívar region and twelve in the isolates from the Meta region. In general, a more complex pattern was present in MCA13-reactive isolates than in the non-reactive ones (Figure 1).
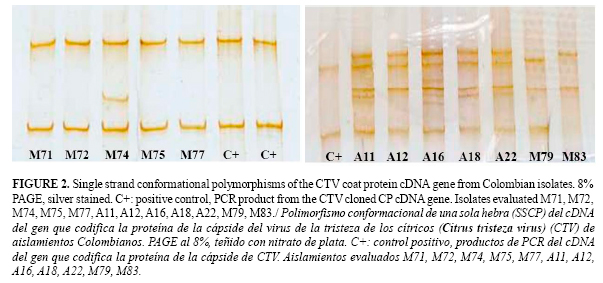
Single strand conformational polymorphism, (SSCP)
Using the SSCP method, a single base mutation could be detected (9) without sequencing saving money and time. Seven different SSCP patterns were identified in the isolates collected. The more complex patterns corresponded to the isolates from Antioquia (Figure 2). As for dsRNA results, MCA13-reactive strains in general showed more complex SSCP patterns than non MCA13-reactive strains (Figure 2). To identify a possible strain mixture, the CTV coat protein gene from isolates A01 (MCA13 reactive) and M79 (MCA13 non-reactive) were cloned and the SSCP patterns between an individual clone and the original isolate compared. Two bands in the clone SSCP pattern and three (M79) or four (A01) in the corresponding isolates were found (Figure 3).
A similar result was obtained when a band was excised from the polyacrylamide gel and the SSCP pattern compared with the corresponding isolate. These results indicated that some isolates were composed of a strain mixture and that Tristeza disease could involve a complex rather than an individual strain of CTV. It is probable that most isolates evaluated by SSCP and dsRNA were composed by several strains since most patterns were complex. Interestingly, in the Mompox-Bolívar region, most isolates were MCA13 non-reactive (Table 1) and exhibited only two bands in the SSCP pattern compared with the complex patterns from Antioquia and Meta (Figure 2).
All the citrus trees pre-infected with mild strains from the region of Mompox and planted in the Meta department were positive to MCA13, indicating that they could have had a mixture of mild (pre-infected) and severe (in-field acquired) strains. Furthermore, biological, biochemical and molecular evidences of infection of individual trees by several different strains of CVT had been reported before (20, 21, 24).
Tristeza disease may have components of several strains and strain interactions can be very complex (22). The CTV virion contains a monopartite positive-sense RNA genome (22). RNA viruses can evolve very fast but evidence of slow evolution of CTV has been found (23). The molecular analysis of isolates can be difficult for the presence of many strains in an individual tree. Sophisticated molecular techniques have been developed to detect and characterize RNA viruses, but most of them require expensive equipment and skilled scientists (20, 24, 25). Virus gene amplification by PCR followed by SSCP analysis discriminates precisely virus strains using a simple technique (6, 9). It is important to clone genes for future studies to avoid any interference caused by a mixture of strains.
The dendogram showed 13 divisions, three corresponded to groups with similar isolates (Figure 4). Isolates A1, A2 and A3 clustered together and were collected in Antioquia from lime cv. Tahiti; a second cluster with isolates A12, A16 and A22 collected in Antioquia from orange cv. Valencia; isolates in this group showed a complex SSCP pattern (Figures 2). A third group was integrated by isolates M71, M72, M75 and M77 collected in Mompox-Bolívar from orange cv. Margarita. This group exhibited a simple SSCP pattern (Figure 2). Isolates A4, A10, A11, A18, M10v, M74, M79, M81, M83 and M87, plotted at independent points suggesting diversity compared to themselves and to the other three groups (Figure 4). Most isolates plotting independently showed a complex SSCP pattern (Figure 2) indicating a possible strain mixture.
From Mompox region, we identified potential mild strains that could be evaluated against severe strains. These strains have been sequenced (5, 6) and characterized by other methods like polyclonal and monoclonal antibodies and dsRNA (5). In this study, SSCP patterns that can help for further strain differentiation are shown. At least for two isolates, the evidence indicated that they were composed of a mixture of different strains in an individual tree. The SSCP method is an economic and fast technique to test the strain mixture within an individual tree.
CTV is well established in all the Colombian citrus growing areas evaluated. The socially and economically acceptable disease management strategy in the short term is mild strain cross-protection; this type of control requires a simple procedure for the selection of potential mild strain candidates to overcome the long greenhouse period of evaluation before field tests. The first example for Colombia of a field assay of mild strain cross-protection was carried out in the Meta Departmet. Because of the favorable conditions in Meta fields for the development of Tristeza disease, these results are promising. Mild strains from Mompox-Bolívar are characterized at the biological and molecular level and constitute the best candidates to be tested in a program of cross-protection in a large scale in the near future.
CONCLUSIONS
CTV disease is established in Antioquia, Meta, and Mompox Colombian regions. Incidence of CTV is 84%; incidence of severe strains is 62% (MCA13-reactive). In two cases, evidence of several strains within one individual tree was found. The SSCP analysis is useful to determine the presence of a mixture of CTV strains in an individual tree. Mild strains from the Mompox-Bolívar region are the most promising strains for a cross-protection program in Colombian citrus fields.
ACKNOWLEDGMENTS
This work was supported by COLCIENCIAS, UNIVERSIDAD NACIONAL DE COLOMBIA and CORPOICA.
REFERENCES
1. Moreno P, Ambrós S, Albiach-Mart í M, Guerri J, Peña I. Citrus tristeza virus: a pathogen that changed the course of the citrus industry. Mol Plant Pathol. 2008;9(2):251-268.
2. Herrera-Isidrón L, Ochoa-Sánchez J, Rivera-Bustamante R, Martínez-Soriano J. Sequence diversity on four ORFs of Citrus tristeza virus correlates with pathogenicity. Virology Journal. 2009;6:116.
3. Roy A, Brlansky R. Population dynamics of a Florida Citrus tristeza virus isolate and aphid-transmitted subisolates: Identification of three genotypic groups and recombinants after aphid transmission. Phytopathology. 2009; 99:1297-1306.
4. Permar T, Garnsey S, Gumpf D, Lee R. A monoclonal antibody that discriminates strains of citrus tristeza virus. Phytopathology. 1990;80:224-228.
5. Acosta O, Alegria A, Guzmán M, Lee R, Niblett C, Peñaranda J. El virus de la Tristeza de los cítricos: una grave amenaza para la citricultura colombiana. Ed. Científica, Santa Fe de Bogotá, Colombia; 1994. 58p.
6. Oliveros-Garay OA, Martinez-Salazar N, Torres-Ruiz Y, Acosta O. CPm gene diversity in field isolates of Citrus tristeza virus from Colombia. Archives of Virology. 2009;154(12):1933-1937.
7. Soler N, Plomer M, Fagoaga C, Moreno P, Navarro L, et al. Transformation of Mexican lime with an intron-hairpin construct expressing untranslatable versions of the genes coding for the three silencing suppressors of Citrus tristeza virus confers complete resistance to the virus. Plant Biotechnology Journal. 2012;10:597-608.
8. Muller G, Costa S. Search for outstanding plants in Tristeza infected citrus orchards: the best approach to control the disease by preimmunization. Phytophylactica. 1987;19:187-191.
9. Rubio L, Ayllon M, Guerri J, Pappu H, Niblett C, Moreno P. Differentiation of citrus tristeza closterovirus (CTV) isolates by single-strand conformation polymorphism analysis of the coat protein gene. Annals of applied biology. 1996;129:479-489.
10.Costa A, Nunes W, Zanutto C, Müller G. Stability of Citrus tristeza virus protective isolates in field conditions. Pesq Agropec Bras. 2010;45(7):693-700.
11.Jarupat T, Lee J, Dodds J. Additional factors Affecting dsRNA Analysis of Citrus Tristeza Virus. Proceedings of the Eleventh IOCV Conference. 1991; pp 137-145.
12.Garnsey SM, Cambra M. Enzyme-linked immunosorbent assay (ELISA) for citrus pathogens. En: Roistacher CN (Ed). Graft-transmissible diseases of citrus. Handbook for detection and diagnosis. FAO, Rome. 1991; pp 193-216.
13.Dodds J, Jarupat T, Roistacher C, Lee R. Detection of strain double-stranded RNAs in citrus species infected with citrus tristeza virus: a review. Phytophylactica. 1987;19:131-137.
14.Valverde R. Analysis of double-Stranded RNA for Plant Virus Diagnosis. Plant Disease. 1990;74(3):255-258.
15.Pappu H, Pappu S, Niblett C, Lee R, Civerolo E. Comparative sequence analysis of the coat proteins of biologically distinct citrus tristeza closterovirus isolates. Virus Genes. 1993;7(3):225-264.
16.Sambrook J, Fritsch E, Maniatis T. Molecular Cloning: A Laboratory Manual, 2nd. ed. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press. 1989.
17.Morales J. Caracterización de aislamientos colombianos del virus de la tristeza de los cítricos, mediante análisis serológico de la proteína de la cápside y molecular de su gen. [Tesis de Maestría en Bioquímica], Facultad de Medicina, Universidad Nacional de Colombia sede Bogotá. 2000; pp 96.
18.Hammer Ø, Harper D, Ryan P. PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica. 2001;4(1):9.
19.Hammer Ø. PAST PAleontological Statistics version 2.16. Reference Manual. Natural History Museum, University of Oslo Norway. 2012; 227 p.
20.Roy A, Ananthakrishnan G, Hartung JS, Brlansky RH. Development and application of a multiplex reverse transcription polymerase chain reaction assay for screening a global collection of Citrus tristeza virus isolates. Phytopathology. 2010;100:1077-1088.
21.Folimonova S, Robertson C, Shilts T, Folimonov A, Hilf M, Garnsey S, Dawson W. Infection with strains of Citrus Tristeza Virus does not exclude superinfection by other strains of the virus. Journal of virology. 2010;84(3):1314-1325.
22.Tatinenia S, Robertsona C, Garnsey S, Dawson W. A plant virus evolved by acquiring multiple non conserved genes to extend its host range. PNAS. 2011;108(42):1736617371.
23.Silva G, Marques N, Nolasco G. The evolutionary rate of citrus tristeza virus ranks among the rates of the slowest RNA viruses. J Gen Virol. 2012;93:419-429.
24.Ananthakrishnan G, Venkataprasanna T, Roy A, Brlansky R. Characterization of the mixture of genotypes of a Citrus tristeza virus isolate by reverse transcription-quantitative real-time PCR. Journal of Virological Methods. 2010;164(1-2):75-82.
25.Licciardello G, Raspagliesi D, Bar-Joseph M, Catara A. Characterization of isolates of Citrus tristeza virus by sequential analyses of enzyme immunoassays and capillary electrophoresis-single-strand conformation polymorphisms. Journal of Virological Methods. 2012;181(2):139-147.
Recibido: 17-2-2012.
Aceptado: 26-9-2012.
†(Q.E.P.D.) In memoriam of José Peñaranda Valverde.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}