










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752
Rev. Protección Veg. vol.28 no.3 La Habana sep.-dic. 2013
COMUNICACIÓN CORTA
Efecto de las micorrizas arbusculares y Meloidogyne spp. en tomate (Solanum lycopersicum L.)
Effect of arbuscular mycorrhizas and Meloidogyne spp. on tomato (Solanum lycopersicum L.)
Alejandro AlarcónI, Tony BoicetI, María GodefoyII, Macario Bacilio-JiménezIII, Wilson CeiroI, Yurdanys BazánI
IUniversidad de Granma, Km 17½ carretera de Manzanillo-Bayamo, Apto 21, CP 85100, Bayamo, Granma, Cuba. Correo electrónico: aalarconz@udg.co.cu.
IIFacultad de Ciencias Médicas de Manzanillo, Universidad de Ciencias Médicas de Granma, CP 87 510, Manzanillo, Granma, Cuba.
IIICentro de Investigaciones Biológicas del Noroeste (CIBNOR). Mar Bermejo 195. Colonia Playa Palo de Santa Rita. C.P. 23070. La Paz, BCS. México.
RESUMEN
El estudio se desarrolló en la Universidad de Granma (Cuba) en el período comprendido de octubre/2007 a enero/2008, para determinar los efectos de las micorrizas arbusculares y Meloidogyne spp., en el cultivo del tomate (Solanum lycopersicum L.) cv. Vyta. Se utilizaron siete tratamientos, consistentes en aplicaciones simples y combinadas de un concentrado de cepas nativas y no nativas de micorrizas (Glomus mosseae (Gerdemann y Trappe) y Glomus sp. (Schenck y Smith), incluyendo un testigo y dos niveles poblacionales de Meloidogyne spp., con 30 repeticiones, los cuales se establecieron sobre un diseño completamente aleatorizado en condiciones semi-controladas, con arreglo factorial. Se utilizaron macetas de 2 kg de capacidad, que contenían un sustrato estéril, compuesto por una mezcla de suelo y estiércol ovino en proporción 3:1 v:v y una población de Meloidogyne spp. procedente de Granma. Se evaluaron las variables: índice de agallamiento e indicadores del crecimiento de las plantas: altura promedio (cm), masa seca promedio (g) y el rendimiento (kg.planta-1) a los 60 días después de la inoculación del nematodo. Los datos fueron procesados mediante un análisis de varianza y comparación de medias por la prueba del rango múltiple de Duncan (p<0,05). Los mejores resultados para los indicadores del crecimiento se obtuvieron cuando se aplicó el concentrado de cepas nativas y su combinación con Glomus mosseae y Glomus sp., y el menor índice de infestación del nematodo (1,5 huevos-juveniles infestivos (Ji).g-1 de sustrato). Sin embargo; al incrementar la concentración del inóculo a 2,5 huevos-juveniles infestivos (Ji).g-1 de sustrato, no hubo efecto significativo de las micorrizas arbusculares en la reducción de los daños producidos por Meloidogyne spp.
Palabras clave: Meloidogyne spp., Glomus mosseae, Glomus sp.
ABSTRACT
This study was carried out at the University of Granma (Cuba) from October 2007 to January 2008 in order to determine the effect of arbuscular mycorrhizas and Meloidogyne spp. on tomato (Solanum lycopersicum L.), var. Vyta. Seven treatments of single or combined applications of native and non-native strains(Glomus mosseae (Gerdemann and Trappe), and Glomus sp. (Schenck and Smith), of arbuscular mycorrhizas and an uninoculated control) and two levels of Meloidogine spp. were included in a completely randomized design set up in a factorial arrangement with thirty replicates under semi-controlled conditions. Two kg plastic pots containing a sterile substrate with a mixture of soil and ovine manure in a 3:1 v:v. ratio, as well as a population of Meloidogyne spp. from Granma were used. The gall index and plant growth parameters such as average height (cm), average dry weight (g) and crop yield (kg.plant-1) were evaluated 60 days after nematode inoculation. The data were analyzed by an analysis of variance and the means compared by Duncan's multiple range test (p<0,05). The results indicated that the plant growth parameters evaluated were higher when the native strain was combined with Glomus mosseae and Glomus sp. and the nematode infection level (1,5 eggs.g-1 substrate) was the lowest. However, when the inoculum concentration was increased to 2,5 eggs.g-1 substrate, no significant effect of the arbuscular mycorrhizas on the reduction of the damages produced by Meloidogyne spp. were observed.
Key words: Meloidogyne spp., Glomus mosseae, Glomus sp.
El tomate (Solanum lycopersicum L.) constituye una de las principales hortalizas del mundo y representa un cultivo fuertemente afectado por diferentes plagas, entre ellos los nematodos (1).
Se conoce que, los organismos fitoparásitos son componentes habituales de los sistemas agronómicos y se comprobó experimentalmente que el ataque de nematodos endoparásitos (Meloidogyne spp.) reduce el rendimiento agrícola de los cultivos entre 15 y 25% y en algunos casos hasta 75% (2).
Los nematodos y hongos micorrízicos arbusculares se encuentran relacionados en la rizósfera de las plantas, y ambos organismos interactúan en el momento de colonizar la raíz (2,3).
Los hongos micorrízicos arbusculares (HMA) forman asociaciones de tipo mutualista con las raíces de la mayoría de las plantas superiores y poseen importante función en la captación de nutrientes, mejoramiento del crecimiento y protección frente a patógenos radicales (4).
Pocas investigaciones se efectuaron en las condiciones edafoclimáticas de la provincia Granma (Cuba) relacionadas con el efecto de cepas de micorrizas arbusculares (certificadas introducidas y/o nativas) para reducir los daños producidos por nematodos endoparásitos del género Meloidogyne, las que ofrecerían elementos importantes para el manejo de las poblaciones de nematodos en las hortalizas.
El objetivo de este estudio fue determinar los efectos de las micorrizas arbusculares y Meloidogyne spp., en el cultivo del tomate cv. Vyta.
El experimento se realizó en la Universidad de Granma en el período comprendido entre octubre/2007 y enero/2008, en condiciones semi-controladas con temperatura de 25 ± 2° C y humedad relativa entre 81-86%. Se utilizaron macetas de 2 kg de capacidad contentivas de un sustrato, esterilizado en autoclave (121°C, 1 atm, por 2 h), compuesto por una mezcla de estiércol ovino y suelo del tipo Cambisol (5) en proporción 1:1 (v:v).
Se emplearon inóculos de un concentrado de cepas nativas (CCN) de micorrizas arbusculares (MA) (aisladas del suelo empleado para el experimento) y no nativas (Glomus mosseae, Gerdemann y Trappe) (cepa IES-6) y Glomus sp., Schenck y Smith (cepa IES-13), procedentes, reproducidas y certificadas por el Instituto de Ecología y Sistemática (IES), La Habana (Cuba), constituidas por una mezcla de esporas, esporocarpos, hifas y fragmentos de raíces.
Las cepas de micorrizas arbusculares (no nativas y nativas) presentaban las siguientes características de calidad micorrízica: 60-70 % de raíces infectadas y aproximadamente 250 esporas por gramo de suelo.
Las semillas de tomate del cv. Vyta (93 % de germinación) se sembraron en bandejas (cepellones) de 104 alvéolos, con capacidad de 32,5 cm2 cada uno y transcurridos 21 días desde la germinación, se trasplantó una planta por cada maceta. Las cepas de micorrizas se inocularon a una profundidad de 10 cm en el sustrato, a razón de 3 g.maceta-1 para la inoculación simple y de 1,5 g del CCN + 1,5 g de G. mosseae o Glomus sp., para la doble inoculación. Seguidamente se añadió a cada maceta, una suspensión del nematodo en dosis de 1,5 ó 2,5 huevos y/o juveniles infestivos (Ji).g-1 de sustrato (según el tratamiento).
El inóculo de Meloidogyne spp., se obtuvo a partir de una población nativa que infestaba Achyranthes aspera L., utilizando la metodología descrita por Hussey y Barker (6).
Se establecieron siete tratamientos y treinta repeticiones, con un diseño experimental completamente aleatorizado, donde cada maceta contenía una planta y representaba cada una de ella una repetición. Los tratamientos fueron: 1. Control (solo con nematodo); 2. G. mosseae; 3. Glomus sp.; 4. Concentrado de cepas nativas (CCN); 5. G. mosseae + Glomus sp.; 6. CCN + G. mosseae; 7. CCN + Glomus sp. Los tratamientos del 2 al 7 recibieron nematodos.
Se establecieron 30 plantas por tratamiento y se emplearon 10 para cuantificar el índice de agallamiento (IA), 60 días después de la inoculación de los nematodos, utilizando la escala de Taylor y Sasser (7).
Al final del experimento, se cosecharon las restantes 20 plantas y se evaluaron los indicadores: altura promedio de la planta (cm), masa seca promedio de la planta (g) y rendimiento (kg.planta-1).
A los datos obtenidos se les verificó la normalidad por la prueba de Kolmogorov-Smirnov y la homogeneidad de varianza por la prueba de Bartlett, referida por Sokal y Rohlf y fueron procesados estadísticamente mediante un análisis de varianza de clasificación simple (para cada nivel de inóculo) y se compararon las medias por la prueba paramétrica del rango múltiple de Duncan (p<0,05), empleando el paquete estadístico «STATISTICA» para Windows, versión 7.0.
La combinación de cepas de micorrizas provocó disminución del IA por debajo de 2, en las plantas que recibieron menor nivel de inóculo de Meloidogyne spp., mientras que en las que recibieron 2,5 huevos y/o juveniles por cada gramo de sustrato, los valores del IA se mantuvieron por encima de 3 (Tabla 1).
Se lograron los mejores resultados para los tratamientos, con las diferentes combinaciones de inóculos micorrízicos nativos o no nativos (tratamientos 5, 6 y 7), donde el índice de agallamiento disminuyó significativamente en comparación con la inoculación simple y el control (-M+N).
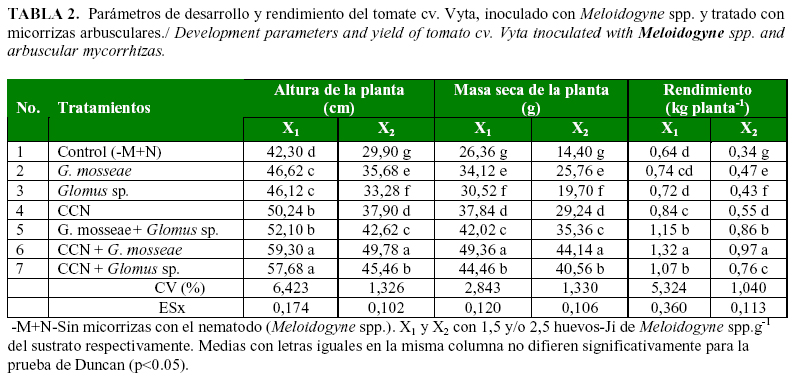
El efecto de los tratamientos en la reducción del índice de agallamiento por el nematodo, se reflejó en los indicadores del crecimiento (altura de la planta y masa seca de la planta) y el rendimiento (Tabla 2).
Dichos valores fueron mayores en las variantes, donde el nivel de inóculo fue menor. De igual modo, estas variables fueron significativamente mayores en los tratamientos, que recibieron la combinación de micorrizas arbusculares, CCN + G. mosseae en presencia de ambos niveles de Meloidogyne spp.
La simple y doble inoculación de micorrizas arbusculares produjo incrementos significativos de estos indicadores del crecimiento en comparación con el tratamiento control (-M+N), que mostró los peores resultados frente a ambos niveles poblacionales del nematodo.
Los valores más altos de IA se produjeron en los tratamientos sin micorrizas en ambos niveles de inoculo y se constató que este índice aumentó en las plantas, a medida que se incrementó la concentración del inoculo. Este fenómeno posee relación directa con la alta susceptibilidad de esta variedad a nematodos agalleros (8).
La presencia de los índices más bajos de agallamiento en los tratamientos, que recibieron doble combinación de micorrizas arbusculares (nativos o no nativos), sugieren que estas tienden a colonizar o infectar más agresivamente el sistema radical de la planta hospedante, que cuando se aplican las cepas por separado.
Numerosos estudios sugieren mecanismos de acción de las micorrizas en sus interacciones con los nematodos fitoparásitos, entre ellos, la competencia por el espacio; la adquisición de nutrientes por las plantas micorrizadas, que puede tener relación con la tolerancia de las plantas micorrizadas a los nematodos y el potencial de las micorrizas de inducir resistencia en las plantas a los patógenos. Sin embargo; los mecanismos moleculares acerca de cómo la simbiosis de plantas y micorrizas provee el control de nematodos son aun pobremente entendidos (9).
En estudios en Cuba, Solórzano et al. (10), informaron que las micorrizas arbusculares del género Glomus (Glomus manihotis y Glomus fasciculatum) incrementaron la resistencia del tomate (Solanum lycopersicum L.) variedad INCA-17 a la infección por patógenos, pues fueron capaces de inducir la síntesis de enzimas relacionadas con la defensa contra dichos patógenos como: peroxidasas, ß(1-3)-glucanasas, fenilalanina-amonioliasas, quitinasas y polifenoloxidasas. Los mecanismos involucrados en el sistema planta-micorrizas-nematodos deberán ser objeto de estudios futuros, de manera de lograr una eficiente explotación de las micorrizas en el manejo de nematodos en hortalizas como el tomate.
Estos resultados están en correspondencia con los obtenidos e informados por Hernández-Socorro (11) para el cultivo del tomate (S. lycopersicum), y por los reportados por Rodríguez-Romero y Jaizme-Vega (12) para el plátano (Musa spp.) y Castillo et al. (13) en plantas de oliva, evaluando el efecto de micorrizas arbusculares en la protección contra nematodos agalleros en condiciones de suelo y clima de España.
Los resultados obtenidos en este estudio indican que no existe una simple respuesta en la interacción planta-micorriza arbuscular-nematodo y que dicha respuesta depende de factores como las especies de nematodo, micorriza y planta, factores ambientales, tiempo del proceso de micorrización y período de exposición al nematodo.
Por lo tanto, futuras investigaciones se requerirán para seleccionar las cepas de micorrizas que puedan proteger más eficientemente a este cultivo contra los daños del nematodo y para estudiar con mayor profundidad los mecanismos involucrados en dicha interacción. En tal sentido, es recomendable usar la micorrización en el cultivo de tomate, como medida para reducir los daños causados por Meloidogyne spp., siempre que las poblaciones de la plaga no sean significativamente altas.
REFERENCIAS
1. Maroto JV. Consumo de hortalizas y salud. Agrícola Vergel. 2008; 315:138-143.
2. Karssen G, Moens M. Root-knot nematodes. In: Perry R, Moens M, editors. Plant Nematology. CABI Publising, UK. p.59-90. 2006.
3. Rodríguez-Romero AS, Jaizme-Vega MC. Ed. Khan MS, Zaidi A, Mussarat J. Contribution of arbuscular mycorrhizal fungi and plant growth-promoting rhizobacteria to plant health and development. Microbes in Sustainable Agriculture. Editorial Nova Science Publishers. Inc. 2008.
4. Gianinazzi S, Gollotte A, Binet MN, Tuinen DV, Redecker D, Wipf D. Agroecology: the key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza. 2010;20:519-530.
5. Hernández A, Pérez J, Bosch A, Rivero LD. Nueva versión de clasificación Genética de los suelos de Cuba. Instituto de Suelos. Ministerio de la Agricultura. Editora AGRINFOR. La Habana, Cuba. 64 pp. 1999.
6. Hussey RS, Barker KR. A comparison of methods of collecting inocula of Meloidogyne spp., incluiding a new technique. Plant Dis Res. 1973;57:1025-1028.
7. Taylor AL, Sasser JB. N.C. State Univ. Biology, identification and control of root-knot nematodes (Meloidogyne spp.). Raleigh. 1978.
8. Rodríguez MG, Gómez L, González FM, Carrillo Y, Piñón M, Gómez O, et al. Comportamiento de la familia Solanaceae frente a Meloidogyne incognita (Kofoid y White) Chitwood. Rev Protección Veg. 2009;24(3):137-145.
9. Hallmann J, Sikora RA. Endophytic Fungi. En: K. Davis and Y. Spiegel (Eds.). Biological Control of Plant Parasitic Nematodes. 227 Building Coherence between Microbial Ecology and Molecular Mechanisms, Progress in Biological Control 11, Springer Science + Business Media, B.V. Pp 227-258. 2011.
10.Solórzano E, Meneses AR, Rodríguez Y, Pérez E, Fernández A, Peteira B, et al. Inducción de cinco sistemas enzimáticos en la simbiosis tomate-micorriza arbuscular. Rev Protección Veg. 2001;16(1):30-39.
11.Hernández-Socorro MA. Interacción de Glomus mosseae-Pochonia chlamydosporia var. catenulata y Meloidogyne incognita en tomate (Solanum lycopersicum L.). Rev Protección Veg. 2010;25(2):136.
12.Rodríguez-Romero AS, Jaizme-Vega MC. Effect of the arbuscular mycorrhizal fungus, Glomus manihotis on the root-knot nematode, Meloidogyne javanica in banana. Nematologia Mediterranea. 2005;33:217-221.
13.Castillo P, Azcón-Aguilar C, Del Rincón R, Calvet C, Jiménez RM. Protection of olive planting stocks against parasitism of root-knot nematodes by arbuscular mycorrhizal fungi. Plant Pathology. 2006;5:705-713.
Recibido: 30-1-2013.
Aceptado: 9-5-2013.

{kind=link}
{kind=link}