










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752
Rev. Protección Veg. vol.29 no.1 La Habana ene.-abr. 2014
ARTÍCULO RESEÑA
Potencialidades de las bacterias diazotróficas asociativas en la promoción del crecimiento vegetal y el control de Pyricularia oryzae (Sacc.) en el cultivo del arroz (Oryza sativa L.)
Potentialities of associated diazotrophic bacteria in plant growth promotion and biocontrol of Pyricularia oryzae (Sacc.) in rice (Oryza sativa L.)
Annia Hernández-RodríguezI*, Narovis Rives-RodríguezII, Yanelis Acebo-GuerreroI, Acela Diaz-de la OsaI, Mayra Heydrich-PérezI, Vera Lucia Divan BaldaniIII
IDepartamento de Microbiología y Virología, Facultad de Biología, Universidad de La Habana, Calle 25 # 455 e/ J e I, Vedado, La Habana, Cuba.
IIInstituto de Investigaciones de Granos. Autopista Novia del Mediodia, Km 16 ½ Bauta, Artemisa, Cuba.
IIILaboratorio de Gramíneas. EMBRAPA Agrobiología. Seropédica, Rio de Janeiro, Brasil.
RESUMEN
Este trabajo tuvo como objetivos abordar las potencialidades de uso de bacterias diazotróficas asociativas en la producción sostenible del cultivo del arroz (Oryza sativa L.), tratando temas como la fertilización nitrogenada y su relación con la incidencia en la piriculariosis, añublo o quemazón, así como sus perspectivas de uso en la promoción del crecimiento y control de Pyricularia oryzae Sacc. Al aplicar bacterias diazotróficas asociativas se logran aportes de nitrógeno atmosférico a la planta que permiten disminuir el uso de productos químicos nitrogenados y con ello limitar el crecimiento de P. oryzae. Estas bacterias, además, liberan metabolitos en la rizosfera que disminuyen la concentración de la plaga y la severidad de la enfermedad, lo que trae consigo reducciones en la aplicación de productos químicos y menor incidencia de la piriculariosis en condiciones de campo. Cepas autóctonas cubanas tienen la capacidad de controlar in vitro a P. oryzae, lo que sugiere que las bacterias diazotróficas asociativas que estimulan el crecimiento vegetal, pudieran disminuir los efectos nocivos ocasionados por esta plaga en el cultivo del arroz.
Palabras clave: Bacterias diazotróficas, arroz, nitrógeno, Pyricularia oryzae.
ABSTRACT
This work was aimed at approaching the potentialities of associated diazotrophic bacteria for the sustainable production of rice (Oryza sativa L.). Relevant topics such as nitrogen fertilization, its relationships with rice blast incidence, and the perspectives of use of associated diazotrophic bacteria for plant growth promotion and biocontrol of Pyricularia oryzae in this important crop are discussed. Associated diazotrophic bacteria have been shown to contribute with fixed atmospheric nitrogen to the rice crop making possible to reduce the application of nitrogen fertilizers, limiting with it the growth of Pyricularia oryzae. In addition, the metabolites released by these bacteria in the rhizosphere make the plague concentration and the disease severity decrease leading to the reduction of chemical product applications and a lower incidence of rice blast under field conditions. Cuban autochthonous strains have the ability to control Pyricularia oryzae in vitro, which confirms the fact that associated diazotrophic bacteria stimulate plant growth and mitigate the deleterious effects caused by this disease in rice.
Key words: Diazotrophic bacteria, rice, nitrogen, Pyricularia oryzae.
INTRODUCCIÓN
El arroz (Oryza sativa L.) representa el alimento básico para más de la mitad de la población mundial, considerado el cultivo más importante del mundo por la extensión de la superficie que ocupa y la cantidad de personas que dependen de su cosecha. Su producción promedio anual alcanza, aproximadamente, unos 650 millones de toneladas (1). Sin embargo, a pesar de su importancia como cultivo alimenticio, las producciones del grano no cubren la demanda mundial, entre otros factores, por la gran explosión demográfica en los últimos años y limitaciones en la disponibilidad de suelo y agua (2).
En Cuba, el arroz constituye parte importante en la dieta diaria de la población, con consumo per cápita actual cercano a los 60 kg anuales, uno de los más altos de América Latina, aportando el 20 % de las calorías que se consumen diariamente (3). Por ello, representa un reto actual, lograr mayores producciones del cereal, sin que esto conlleve al aumento de las aplicaciones de fertilizantes químicos, fundamentalmente nitrógeno, los que además de ser excesivamente costosos, provocan la contaminación de los suelos agrícolas y el ambiente en general.
El nitrógeno es un elemento limitante para la productividad vegetal y su exceso favorece la aparición de Pyricularia oryzae Sacc., principal patógeno que afecta al cultivo y que puede causar pérdidas de hasta el 80% de los rendimientos agrícolas (4).
Los inoculantes microbianos a base de bacterias diazotróficas, constituyen una alternativa eficaz al uso de fertilizantes nitrogenados (5, 6). Estas bacterias tienen la capacidad de reducir el nitrógeno atmosférico, donde se encuentra como nitrógeno elemental de forma ilimitada, y hacerlo disponible para los cultivos. Los fijadores de nitrógeno pueden interactuar de forma simbiótica con la planta, como en el caso rizobio-leguminosa, o comportarse como diazótrofos asociativos (7).
Aunque las potencialidades de las bacterias diazotróficas ofrecen perspectivas prometedoras, la ausencia de cepas autóctonas caracterizadas en cuanto a sus mecanismos de acción imposibilitó hasta hoy, la obtención de productos eficientes que puedan ser aplicados para mitigar el deterioro ambiental causado por el uso excesivo de insumos químicos y para lograr la producción sostenible del cultivo del arroz.
Este trabajo tuvo como objetivo ofrecer una panorámica actual acerca de las potencialidades de uso de las bacterias diazotróficas asociativas en la promoción del crecimiento vegetal y el manejo de P. oryzae en el cultivo del arroz (O. sativa) en Cuba.
PARTE ESPECIAL
Piriculariosis o añublo del arroz: Importancia
En el cultivo del arroz, las enfermedades de origen microbiano constituyen uno de los factores que inciden en la obtención de bajos rendimientos, calidad y manchado de los granos. Estas pueden ser provocadas por hongos y bacterias, constituyendo las de origen fúngico las más frecuentes.
Una gran diversidad de hongos que habitan en el ecosistema aéreo fueron identificados como patógenos del arroz, entre ellos, Curvularia aeria (Bat., J.A. Lima & C.T. Vasconc.), Curvularia affinis Boedijn, Curvularia brachyospora Boedijn, Curvularia clavata B.L. Jain, Curvularia eragrostridis (Henn.) J.A. Mey., Curvularia lunata (Wakker) Boedijn, Curvularia pallescens Boedijn, Curvularia trifolii (Kauffman) Boedijn, Bipolaris australiensis (M.B. Ellis) Tsuda & Ueyama, Bipolaris hawaiiensis (M.B. Ellis) J.Y. Uchida & Aragaki, Bipolaris oryzae (Breda de Haan) Shoemaker, Bipolaris sorokiniana (Sacc.) Shoemaker y Pyricularia oryzae Cavara (8). Estos hongos pueden afectar directamente a la planta o al grano, provocando diferentes enfermedades (9).
La piriculariosis o añublo del arroz, causado por P. oryzae, es una de las enfermedades más importantes en este cultivo por su amplia distribución mundial, poder destructivo y alta patogenicidad (10, 11). Los síntomas pueden aparecer en toda la planta en diferentes etapas de su desarrollo (4). Así por ejemplo, en la parte aérea la enfermedad se manifiesta como manchas elípticas a romboides, cuyo tamaño y color varían de acuerdo con las condiciones ambientales y con la susceptibilidad de los cultivares (12). El hongo P. oryzae tiene la capacidad de infectar la raíz provocando síntomas típicos, similares a los inducidos por otras enfermedades fúngicas (13)
La enfermedad se presenta generalmente, a partir de los 35 días de la siembra, incidiendo durante la fenofase de ahijamiento activo, dismunuyendo en la fase de cambio de primordio e incide nuevamente en la paniculación (14). P. oryzae puede permanecer en los residuos de cosecha, las semillas y en otras plantas hospedantes. En los residuos de cosecha, es probable encontrar inóculo del patógeno hasta después de dos años de ser eliminadas las plantas. Almaguer-Chavez et al. (11) demostraron que la temperatura y la humedad relativa, influyen positivamente en la concentración de propágalos de P. oryzae en el aire.
Los principales aspectos que se deben considerar para evitar los daños causados por esta plaga están relacionados con el uso de cultivares resistentes o tolerantes, densidad de siembra adecuada (no superior a 150 o 200 plantas/ m2) y dosis correctas de fertilizante nitrogenado (15). También es importante tener en cuenta el manejo del agua, aumentando la lámina de agua cuando hay incidencia de la enfermedad, así como el uso de tratamiento químico y biológico a la semilla, mediante la incorporación de restos de vegetales al suelo y el uso de los fungicidas específicos (4).
Fertilización nitrogenada e incidencia de P. oryzae
La planta de arroz demanda nitrógeno desde las fases iniciales de su ciclo de vida hasta el inicio de la maduración, coincidiendo la mayor demanda con el punto de algodón. La fertilización nitrogenada en el cultivo se realiza de forma fraccionada para favorecer mayor eficiencia en la nutrición, ya que la planta no tiene capacidad biológica para absorber dosis superiores a 50 kg.ha-1 de nitrógeno. El primer fraccionamiento se realiza en la etapa de inicio de ahijamiento, para favorecer el crecimiento de la planta y su ahijamiento, mientras que los últimos fraccionamientos se realizan en la etapa de punto de algodón y 10 días después, para favorecer el mayor número de granos llenos por panícula y el área foliar (3).
Los principales portadores de fertilizantes nitrogenados son: urea (46% de N), sulfato de amonio (21% de N) y amoniaco anhidro (82% de N). El rendimiento del cultivo (t. ha-1) es proporcional a los kg de N. ha-1 (3). Sin embargo, cuando las condiciones ambientales son favorables y se aplican elevadas dosis de fertilización nitrogenada, se incrementa la severidad de la piriculariosis, se producen pérdidas superiores al 70% del rendimiento agrícola y muchos de los granos cosechados pierden calidad (4). En este sentido, Pinciroli et al. (14), demostraron que las dosis de nitrógeno y el momento de aplicación, influyen en la ocurrencia y severidad de esta enfermedad, así como en el tipo y tamaño de la lesión.
Estos aspectos están relacionados con la fisiología del patógeno, pues al existir mayor disponibilidad de fertilizantes químicos nitrogenados en el suelo y la planta no absorber dosis superiores a 50 kg. ha-1 de nitrógeno, los compuestos residuales son aprovechados por P. oryzae para su nutrición (16). Esto permite que se incremente la concentración del patógeno y se alcancen concentraciones superiores del inóculo, lo que provoca que se disemine la enfermedad (17).
Bacterias diazotróficas asociativas: principales mecanismos de acción
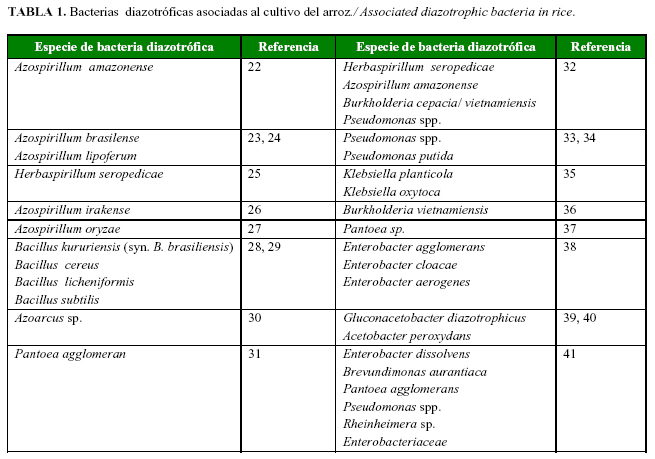
Los diazotrofos incluyen representantes de arquibacterias, cianobacterias y bacterias gram positivas y negativas, que poseen una amplia diversidad morfológica, fisiológica y genética. En particular, las bacterias diazotróficas están representadas por diversos grupos filogenéticos que tienen la capacidad de fijar nitrógeno atmosférico, pueden vivir libres en diversos ecosistemas, establecer simbiosis o estar asociadas a las plantas (18). Se ubican en diferentes géneros como Azotobacter, Beijerinckia, Derxia, Azospirillum, Herbaspirillum, Burkholderia, Gluconacetobacter, Azoarcus, Bacillus, Paenibacillus, Klebsiella, Enterobacter, Pantoea, Citrobacter y Serratia (5, 19).
Diferentes especies de bacterias diazotróficas asociativas fueron aisladas de las raíces y parte aérea de cultivos de importancia agrícola, como gramíneas (18, 19), tubérculos (20) y frutales (21). Se encuentran colonizando la rizosfera de los cultivos en concentraciones del orden de 107ufc.g de suelo-1 (18). En el cultivo del arroz, la mayor cantidad de estudios están relacionados con los géneros Azospirillum y Herbaspirillum; sin embargo, existe una gran diversidad de bacterias diazotróficas asociadas a esta gramínea (Tabla 1).
Las bacterias diazotróficas, además del aporte de nitrógeno que hacen a través de la Fijación Biológica (FBN), pueden promover de forma directa el crecimiento de las plantas mediante otros mecanismos de acción. Estos efectos directos se evidencian en ausencia de otros microorganismos, es decir, en la interacción planta-microorganismo, e incluyen la solubilización de minerales (9) y la síntesis de fitohormonas (42). Se observó además, su influencia en la absorción de elementos minerales, debido a incrementos en los flujos iónicos de la superficie de las raíces (42). Los mecanismos indirectos se ponen de manifiesto cuando ocurre la interacción del microorganismo de interés con un fitopatógeno, provocando la disminución de los efectos dañinos de este último sobre la planta (4, 43).
Aporte de nitrógeno a la planta
La FBN realizada por las células procariotas, es el proceso mediante el cual, parte del nitrógeno atmosférico se incorpora a la materia viva y constituye la principal vía de incorporación de nitrógeno al ecosistema del suelo, siendo devuelto a la atmósfera principalmente, por la acción de organismos del suelo descomponedores de la materia orgánica.
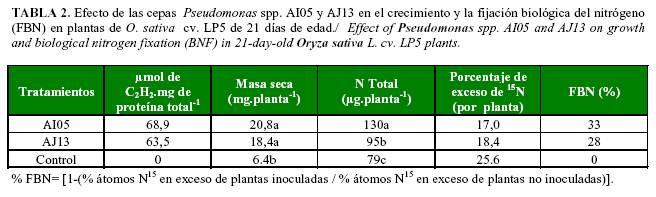
Diversos estudios revelaron que los organismos diazotróficos colonizan sus hospedantes, contribuyendo con cantidades sustanciales de nitrógeno fijado biológicamente (44). Baldani et al. (45) inocularon plantas de arroz con cepas de Herbaspirillum seropedicae y Burkholderia spp. en condiciones in vitro, logrando aportes de nitrógeno a la planta entre 31 y 54%. Asimismo, Kennedy et al. (46) señalaron que algunas especies de Herbaspirillum aportaron a la planta de arroz del 19 al 54% del nitrógeno requerido para su crecimiento y desarrollo, en dependencia del cultivar. En bioensayos in vitro desarrollados en EMBRAPA Agrobiología, utilizando Sulfato de amonio marcado con 15N, se demostró que las cepas autóctonas cubanas Pseudomonas spp. AI05 y AJ13 tuvieron la capacidad de llevar a cabo la FBN, con contribuciones de nitrógeno a la planta de entre 28 y 33% (Tabla 2).
En esta investigación se evidenció que AI05 y AJ13 son cepas de bacterias diazotróficas que tienen potencialidades de uso como inoculantes microbianos en el cultivo del arroz. Otros resultados experimentales demostraron además, su eficiencia en la estimulación del crecimiento vegetal de los cultivares J-104 y Perla de Cuba, a través de incrementos en el área foliar, área de la raíz y el rendimiento del cultivo (Datos no publicados). Sin embargo, para diseñar un inoculante microbiano con máxima potencialidad en los distintos cultivos, suelos y climas donde estos sean aplicados, se debe tener en cuenta la microbiota nativa, la optimización del medio de cultivo y la fertilidad del suelo (6).
Incremento de la disponibilidad de fósforo en el suelo
Varios procesos importantes de la Fisiología vegetal están asociados a la nutrición del fósforo y determinan el papel limitante de este elemento para el desarrollo de las plantas. De igual manera, el fósforo incide en la fijación biológica del nitrógeno, en la calidad y el rendimiento de los cultivos y en la resistencia de las plantas a las enfermedades (47). Sin embargo, es el elemento de menor disponibilidad en el suelo, debido a su inmovilización por microorganismos y a la precipitación (fijación) en complejos minerales insolubles, lo que trae como consecuencia una disminución de hasta el 75% del fósforo disponible. En condiciones naturales, este nutriente se encuentra tanto en formas orgánicas como inorgánicas (48).
Las bacterias diazotróficas realizan la solubilización de fosfatos orgánicos, mediante un proceso catalizado por enzimas y la de fosfato inorgánico, a través de la producción de ácidos orgánicos (47). La liberación de ácidos orgánicos por las bacterias provoca disminución del pH en el medio, lo que facilita la liberación de fósforo inmovilizado, mediante la sustitución y excreción de H+ o la liberación de iones Ca2+ (47). Otro mecanismo está relacionado con la formación de complejos estables que permiten la liberación de los grupos fosfatos. Estos complejos se forman debido a la alta afinidad que tienen los aniones carboxílicos de los ácidos orgánicos liberados por las bacterias, por el calcio y otros iones metálicos, como el hierro y el aluminio, presentes en los fosfatos minerales (48).
Las bacterias solubilizadoras de fosfato están presentes en formas y poblaciones variadas en los diferentes suelos, e incluyen bacterias aerobias y anaerobias. Entre toda la población microbiana del suelo, estas constituyen del 1 al 50% y la mayor proporción se concentra en la rizosfera (49). Sin embargo, se ha demostrado que no todas las bacterias que forman halos de solubilización en medio NBRIP sólido, son verdaderas solubilizadoras de fósforo en condiciones de campo (50).
Bashan et al. (50), señalaron que para comprobar que un microorganismo solubiliza fosfato, después de la determinación de la formación de halo en medio sólido con fuentes de fósforo orgánico e inorgánico, se debe cuantificar el fósforo soluble en medio líquido y posteriormente determinar la producción de ácidos, entre ellos, ácido oxálico, fumárico y cítrico. Además, se debe determinar la capacidad de las bacterias para solubilizar fosfatos orgánicos vía enzimática. La nueva metodología de selección de estas bacterias debe incluir, bioensayos in vivo e in situ que permitan determinar el aporte de fósforo a la planta y la contribución de este mecanismo a la estimulación del crecimiento vegetal.
Producción de fitohormonas y reguladores del crecimiento vegetal
La producción de fitohormonas por parte de las bacterias y su impacto en la morfogénesis de la raíz, podrían explicar en gran parte los efectos positivos de las bacterias diazotróficas en el crecimiento vegetal. Al incrementarse el número de pelos radicales y raíces laterales, existe mayor absorción de agua y nutrientes por la planta (42) y un mayor intercambio con el ambiente de la rizosfera, lo que contribuye a incrementos en el rendimiento del cultivo.
La fitoestimulación está dada particularmente por la compleja y balanceada red de hormonas de plantas o compuestos similares, que influyen directamente en el crecimiento o estimulan la formación de raíces. Por ejemplo, muchas especies de Azospirillum producen auxinas, citoquininas y giberelinas (4) que estimulan el desarrollo del sistema radicular, lo que provoca incrementos en los rendimientos agrícolas (51).
La producción bacteriana de Ácido Índol Acético (AIA) fue estudiada no solo por su efecto fisiológico en las plantas, sino también por la función que pueda desempeñar esta fitohormona en la interacción planta-microorganismo (29, 41, 42).
Control biológico de patógenos
El control biológico de microorganismos patógenos puede ocurrir por diferentes mecanismos de acción, tales como la competencia por nutrientes, minerales y espacio; la síntesis de metabolitos y la inducción de resistencia sistémica en la planta (52). Entre estos metabolitos se destacan los sideróforos, que hacen al hierro inaccesible para los patógenos; los antibióticos, tales como 2,4-diacetilfloroglucinol (DAPG), pioluteorina, pirrolnitrina y derivados de fenazina (53), y lipopeptidos cíclicos (54). La detoxificación de factores de virulencia de los patógenos constituye otro mecanismo de control biológico que puede incluir la producción de una proteína que se une, reversiblemente, a la toxina, la detoxificación irreversible mediada por una esterasa y la hidrólisis (4, 52).
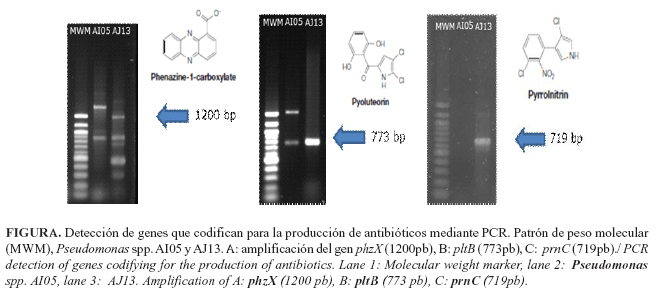
Estudios realizados en el Laboratorio de Ecología Microbiana de la Facultad de Biología de la Universidad de La Habana (Cuba), demostraron que las cepas de Pseudomonas spp. AI05 y AJ13 presentan genes que codifican para la producción de sideróforos y antibióticos. Ambas tienen la capacidad de producir pioluteorina y Pseudomonas spp. AJ13, adicionalmente, fenazina-1-carboxilato y pirrolnitrina (Figura). Sin embargo, bioensayos in vitro deben ser realizados para confirmar estas potencialidades.
La excreción de sideróforos por parte de los microorganismos, ocurre cuando estos se encuentran en medios con bajas concentraciones de hierro. Una vez formados los complejos sideróforo-ion férrico, estos son reconocidos e internalizados en la célula por proteínas receptoras específicas que se encuentran en la membrana celular (53).
La antibiosis es definida como la inhibición o destrucción de un microorganismo por los productos metabólicos de otros, que incluyen la acción de antibióticos, enzimas líticas, así como, compuestos volátiles y difusibles. Dentro de las enzimas líticas se estudiaron las quitinasas, las lipasas y las celulasas (54).
Las b-glucanasas y las proteasas, constituyen otro grupo de enzimas líticas relacionadas con el control biológico de fitopatógenos (55). Hernández-Rodríguez et al. (6) demostraron que Pseudomonas spp. cepas AI05 y AJ13 tienen efecto antagónico ante Curvularia pallescens Boedijn AAD 430 y Curvularia trifoli (Kauffman) Boedijn AAD 533, lo que sugieren que estas bacterias diazotróficas producen metabolitos que inhiben el crecimiento micelial de patógenos fúngicos. Sin embargo, el crecimiento de estos antagonistas sobre el patógeno en cultivo dual, no es garantía de alta capacidad biorreguladora del patógeno, ya que en la práctica se deben tener en cuenta otros aspectos que se relacionan con la interacción planta-microorganismos-medio ambiente. Esto evidencia, que la selección de aislamientos promisorios para el control de plagas debe incluir experimentos de interacción planta-microorganismos, que permitan establecer la capacidad de los aislados para activar resistencia sistémica en plantas (ISR, Induced Systemic Resistance, de sus siglas en inglés) contra diferentes tipos de patógenos.
Potencialidades de uso de las bacterias diazotróficas asociativas para el control de Pyricularia oryzae L. en el cultivo del arroz
Las bacterias diazotróficas asociativas con capacidad para promover el crecimiento de las plantas, representan una fuente interesante para utilizar, con el objetivo de elaborar productos eficientes que pudieran ser utilizados en el manejo integrado de plagas en cultivos de importancia económica, y específicamente de P. oryzae en el cultivo del arroz.
En este contexto, con la aplicación de bacterias diazotróficas como inoculantes microbianos se logra un mayor aporte de nitrógeno atmosférico a la planta (5,15,18, 22), lo que permite disminuir el uso de productos químicos nitrogenados. De este modo se limita el crecimiento de plagas que necesitan de fuentes nitrogenadas para su nutrición (16), a la vez estas bacterias liberan metabolitos en la rizosfera que contribuyen al equilibrio ecológico del suelo. Así por ejemplo, si se tiene en cuenta que en Cuba para obtener 4 t. ha-1de arroz es necesario aplicar 117 kg.N-1 y que las bacterias diazotróficas Pseudomonas spp. AI05 y AJ13 tienen la capacidad de aportar a la planta entre el 33 y 28% del nitrógeno total que necesita, la aplicación de biofertilizantes a base de estas bacterias permitiría reducir entre 32,76 y 38,61 kg de N.ha-1, respectivamente. Estas reducciones en la aplicación de fertilizantes nitrogenados, podrían afectar la fisiología de cepas de P. oryzae que necesitan de este elemento para su propia nutrición, lo que provocaría la disminución de las concentraciones de la plaga y menor incidencia de la piriculariosis en condiciones de campo. Sin embargo, la demostración de esta hipótesis requerirá de investigaciones futuras.
Es importante tener en cuenta que, las potencialidades de las bacterias diazotróficas dependen de la cepa, ya que aislados de una misma especie pueden tener diferentes mecanismos de acción, lo que les permite actuar de forma directa o indirecta sobre la plaga. Esto refuerza la necesidad de trabajar con cepas autóctonas, procedentes de los ecosistemas arroceros, de forma tal que se puedan obtener resultados repetibles y consistentes cuando son aplicados estos productos en condiciones de campo.
CONCLUSIONES
La utilización de microorganismos con capacidad para promover el crecimiento de las plantas se presenta como una alternativa de biofertilización. Estudios desarrollados a escala de laboratorio, macetas y campo demostraron que las bacterias diazotróficas asociativas pueden mitigar los efectos nocivos ocasionados por P. oryzae en O. sativa, lo que denota sus potencialidades de uso en el manejo integrado del cultivo. La adquisición de nutrientes del suelo a través del crecimiento radical, la fijación biológica del nitrógeno, la solubilización de fosfatos, la producción de fitohormonas y de metabolitos antifúngicos, hicieron posible la elaboración de productos biológicos eficientes a partir de bacterias diazotróficas, con características amigables con el ambiente. Sin embargo, el éxito de estos inoculantes bacterianos depende de la selección de cepas autóctonas.
AGRADECIMIENTOS
Este estudio fue financiado por el Proyecto CAPES/MES 110/11 «Aplicación de Bacterias diazotróficas promotoras del crecimiento vegetal en la producción sustentable del cultivo del arroz (Oryza sativa L.)».
REFERENCIAS
1. Food and Agriculture Organization of the United Nations (FAO). FAO production yearbook. Roma, Italia. 2008. 78 pp.
2. Ministerio de la Agricultura (MINAG). Instructivo Técnico de Arroz. Centro Nacional de Sanidad Vegetal, Instituto de Investigaciones del Arroz. 2006: 50-55.
3. Food and Agriculture Organization of the United Nations (FAO). FAO production yearbook. Roma, Italia. 2010. 50 pp.
4. Acebo-Guerrero Y, Hernández-Rodríguez A, Rives-Rodríguez N, Velázquez del Valle MG, Hernández-Lauzardo AN. Perspectivas del uso de bacterias rizosféricas en el control de Pyricularia grisea (CookeSacc.) en el cultivo del arroz (Oryza sativa L.). Rev Colomb Biotecnol. 2011;XIII(1):16-22.
5. Pedraza RO, Teixeira KRS, Fernández Scavino A, García de Salamone I, Baca BE, Azcón R, et al. Microorganismos que mejoran el crecimiento de las plantas y la calidad de los suelos. Revista Corpoica-Ciencia y Tecnología Agropecuaria. 2010;11(2):155-164.
6. Hernández-Rodríguez A, León-Plasencia D, Rives-Rodríguez N, Díaz de la Osa A, Almaguer-Chávez M, Acebo-Guerrero Y. Identificación de aislamientos autóctonos de pseudomonas fluorescentes con actividad antagónica ante Curvularia spp. Protección Vegetal. 2010;26(2):21-29.
7. Atlas R, Bartha R. Microbial ecology: fundamentals and applications. 3rd ed. Benjamin/Cummings Publishing Company. 1993.
8. Almaguer-Chávez M, Rojas TI, Dobal V, Batista A, Aira MJ. Effect of temperature on growth and germination of conidia in Curvularia and Bipolaris species isolated from the air. Aerobiología. 2012.
9. Barrios LM, Pérez IO. Nuevos registros de hongos en semillas de Oryza sativa en Cuba. Manejo Integrado de Plagas y Agroecología. 2005;75:64-67.
10.Rodríguez AT, Ramirez MA, Cárdenas RM, Hernández AN, Velazquez MG, Bautista S. Induction of defense response of Oryza sativa L. against Pyricularia grisea (Cooke) Sacc. by treating seeds with chitosan and hydrolyzed chitosan. Pesticide Biochemistry and Physiology. 2007;89(3):206-215.
11.Almaguer-Chávez M, Rojas TI, Dobal V, Batista A, Rives N, Aira MJ, et al. Aerobiological dynamics of potentially pathogenic fungi in a rice agroecosystem in La Habana. Aerobiología. 2012;28:177-183.
12.Cárdenas RM, Pérez N, Cristo E, González MC, Fabre L. Estudio sobre el comportamiento de líneas y variedades de arroz (Oryza sativa Lin.) ante la infección por el hongo Pyricularia grisea Sacc. Cultivos Tropicales. 2005;26(4):83-87.
13.Sesma A, Osbourn AE. The rice leaf blast pathogen undergoes developmental processes typical of root-infecting fungi. Nature. 2004;431:582-586.
14.Pinciroli M, Cordo MC, Bezus R, Vidal AA, Delucis M. Development of rice blast under two nitrogen availability conditions. Summa Phytopathologica. 2006;32(3):280-282.
15.Pedraza RO, Bellone CH, Carriz S, Fernandes PM, Santos Teixeira KR. Azospirillum inoculation and nitrogen fertilization effect on grain yield and on the diversity of endophytic bacteria in the phyllosphere of rice rainfed crop. European Journal of Soil Biology. 2009;45:36-43.
16.Brook TD, Madigan MT. Biology of Microorganisms. La Habana: ed. Felix Varela. 2008; 93-131.
17.Agrios GN. Burlington MA. Plant Pathology. Plant diseases caused by fungi. 5th ed. Elsevier. 2005; p. 724-820.
18.Souza Moreira FM, Da Silva K, Abrahão Nobrega RS, De Carvalho F. Bactérias diazotróficas associativas: diversidade, ecología e potencial de aplicações. Comunicata Scientiae. 2010;1(2):74-99.
19.Döbereiner J. History and new perspectives of diazotrophs in association with non-leguminous plants. Symbiosis. 1992;13:1-13.
20.Paula MA. Interação micorrizas vesículo arbusculares-bactérias diazotróficas em batata doce (Ipomoea batatas (L.) Lam. 168f. Tese de Doutorado- Universidade Federal Rural do Rio deJaneiro, Itaguaí, Brasil, 1992.
21.Weber OB, Baldani VLD, Teixeira KRS, Kirchhof G, Baldani JI, Döbereiner J. Isolation and characterization of diazotrophic bacteria from banana and pineapple plants. Plant and Soil. 1999;210:103-113.
22.Baldani JI, Baldani VLD, Sanpaio MJAM, Döbereiner J. A fourth Azospirillum species from cereal roots. Anais da Academia Brasileira de Ciência. 1984;56:365.
23.Döbereiner J, Pedrosa FO. Nitrogen-fixing bacteria in non-leguminous crop plants. Science Tech, Madison, USA. 155p, 1987.
24.Acebo Y, Rives N, Heydrich M, Hernández-Rodríguez A. Efecto promotor del crecimiento vegetal de cepas de Azospirillum en el cultivo del arroz (Oryza sativa). CultivosTropicales. 2007;28(3):29-32.
25.Baldani JI, Baldani VLD, Seldin L, Döbereiner J. Characterization of Herbaspirillum seropedicae nov. sp. a root associated nitrogen fixing bacterium. International Journal of Systematic Bacteriology. 1986;36:86-93.
26.Khammas KM, Ageron E, Grimont PA, Kaiser P. Azospirillum irakense nov. sp. a nitrogen fixing bacterium associated with rice roots and rhizosphere soil. Research in Microbiology. 1989;140:679-693.
27.Xie C, Yokota A. Azospirillum oryzae sp. nov., a nitrogen-fixing bacterium isolated from the roots of the rice plant Oryza sativa. International Journal of Systematic and Evolutionary Microbiology. 2005;55:1435-1438.
28.Xie GH, Su BL, Cui ZJ. Isolation and identification of N2-fixing strains of Bacillus in rice rhizosphere of the Yangtze River valley. Acta Microbiologica Sinica. 1998;38:480-483.
29.Rojas MM, Larrea JA, Mahillon J, Heydrich M. Aislamiento y caracterización de cepas de Bacillus asociadas al cultivo del arroz (Oryza sativa L.). Rev Bras de Agroecología. 2011;6(1):90-99.
30.Reinhold-Hurek B, Maes T, Gemmer S, Van Montegu M, Hurek T. An endoglucanse is involved in infection of rice roots by the not-cellulose-metabolizing endophyte Azoarcus sp. strain BH72. Mol Plant Microbe Interact. 2006;19:181-188.
31.Feng Y, Shen D, Song W. Rice endophyte Pantoea agglomerans YS19 promotes host plant growth and affects allocations of host photosynthates. J Appl Microbiol. 2006;100:938-945.
32.Jha B, Thakur MC, Gontia I, Albrecht V, Stoffels M, et al. Isolation, partial identification and application of diazotrophic rhizobacteria from traditional indian rice cultivars. European J of Soil Biology. 2009;45:62-72.
33.Watanabe I, Roger PA. Nitrogen fixation in wetland rice field. In: Subba-Rao, N.S. (ed.) Current developments in biological nitrogen fixation. Oxford & IBH Publishing Company, New Delhi, India. p. 237-276, 1984.
34.Rives-Rodríguez N, Baldani V, Hernández-Rodríguez A. Aislamiento e identificación de bacterias diazotróficas asociadas al cultivo del arroz (Oryza sativa L.). Revista Cubana del Arroz. 2012;14(2):19-25.
35.You ID, Fijii T, Sano Y, Komagata K, Yoneyama T, et al. Dinitrogen fixation of rice Klebsiella associations. CropScience. 1986;26:297-301.
36.Gillis M, Van TV, Bardin R, Goor M, Hebbar P, Willems A, et al. Polyphasic taxonomy in the genus Burkholderia leading to an amended description of the genus and proposition of Burkholderia vietnamiensis sp. nov. for N2-fixing isolates from rice in Vietnam. International Journal of Systematic Bacteriology. 1995;45:274-289.
37.Verma SC, Singh A, Chowdhury SP, Tripathi AK. Endophytic colonization ability of two deep-water rice endophytes, Pantoea sp. and Ochrobactrum sp. using green fluorescent protein reporter. Biotechnology Letter. 2004;26:425-429.
38.Korhonen TK, Laakso T, Roenkkoe R, Haahtela A. In: Hattori I, Maruyama M. (Ed). Recent advances in microbial ecology. Japan Scientific Societies Press, Tokio, Japan. p. 192-195.
39.Muthukumarasamy R, Cleenwerck I, Revathi G, Vadivelu M, Janssens D, Hoste B, et al. Natural association of Gluconacetobacter diazotrophicus and diazotrophic Acetobacter peroxydans with wetland rice. System Appl Microbiol. 2005;28:277-286.
40.Prakamhang J, Minamisawa K, Teamtaisong K, Boonkerd N, Teaumroong N. The communities of endophytic diazotrophic bacteria in cultivated rice (Oryza sativa L.). Applied Soil Ecology. 2009;42:141-149.
41.Dibut B, Martínez-Viera R, Ortega M, Ríos Y, Tejeda G, et al. Situación actual y perspectiva de las relaciones endófitas planta-bacteria. Estudio de caso Gluconacetobacter diazotrophicus-cultivos de importancia económica. Cultivos Tropicales. 2009;30(4):16-23.
42.Hernández-Rodríguez A, Heydrich M, Diallo B, El Jaziri M, Vandaputte OM. Cell-free culture medium of Burkholderia cepacia improves seed germination and seedling growth in maize (Zea mays) and rice (Oryza sativa). Plant Growth Regul. 2010;60:191-197.
43.Hernández-Rodríguez A, Heydrich-Pérez M, Acebo-Guerrero Y, Velázquez del Valle MG, Hernández-Lauzardo AN. Antagonistic activity of Cuban native rhizobacteria against Fusarium verticillioides (Sacc.) Nirenb. in maize (Zea mays L.). Applied Soil Ecology. 2008;39:180-186.
44.De Souza-Moreira FM, Da Silva K, Abrahaõ-Nobrega SR, Carvalho F. Bactérias diazotróficas associativas: diversidade, ecología e potencial de aplicações. Comunicata Scientiae. 2010;1(2):74-99.
45.Baldani VLD, Baldani JI, Döbereiner J. Inoculation of rice plants with the endophytic diazotrophs Herbaspirillum seropedicae and Burkholderia spp. Biology and Fertility Soils. 2000;30:485-491.
46.Kenendy IR, Choudhury ATMA, Kecskés ML. Non-symbiotic bacterial diazotrophs in crop-farming systems: can their potential for plant growth promotion be better exploited? Soil Biology and Biochemestry. 2004;36:1229-1244.
47.Khan AA, Jilani G, Akhtar MS, Naqvi SSM, Raheed M. Phosphorus Solubilizing Bacteria: Ocurrence, Mechanisms and their Role in Crop Production. J Agric Biol Sci. 2009;1(1):48-58.
48.Khan MS, Zaidi A, Wani PA. Role of phosphate - solubilizing microorganisms in sustainable agricultura. A review. Agron Sustain Dev. 2007;27:29-43.
49.Khan AA, Jilani G, Akhtar MS, Naqui M, Rasheed M. Phosphorus Solubilizing Bacteria: occurrence, Mechanisms and their Role in Crop Production. J Agric Biol SCI. 2009;1(1):48-58.
50.Bashan Y, Kamnev AA, de-Bashan LE. A proposal for isolating and testing phosphate-solubilizing bacteria that enhance plant growth. Biol Fertil Soil. 2013;49:1-2.
51.García de Salamone IE, Salvo LP, Escobar Ortega JS, Boa Sorte PMF, et al. Field response of rice paddy crop to inoculation with Azospirillum: physiology of rhizosphere bacterial communities and the genetic diversity of endophytic bacteria in different parts of the plants. Plant and Soil. 2010;336(1-2):351-362.
52.Höfte M, Altier N. Fluorescent pseudomonads as biocontrol agents for sustainable agricultural systems. Research in Microbiology. 2010;161:464-471.
53.Faraldo-Gómez JD. Protein-mediated Siderophore Uptake in Gram-negative Bacteria: A Structural Perspective. Springer-Verlag Berlin Heidelberg, Berlin. 2007.
54.Raaijmakers JM, Bruijn ID, Nybroe O, Ongena M. Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol Rev. 2010;34:1037-1062.
55.Selin C, Habibian R, Poritsanos N, Athukorala SNP, Fernando D, de Kievit TR. Phenazines are not essential for Pseudomonas chlororaphis PA23 biocontrol of Sclerotinia sclerotiorum, but do play a role in biofilm formation. FEMS Microbiol Ecol. 2010;71:73-83.
Recibido: 5-7-2013.
Aceptado: 8-11-2013.
*Autor para la correspondencia: Annia Hernández-Rodríguez. Departamento de Microbiología y Virología, Facultad de Biología, Universidad de La Habana, Calle 25 # 455 e/ J e I, Vedado, La Habana, Cuba. Correo electrónico: annia@fbio.uh.cu.

{kind=link}
{kind=link}
{kind=link}