










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752
Rev. Protección Veg. vol.29 no.1 La Habana ene.-abr. 2014
ARTÍCULO ORIGINAL
Diversidad genética de Trichoderma spp. en Venezuela, determinada mediante análisis combinado ITS-AFLP
Genetic diversity determined by its-aflp analysis of Trichoderma spp. in Venezuela
Daynet SosaI*, Simón Pérez MartínezI, Sandy MolinaI, Jhonny DemeyI, Kelly GómezI, Diamarys DomínguezI, María IstúrizI, Raisa RumbosII, Dercy ParraIII
IInstituto de Estudios Avanzados (IDEA). Carretera Nacional Hoyo de la Puerta-Baruta. Valle de Sartenejal. Baruta. Caracas. Edo. Miranda, Venezuela.
IIInstituto Nacional de Investigaciones Agrícolas (INIA). Estación Local Chama. Km 41 vía Santa Bárbara del Zulia al Vigía. El Vigía. Estado Zulia, Venezuela.
IIIInstituto Nacional de Investigaciones Agrícolas (INIA). Estación Experimental Padrón, Caucagua, Estado Miranda, Venezuela.
RESUMEN
Se estudió la diversidad genética entre 68 aislados de Trichoderma procedentes de diferentes cultivos, sustratos y localidades, mediante los análisis ITS-AFLP. Se secuenció la región ITS1-5.8S-ITS2 del ADN ribosomal amplificada con los cebadores ITS1 e ITS4 y en el caso de los AFLP se utilizaron cuatro combinaciones de oligonucleótidos. Las relaciones genéticas entre los aislados se analizaron mediante el uso combinado del Análisis de Coordenadas Principales, el Análisis de Conglomerados y el ajuste de un Biplot Logístico Externo sobre datos de disimilitud, utilizando los coeficientes de Jaccard, Emparejamiento simple, Dice y Rogers y Tanimoto. Se identificaron nueve especies y las más abundantes fueron Hypocrea lixii (anamorfoTrichoderma harzianum) y T. koningiopsis representadas por 22 y 20 aislados, respectivamente. Le siguen Hypocrea virens (anamorfo T. virens), Trichoderma ghanense; Trichoderma asperellum y Trichoderma brevicompactum con 7, 6, 4 y 4 aislados respectivamente. Las especies menos frecuentes fueron Trichoderma erinaceum con dos aislados y Trichoderma spirale y Trichoderma longibrachiatum con un aislado cada una. Los AFLP formaron cuatro grupos, correspondiendo uno de ellos al 99,52 % de los aislados de Trichoderma asperellum, otros dos agruparon al 85,54 y 50% de H virens y H. lixii, respectivamente. Las otras seis especies se ubicaron el cuarto grupo, y no pudieron ser diferenciadas entre si. El iniciador con mayor contenido de Información Polimórfica fue AG+CAG, que además permitió separar la especie H. lixii del resto. La combinación AG+CAG separó a T. asperellum de las otras especies.
Palabras clave: Trichoderma, ITS, AFLP, polimorfismo, control biológico.
ABSTRACT
The genetic diversity among 68 Trichoderma isolates from different cultivars, substrates and localities was established by ITS-AFLP analysis. The ribosomal DNA region ITS1-5.8S-ITS2 was sequenced and amplified with the primers ITS1 and ITS4. In the case of AFLP, four oligonucleotide combinations were used. The genetic relationships between isolates were analyzed by the combined use of Principal Component Analysis, Cluster analysis, and the adjustment of an External Logistic Biplot over dissimilarity data using Jaccard, simple coupling, Dice and Rogers, and Tanimoto coefficients. Nine species were identified, and the most abundant were Hypocrea lixii (anamorf Trichoderma harzianum) and T. koningiopsis, with 22 and 20 isolates respectively. They were followed by Hypocrea virens (anamorf T. virens), Trichoderma ghanense; Trichoderma asperellum and Trichoderma brevicompactum, with 7, 6, 4 and 4 isolates, respectively. The least frequent species were Trichoderma erinaceum, Trichoderma spirale and Trichoderma longibrachiatum (2, 1 and 1 isolates, respectively). All the isolates clustered in four AFLP groups, with one containing 99,52% of T. asperellum, and the others with 85,54% and 50% of H. virens and H. lixii, respectively. The other six species grouped together in the four group, and were indistinguishable from one another. The most informative initiator was AG+CAG showing the greatest content of polymorphic information, and which additionally allowed discriminating H. lixii from the other isolates. AG+CAG combination clearly separated T. asperellum from the other species.
Key words: Trichoderma, ITS, AFLP, polymorphism, biological control.
INTRODUCCIÓN
El control biológico de plagas es una alternativa ecológicamente limpia y compatible con diferentes modelos de agricultura. En este sentido, las especies del género Trichoderma (teleomorfo: género Hypocrea) son los antagonistas más utilizados como agentes de control biológico, debido a su adaptabilidad a varias condiciones ecológicas (1), así como a su habilidad antagónica ancestral, conocida como hiperparasitismo necrotrófico o micoparasitismo (2), contra hongos patógenos de plantas. Las especies del género, además, muestran un alto nivel de diversidad genética y pueden ser utilizadas para producir una amplia gama de productos de interés comercial y ecológico (3,4).
Los marcadores moleculares basados en la Reacción en Cadena de la Polimerasa (PCR, por sus siglas en inglés), permiten explorar diferentes regiones del genoma para inferir la estructura genética de las poblaciones, y su asociación con características significativas desde el punto de vista agronómico. Estos se basan en el grado de polimorfismo presente naturalmente en el material genético de los organismos y se emplean, entre otras, para estudiar la diversidad genética de los mismos, a nivel intra e interespecífico. Teniendo en cuenta que Trichoderma / Hypocrea es un género muy diverso, y que la identificación de cepas a nivel de especie mediante caracteres morfológicos es difícil y confusa, debido a la complejidad del género (3); el empleo de los marcadores moleculares y el análisis de secuencias nucleotídicas específicas, incrementaron las facilidades para la correcta ubicación filogenética, identificación y caracterización de la diversidad de las especies que lo conforman (1, 5, 6, 7, 8, 9, 10).
El polimorfismo de la longitud de los fragmentos amplificados (AFLP) es un método de genotipado, basado en la amplificación selectiva de un subconjunto de fragmentos de ADN generados por la digestión del mismo con enzimas de restricción (11). Para una amplia variedad de taxones, incluyendo plantas, hongos y bacterias, los marcadores AFLP permitieron detectar especies muy relacionadas o crípticas; siendo una técnica altamente discriminatoria y reproducible, utilizada extensivamente para el estudio de muchos microorganismos (12), facilitando la compilación de patrones AFLP de referencia estándar en bases de datos de diversidad, lo que a su vez permite armonizar el diagnóstico de microorganismos a escala mundial.
En Venezuela, estudios previos indicaron la presencia de seis especies de Trichoderma en 38 aislados de la rizósfera de cacao del estado Carabobo en Venezuela, siendo Trichoderma harzianum Rifai la más abundante (13), no existiendo otros reportes hasta el momento.
El objetivo del presente estudio fue determinar la diversidad genética de aislados de Trichoderma identificados mediante PCR ITS y procedentes de diferentes cultivos, sustratos y estados, con el empleo de marcadores moleculares de tipo AFLP.
MATERIALES Y MÉTODOS
Colecta de muestras
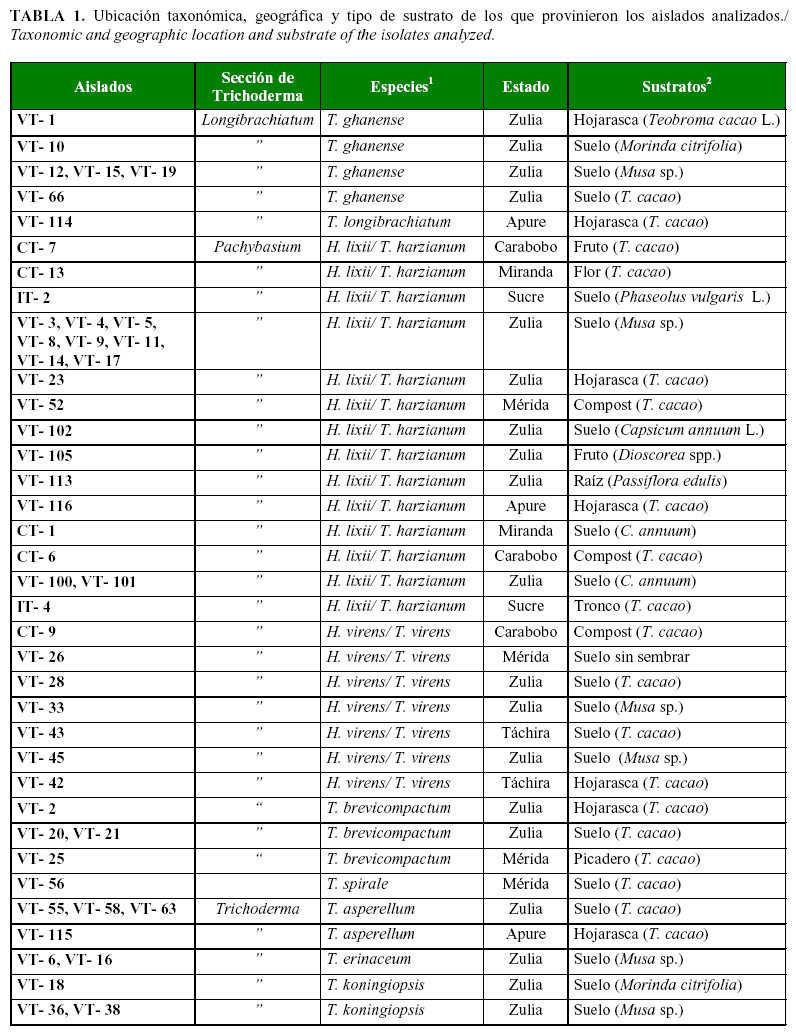
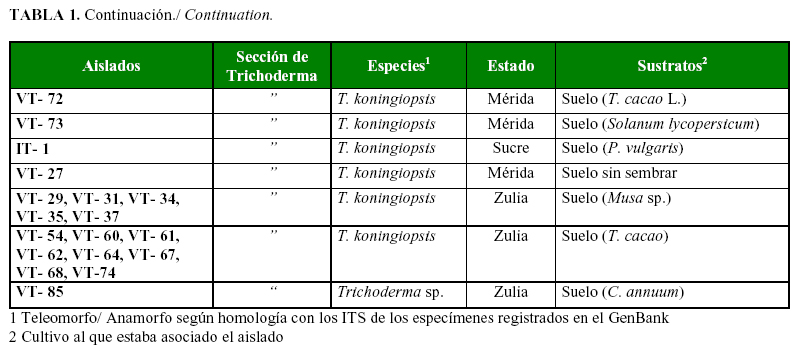
Se realizaron colectas en tres regiones cacaoteras de Venezuela: la región centro-norte-costera, la nor-oriental y la sur-occidental; y se tomaron muestras de suelo, hojarasca, cultivos asociados y composteros a base de cacao (Tabla 1a, 1b).
El muestreo en las parcelas fue no probabilístico, a conveniencia. El posible sesgo sistemático debido al muestro de conveniencia, fue corregido aleatorizando la selección de plantas dentro de cada parcela donde se recogieron las muestras (14).
Aislamiento e identificación morfológica hasta género
Se trituraron 10 g de suelo seco a temperatura ambiente por cada muestra y se agitaron por 30 min en 90 ml de agua destilada estéril. Luego, con un asa de platino estéril, se estrió una alícuota en cápsulas Petri con medio papa dextrosa agar (PDA). Se hicieron dos repeticiones por muestra. A los cinco días de incubación a temperatura ambiente (25±2oC) se purificaron las colonias desarrolladas que mostraron similitud morfológica con Trichoderma.
Identificación de especies por ITS
Para la extracción de ADN a partir de micelio, los aislados se cultivaron en medio líquido Czapek (10 días/oscuridad/ 24±2oC con agitación en zaranda a 250rpm durante 36 a 48 horas). La masa micelial se colectó y secó con papel de filtro Whatman Nº 1 estéril y se preservó en papel aluminio a -20°C hasta el momento de la extracción de ADN (15). La cantidad de ADN total extraído se cuantificó en un espectrofotómetro Eppendorf BioPhotometer y se corroboró su calidad por medio de electroforesis en gel de agarosa al 1%, con buffer de corrida TBE 1X. El mismo se guardó en buffer de suspensión a base de Tris 10mM, pH 8 hasta su utilización.
Amplificación de la región ITS. PCR-ITS. Se amplificó por PCR la región ITS1-5.8S-ITS2 del ADN ribosomal con los cebadores ITS1 e ITS4 descritos por White et al. (16). Para cada reacción de PCR se utilizó un volumen final de 40µl que contuvo Buffer PCR 1X, MgCl2 (1,5mM), dNTPs (0,2mM); 0,125uM de cada cebador, ADN (50ng) y 1U de la enzima GoTaq® Flexi Polimerasa (Promega). Se utilizó un termociclador BIO-RAD (iCycler) de acuerdo al programa descrito por Hermosa et al. (17).
Para visualizar los fragmentos amplificados, se tomaron 10ul de PCR y se mezclaron con buffer de carga, el cual contiene Azul de bromofenol (1mg/ml), glicerol 50% y Syber Green 0,2%; se cargaron en un gel de agarosa al 1% y se realizó una electroforesis horizontal, en buffer TBE 1X (22,5 mM Tris base; 22,5mM H3BO3; 0,5mM EDTA) pH 8.3 a 80V durante una hora. Las bandas obtenidas se compararon con el marcador de 1Kb plus (Invitrogen). Los geles se visualizaron en un digitalizador de imágenes tipo Typhoon 9410 (Amersham).
Secuenciación
Los productos amplificados de la región ITS fueron purificados con el Kit de purificación AccuPrep®PCR marca BioNEER para su posterior secuenciación con el Kit BigDye®Terminator V3.1. Todas las secuencias nucleotídicas obtenidas se procesaron con los programas Chromas Pro versión 1.41 y BioEdit (Sequence Alignment Editor), con el fin de generar los consensos de las cadenas hacia adelante y hacia atrás para su comparación. Las secuencias obtenidas en este estudio se compararon con las disponibles en el registro nucleotídico del Genbank (base de datos del National Center for Biotechnology Information, (NCBI)) utilizando la aplicación Basic Local Alignment Search Tool (BLAST) para identificar los aislados con aquellos de mayor similitud; depositándose las mismas en el Genbank con posterioridad.
Diversidad genética
AFLP. Los fragmentos AFLP se obtuvieron según el protocolo clásico (11). Consistió en la digestión del ADN con las enzimas de restricción EcoRI y MseI, la ligación de los adaptadores específicos a los sitios de restricción de estas endonucleasas, una primera amplificación pre-selectiva por PCR con cola adenina (A) y una segunda amplificación con cuatro combinaciones de oligonucleótidos: E-AG/M-CAA, E-AA/M-CAC, E-AG/M-CAG y E-AG/M-CAC. Los productos amplificados se separaron por electroforesis en geles de poliacrilamida al 6%, con buffer TBE 1X, a 65W durante 1,5h. Los oligonucleotidos se marcaron con los fluoróforos FAM y Cy5 sin realizar tinción; los geles se visualizaron en un digitalizador de imágenes Typhoon 9410 (Amersham).
Análisis de los datos. En ausencia de análisis de segregación no se hizo ningún supuesto sobre la naturaleza genética de los alelos. Los fragmentos de amplificación se codificaron de acuerdo a un marcador dominante, es decir 1 (banda presente) y 0 (banda ausente), generando una columna por locus para cada iniciador. El nivel de polimorfismo y la capacidad discriminatoria de cada iniciador se valoró a través del contenido de información polimórfica (PIC, siglas en inglés) y la probabilidad de obtener parejas idénticas de alelos entre las muestras estudiadas (18,19). La relación genética entre los 68 aislados empleados se estudió aplicando una estrategia metodológica propuesta (20), que planteó el uso combinado del Análisis de Coordenadas Principales, el Análisis de Conglomerados y el ajuste de un Biplot Logístico Externo, sobre datos de disimilitud utilizando los coeficientes de Jaccard, Emparejamiento simple, Dice y Rogers y Tanimoto (21). El número k de dimensiones a ser retenidas, el coeficiente de similitud que mejor define la estructura de los datos y las medidas de la calidad, fueron calculados utilizando también los procedimientos descritos por Demey et al. (20). Para el cálculo de la capacidad informativa y discriminatoria de los iniciadores se utilizó Info-Gen versión 2009p. Para los cálculos y representaciones gráficas de los procedimientos estadísticos se utilizaron se han usado un conjunto de rutinas desarrolladas bajo MatLab versión 2009a (The MathWorks Inc, 2009) (http://www.biplot.usal.es).
RESULTADOS Y DISCUSIÓN
Los datos moleculares, cada vez más disponibles, de los hongos con potencialidad para el control biológico mostraron la importancia de los estudios genéticos en la investigación, desarrollo y registro de estos agentes de control biológico, donde los estudios de diversidad resultan imprescindibles para el uso seguro y fiable de los mismos. En el ámbito de la investigación en el género Trichoderma se formula con frecuencia la interrogante acerca de si la capacidad para el control biológico de este género, es una propiedad general o un atributo específico de un número limitado de cepas. Debido a ello, se hace indispensable el estudio de la naturaleza y diversidad de todo posible agente de control biológico, considerando que el alto nivel de diversidad genética de este género puede ser utilizado en una amplia gama de productos de interés comercial y ecológico (3, 4).
Las secuencias completas de las regiones ITS1-5·8S-ITS2, junto con las secciones terminales de las subunidades pequeña y grande de los genes ribosomales, se obtuvieron para todos los aislados. Se amplificaron fragmentos con 600-650 pares de bases (pb) aproximadamente, conteniendo, en muchos casos, aproximadamente 22 bases del extremo final del gen 18S y de 15 a 40 bases del extremo inicial del gen 28S. El tamaño de Ia región amplificada obtenido concuerda con los datos obtenidos por otros autores para Trichoderma (22, 23, 24).
En BLAST se confirmó la identificación de nueve especies, a partir de las secuencias de los 68 aislados del género Trichoderma. Se identificaron 22 aislados como Hypocrea lixii/ T. harzianum Rifai; Trichoderma koningiopsis (Samuels, C. Suárez & H.C. Evans) con 20; Hypocrea virens/ Trichoderma. virens (Miller, Giddens & Foster) con 7; Trichoderma ganense (Yoshim. Doi, Y. Abe & Sugiy ) con 6 y Trichoderma asperellum Samuels, Lieckf. & Nirenberg y Trichoderma brevicompactum Kraus, Kubicek & Gams representadas por cuatro aislados cada una; Trichoderma erinaceum (Bissett, C.P. Kubicek & Szakacs) con 2 y Trichoderma spirale Bissett y Trichoderma longibrachiatum Rifai con un aislado cada una, más un aislado (VT-85) no identificado Trichoderma sp.
La obtención de porcentajes altos para: I) de identidad (dado por la similitud que existe entre la secuencia que se está evaluando y la secuencia que se encuentra en la base de datos del GenBank) y II) de cobertura (correlación que existe entre el tamaño de la secuencia que se encuentra en el GenBank y la secuencia que se está estudiando), indicaron el grado de confiabilidad en la certeza de que los aislados fueron correctamente identificados.
Se produjo un total de 178 fragmentos polimórficos, que representan aproximadamente el 99% del total de fragmentos obtenidos, estando entre 38 y 51 el número de fragmentos por iniciador (Tabla 2). El iniciador más informativo fue el AG+CAG con un Contenido de Información Polimórfica cercano al 50% del intervalo teórico de 0,01 a 0,50; el que además mostró la mayor capacidad discriminatoria. Utilizando los cuatro iniciadores, la probabilidad media de que dos aislados diferentes tengan igual identidad, fue de 4,9x10-41, lo que indicó un alto grado de confianza en el conjunto de iniciadores seleccionados, dado que la región del genoma explorada permite la separación de hasta 1041 aislados de manera simultánea (Tabla 2).
De Souza et al. (28) utilizaron AFLP para analizar la diversidad genética de 91 aislados de T. stromaticum de cacao de las regiones del amazonas de Brasil, Colombia y Ecuador, y obtuvieron 144 bandas polimórficas, logrando separar dos grupos dentro de la población de esta especie. Otros estudios revelaron que, con el empleo de 20 combinaciones de cebadores AFLP, se obtuvieron un total de 1127 loci polimórficos (23), y no se encontró relación clara entre los agrupamientos del dendrograma y el origen geográfico de los aislados (5, 23).
Por otra parte, en el estudio de 48 aislados que representan seis especies de Trichoderma de diferentes zonas de cultivo de maní, se obtuvieron 234 bandas polimórficas con seis combinaciones de cebadores (27); mientras que Larralde et al. (26) obtuvieron 75 bandas polimórficas utilizando cuatro combinaciones de cebadores solamente en nueve aislados de seis especies de Trichoderma.
La distribución de los coeficientes de correlación lineal de Pearson entre los n(n-1)/2 elementos distintos fuera de la diagonal de las matrices de distancias observada y estimada , para distintas combinaciones de k-dimensiones retenidas luego de aplicar el Análisis de Coordenadas Principales, permitió seleccionar el coeficiente de similitud de Emparejamiento Simple, como el que refleja la mayor coherencia entre la matriz de distancias observadas y estimadas y garantiza la mayor estabilidad de la representación, debida a la poca dispersión de los autovalores. Adicionalmente, con este coeficiente no es necesario retener más de dos dimensiones para extraer la mayor cantidad de información posible. En términos de similitud, esto significa que es posible probar que dos aislados con posiciones más cercanas en la representación bidimensional, tendrán patrones más similares de ADN respecto a las secuencias utilizadas (20).
La Figura 1a muestra las relaciones genéticas entre los 68 aislados, basada en la disimilaridad debido al coeficiente de Emparejamiento simple y los cuatro iniciadores AFLP. El espacio bidimensional obtenido del Análisis de Coordenadas Principales, explica el 37,62% de la variabilidad total. Bajo el algoritmo UPGMA, utilizando las dos primeras coordenadas principales retenidas, se forman cuatro grupos de aislados demarcados por el diagrama de Voronoi y se representan los fragmentos de amplificación que definen estos grupos seleccionados después del ajuste del Biplot Logístico Externo y corregida por el p-valor, Bonferroni y el pseudo R2 de Nagelkerke/Cragg & Uhler's.
Los grupos AFLP (Figura 1a) mostraron una diversidad genética media entre 0,421-0,426, especificada de la siguiente manera: el grupo 1 formado por H. virens, 0,427; el grupo 4 de T. asperellum, 0,425; y el grupo 3 de H. lixii , 0,421. Particularmente en estas especies pertenecientes a las secciones Pachybasium y Trichoderma se ha observado una alta variabilidad intra e interespecífica a partir de datos fisiológicos, bioquímicos, morfológicos y moleculares, particularmente en el complejo de especies conocido como T. harzianum /T. inhamatum (17, 5). El resto de los valores de diversidad se dividieron en media y baja. Las especies T. spirale (0,419), T. koningiopsis (0,419), T. brevicompactum (0,415) y T. erinaceum y T. ghanense (0,414) formaron el de diversidad media. Con los menores valores de diversidad estuvieron T. ghanense (0,397) y T. longibrachiatum (0,389). Esta estrategia metodológica permite estudiar en la misma representación no sólo la relación entre los individuos como en el Análisis de Conglomerados clásico, sino que además permite estudiar la relación entre individuo-alelo y alelo-alelo (20).
La Figura 1b muestra la superposición de la clasificación generada del análisis de las secuencias de los espaciadores internos transcritos del ADNr sobre la generada por los cuatro iniciadores AFLPs. Los iniciadores AFLP preservaron en más del 85% la estructura de grupos esperada para H. virens y en T. asperellum fue del 99,52%, alrededor del 50% para H. lixii. El resto de los aislados formó un grupo mixto con las otras seis especies identificadas por ITS. Estos resultados sugieren que, estos iniciadores son buenos para una primera clasificación a nivel de género, pero no son sensibles para separar especies dentro del género, a diferencia de lo obtenido con la secuenciación del ITS.
Los AFLP se muestran aquí como una herramienta para la obtención de las huellas genéticas de estos aislados y para realizar estudios futuros de las relaciones entre los mismos a nivel de población y especie. Sin embargo, el análisis basado en las secuencias (genealogías de los genes) puede ser más útil para definir las relaciones filogenéticas entre los clados. Como ambos enfoques representan procesos evolutivos independientes, la complementación de los datos de las secuencias con los datos de los AFLP puede proporcionar medios más sólidos para caracterizar nuevas cepas, aspecto esencial en el desarrollo de agentes de control biológico.
El análisis de los iniciadores mostró la proyección para cada fragmento de amplificación seleccionado después del ajuste del Biplot Logístico Externo y su relación con la formación de grupos. La Figura 2 muestra que el iniciador AG+CAA permitió separar la especie H. lixii del resto y el iniciador AG+CAG a la especie T. asperellum del resto.
Los iniciadores AFLP analizados indicaron que es posible su empleo para la identificación rutinaria a nivel de género de las especies de Trichoderma, lo que sumado a la secuenciación de regiones conservadas, permitió la identificación inequívoca de las mismas, tal como señalaron Agrawal y Kotasthane (29). Alternativamente, se pueden analizar un mayor número de combinaciones de iniciadores a efectos de identificar las especies más importantes.
En una investigación previa en Venezuela (13) se identificaron 6 especies de Trichoderma por las secuencias de los ITS: T. harzianum, T. virens, T. brevicompactum, T. pleurotum, T. koningiopsis y T. asperellum. En el anterior artículo se mencionan otros dos especímenes encontrados, similares a T. theobromicola y T. ovalisporum, pero la certeza en su identificación no fue explicitada por los autores. En este estudio encontramos las mismas especies que Rivas et al. (13), a excepción de T. pleurotum, y otras cuatro adicionales: T. ghanense, T. erinaceum, T. spirale y T. longibrachiatum. En Venezuela también se ha informado la presencia de T. koningii (30), aunque empleando métodos clásicos de identificación.
AGRADECIMIENTOS
Este trabajo fue financiado por el Ministerio del Poder Popular para la Ciencia, Tecnología e Innovación mediante el Proyecto en Red Ruta del Chocolate (Venezuela).
REFERENCIAS
1. Druzhinina IS, Seidl-Seiboth V, Herrera-Estrella A, Horwitz BA, Kenerley CM, Monte E, et al. Trichoderma: the genomics of opportunistic success. Nat Rev Microbiol. 2011;9:749-759.
2. Kubicek CP, Herrera-Estrella A. Comparative genome sequence analysis underscores mycoparasitism as the ancestral life style of Trichoderma. Genome Biol. 2011;12:2-15.
3. Hoyos-Carvajal L, Bissett J. Oscar Grillo (Ed.), Biodiversity of Trichoderma in neotropics, the dynamical processes of biodiversity - case studies of evolution and spatial distribution, PhD Thesis. 2011. 303-320pp.
4. Lorito M, Woo SL, Harman GE, Monte Enrique. Translational research on Trichoderma: from «omics» to the field. Annu Rev Phytopathol. 2010;48:395-417.
5. Naeimi S, Khodaparast SA, Javan-Nikkhah M, Vágvölgyi C, Kredics L. Species pattern and phylogenetic relationships of Trichoderma strains in rice fields of Southern Caspian sea, Iran. Cereal Research Communications 2011;39:560-568.
6. Xia X, Lie TK, Qian X, Zhonghui Z, Yaojian H, Yuemao S. Species diversity, distribution, and genetic structure of endophytic and epiphytic Trichoderma associated with banana roots. Microb Ecol. 2011;61:619-625.
7. Srivastava S, Singh V, Gupta PS. Ribosomal DNA sequence based characterization of Trichoderma isolates antagonistic to Colletotrichum falcatum causing red rot disease of sugarcane. Sugar Tech. 2011;13:245-249.
8. Samuels GJ, de Souza Ismaiel A, Chaverri P. Trichoderma stromaticum and its overseas relatives. Mycol Progress. 2012;11:215-254.
9. Maymon M, Minz D, Barbul O, Zveibil A, Elad Y, Freeman S. Identification of Trichoderma Biocontrol Isolates to Clades According to ap-PCR and ITS Sequence Analyses. Phytoparasitica. 2004;32:370-375.
10.Druzhinina IS, Kopchinskiy AG, Komoj M, Bissett J, Szakacs G, Kubicek CP. An oligonucleotide barcode for species identification in Trichoderma and Hypocrea. Fungal Genetics and Biology. 2005;42:813-828.
11.Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T, Hornes M, et al. AFLP: a new technique for DNA fingerprinting. Nucleic Acids Res. 1995;23:4407-4414.
12.Savelkoul PHM, Aarts HJM, de Haas J, Dijkshoorn L, Duim B, Otsen M, et al. Amplified-fragment length polymorphism analysis: the state of an art. J Clin Microbiol. 1999;37:3083-3091.
13.Rivas M, Pavone D. Diversidad de Trichoderma spp. en plantaciones de Theobroma cacao L. del estado Carabobo, Venezuela, y su capacidad biocontroladora sobre Crinipellis perniciosa (Stahel) Singer. Interciencia. 2010;35:777-783.
14.Levy PS, Lemeshow S. Sampling of Populations: Methods and Applications. Wiley Series in Survey Methodology. 2008. (4th ed).
15.Naranjo L, Urbina H, De Sisto A, Leon V. Isolation of autochthonous non-white rot fungi with potencial for enzymatic upgrading of venezuelan extra-heavy crude oil. Biocatalysis and Biotransformation. 2007;25:341-349.
16.White TJ, Bruns T, Lee S, Taylor J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In Innis MA, Gellfand DH, Sninsky JJ & White TJ. (eds.). PCR Protocols: a guide to methods and applications. Academic Press, Inc., New York. 1990. 315-322 pp.
17.Hermosa MR, Keck E, Chamorro I, Rubio B, Sanz L, Vizcaìno JA, et al. Genetic diversity shown in Trichoderma biocontrol isolates. Mycol Res. 2004;108:897-906.
18.Anderson JA, Churchill GA, Autrique JE, Tanksley SD, Sorrells ME. Optimizing parental selection for genetic linkage maps. Genome. 1993;36:181-186.
19.Demey JR, Zambrano AY, Fuenmayor F, Segovia V. Relación entre las características moleculares y morfológicas en una colección de yuca. Interciencia. 2003;28:684-689.
20.Demey JR, Vicente-Villardón JL, Galindo-Villardón MP, Zambrano AY. Identifying molecular markers associated with classification of genotypes by external logistic biplots. Bioinformatics. 2008;24:2832-2838.
21.Sneath PHA, Sokal RR. Numerical taxonomy: The principles and practice of numerical classification. Freeman W.H. and Co. San Francisco. USA. 1973; 573pp.
22.Ospina-Giraldo MD, Royse DJ, Chen X, Romaine CP. Molecular phylogenetic analyses of biological control strains of Trichoderma harzianum and other biotypes of Trichoderma spp. associated with mushroom green mold. Phytopathology. 1999;89:308-313.
23.Vahabi K, Sharifnabi B, Zafari D. Genetic diversity of Trichoderma spp. associated with button mushroom Agaricus bisporus, inferred from AFLP markers and ITS sequencing. Acta Phytopathologica et Entomologica Hungarica. 2009;44:239-253.
24.Hermosa MR, Grondona I, Iturriaga EA, Diaz-Minguez JM, Castro C, Monte E. Molecular characterization and identification of biocontrol isolates of Trichoderma species. Applied and Environmental Microbiology. 2000;66:1890-1898.
25.De Souza JT, Pomella AWV, Bowers JH, Pirovani CP, Loguercio LL, Hebbark KP. Genetic and biological diversity of Trichoderma stromaticum, a mycoparasite of the cacao witches'-broom pathogen. Phytopathology. 2006;96:61-67.
26.Buhariwalla HK, Srilakshmi P, Kannan S, et al. AFLP Analysis of Trichoderma spp. from India Compared with Sequence and Morphological-based Diagnostics. Journal of Phytopathology. 2005;153:389-400.
27.Larralde CP, Santiago MR, Sifuentes AM, Rodríguez IC, Rodríguez MA, et al. Biocontrol potential and polyphasic characterization of novel native Trichoderma strains against Macrophomina phaseolina isolated from sorghum and common bean. Appl Microbiol Biotechnol. 2008;80:167-177.
28.Agrawal T, Kotasthane AS. Mycoparasitism, AFLP characterization and effectiveness of Trichoderma species isolated from Chhattisgarh in Central India against Rhizoctonia solani infecting rice. J Mycol Pl Pathol. 2010;40:532-539.
29.Iturriaga T, Minter DW. Hongos de Venezuela Disponible en http://www.cybertruffle.org.uk/venefung/index.htm [sitio internet, versión 1.00] (2006).
Recibido: 31-1-2013.
Aceptado: 29-7-2013.
*Autor para la correspondencia: Daynet Sosa. Correo electrónico: daynet.sosa@gmail.com.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}