










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.32 no.2 La Habana may.-ago. 2017
ARTÍCULO ORIGINAL
Desarrollo, reproducción y tablas de vida de Nesidiocoris tenuis Reuter empleando como presa estadios inmaduros de mosca blanca
Development, reproduction and life tables of Nesidiocoris tenuis Reuter using inmature stages of whitefly as preyabstract
Heyker L. Baños-Díaz 1*, Tay Ruiz-Gil2, Marbely del Toro-Benítez1, Ileana Mirada-Cabrera1 y María de los A. Martínez-Rivero1
1 Laboratorio de Entomología -Acarología. Dirección de Sanidad Vegetal. Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana. Cuba.
2 Facultad de Biología. Universidad de la Habana (UH), La Habana, Cuba.
RESUMEN
Se determinaron la duración del ciclo de vida y los parámetros ecológicos en condiciones de laboratorio del depredador Nesidiocoris tenuis Reuter a temperatura de 26,7± 2°C, 66,39± 10% HR y fotoperiodo 10:14. Como substrato se utilizaron plantas de tomate, Solanum lycopersicum L. var. L43 y, como alimento, huevos y ninfas de Bemisia tabaci Gennadius biotipo B. El tiempo de desarrollo para huevos y ninfas fue de 11 y 27 días, respectivamente, y la mortalidad de los estadios inmaduros de 28 %. Las hembras mostraron una longevidad de 13 días, con una relación entre sexos de 0,56. La tasa neta de reproducción del depredador fue de Ro= 23,8; la tasa intrínseca de incremento fue rm= 0,111 día−1; mientras que la tasa finita de crecimiento fue de λ=1,12, con un tiempo medio generacional de T=28,7 días y el tiempo de duplicación de TD=6,3 días.
Palabras clave: Bemisia tabaci; tabla de vida; mosca blanca; N. tenuis.
ABSTRACT
The life cycle duration and the ecological parameters of the predator Nesidiocoris tenuis Reuter were determined under laboratory conditions at 26.7 ± 2 ° C, 66.39 ± 10% RH and photoperiod 10:14. For this purpose, tomato plants, Solanum lycopersicum L. var. L43, were used as substrate and eggs and nymphs of Bemisia tabaciGennadius biotype B as food. The development time for eggs and nymphs of this predator were 11 and 27 days, respectively. The mortality of the immature stages was 28%, the longevity of females 13 days and the sex ratio 0, 56. Other parameters were: the net reproductive rate Ro = 23, the intrinsic rate of increase rm = 0.111 day- 1, the finite growth rate λ = 1.2, the average generation time T = 28.7 days and the population doubling time TD = 6.3 days
Key words: Bemisia tabaci; Life table; White fly; N. tenuis
INTRODUCCIÓN
A nivel mundial, el abuso de los métodos químicos ha provocado graves problemas en los sistemas agrícolas (1), como son la pérdida de la biodiversidad, la contaminación de los suelos y el daño a la salud humana, además de contribuir al desarrollo de plagas más resistentes (2). Por lo que en la actualidad se trabaja en métodos menos agresivos y sustentables; algunos de ellos son el manejo del hábitat y el control biológico por conservación (3).
El control biológico por conservación implica maximizar el impacto de los enemigos naturales, proveer de recursos ecológicos claves y minimizar el uso de plaguicidas. En resumen, “mejora el ambiente” para aumentar la eficiencia de los enemigos naturales (4).
En las últimas décadas, el interés por el uso de depredadores para el control de plagas (lepidópteros, áfidos, trips y mosca blanca) en solanáceas se incrementó y se destaca el empleo de los géneros Dicyphus, Macrolophus y Nesidiocoris (5,6,7,8,9). En este último, se ha hecho especial énfasis en el uso de Nesidiocoris tenuis Reuter como enemigo natural de trips (Thrips palmi Karney) y moscas blancas (Trialeurores vaporariorum Westwood y Bemisia tabaci Gennadius (10,11), con cerca de 40 % de efectividad en condiciones de campo.
En Cuba, N. tenuis se encuentra asociado a cultivos de importancia agrícola y económica, como el tomate, el tabaco y el pimiento (7), y se aprecia reducción en las poblaciones de plagas secundarias como B. tabaci, áfidos y algunos lepidópteros, tras sus apariciones, de forma natural, tanto en cultivos protegidos como en campo abierto. Debido a la eficiencia mostrada para el control de B. tabaci en condiciones de laboratorio (12), resultó interesante conocer aspectos básicos de la biología y tablas de vida de N. tenuis usando como presa a B. tabaci, lo cual constituyó el objetivo del presente trabajo.
MATERIALES Y MÉTODOS
Los experimentos se desarrollaron entre enero y abril de 2015 en los Laboratorios de Entomología-Acarología del departamento de Plagas Agrícolas, Dirección de Sanidad Vegetal del Centro Nacional de Sanidad Agropecuaria (CENSA), Provincia Mayabeque, Cuba.
Se estableció una cría de N. tenuis a partir 250 individuos recolectados, con un aspirador, en parcelas de tomate en el Instituto de Investigaciones Hortícolas “Liliana Dimitrova” en la misma provincia, con adaptación a la metodología de Lins (13). En el laboratorio, los adultos se liberaron dentro de jaulas construidas con acrílico (dimensiones 30x30x60cm), donde se encontraban plantas de tomate (Solanum lycopersicumvar. L43) de, aproximadamente, 25 cm de altura, como sustrato de oviposición. Los insectos se alimentaron ad libitum, dos veces por semana, con estadios inmaduros de B. tabaci procedentes de una cría existente en el laboratorio.
Durante la realización de los experimentos, los valores promedios (±D.E) de la temperatura y la humedad relativa fueron de 26,7ºC (±1,3ºC) y 66,4 % (±8 %), respectivamente; se registraron utilizando un termohigrómetro digital marca Testo (EE±0,01), con un fotoperiodo natural de 10L: 14O.
Desarrollo y reproducción de N. tenuis
Para la obtención de huevos se introdujo una hembra adulta del depredador en una jaula de cría con una planta de tomate como sustrato de ovoposición. Transcurridas 24 horas, se retiró la planta con huevos y se colocó en otra jaula. Diariamente, se revisó la planta hasta confirmar la eclosión de los huevos por la presencia de ninfas.
Posteriormente, se individualizaron 50 ninfas en placas Petri de 5 cm, que contenían un disco de hoja de tomate sobre una capa de agar agua (1 %), la cual se cambió dos veces por semana.
Las ninfas se alimentaron diariamente con estadios inmaduros de la presa ad libitum y se mantuvieron bajo las condiciones ambientales antes descritas. Para determinar los cambios de los estadios se observó diariamente la presencia de exuvias y la mortalidad con un microscopio estereoscópico Zeiss KL 1500 con aumento 10x. Una vez que las ninfas pasaron al estado adulto, 15 parejas de N. tenuis, de 24 horas de edad, se individualizaron en cilindros de cristal con una planta de tomate, con 2 o 3 pares de hojas, como sustrato de oviposición. Los adultos no se alimentaron con B. tabaci biotipo B, su alimentación fue solamente fitófaga.
Los cilindros se taparon con tela de organza y se mantuvieron bajo las condiciones antes descritas por 24 horas para la oviposición. El sustrato de oviposición se cambió diariamente y a las plantas retiradas se les colocó un algodón humedecido que cubriera las raíces; estas se depositaron, individualmente, en placas Petri (15 cm), selladas con una película de PVC y se mantuvieron bajo las mismas condiciones hasta la emergencia de las ninfas.
Tabla de vida de N. tenuis
Diariamente se registraron el número de ninfas emergidas y la mortalidad de los adultos; a partir de estos datos se calcularon el periodo de oviposición (desde el día de la primera puesta hasta el día de la última puesta), la fecundidad diaria y total (número de ninfas/hembra), la longevidad de la especie y el adulto, y se construyeron las tablas de vida del depredador. Los parámetros de la tabla de vida estimados a partir de los datos se muestran en la Tabla 1.
RESULTADOS Y DISCUSIÓN
Desarrollo y reproducción de N. tenuis
El desarrollo posembrionario de N. tenuis comprendió cinco estadios ninfales, cada uno con alrededor de tres días de duración. (Tabla 2)
Los estadios 4to y 5to se alcanzaron en menos de diez días, con un ciclo de desarrollo de un mes aproximadamente, y una sobrevivencia de más del 50 % de los estadios inmaduros.
Las hembras y los machos alcanzaron el estado adulto al mismo tiempo, por lo que no existen diferencias en el periodo de desarrollo entre los dos sexos. La duración del ciclo de vida fue superior a un mes, por lo que se podrían obtener alrededor de siete generaciones anuales, con una longevidad promedio de 10 días para los adultos. La relación entre sexos estuvo a favor de las hembras (0,56), lo cual es muy conveniente, pues son las encargadas de preservar la especie, además presentan una mayor tasa de consumo.
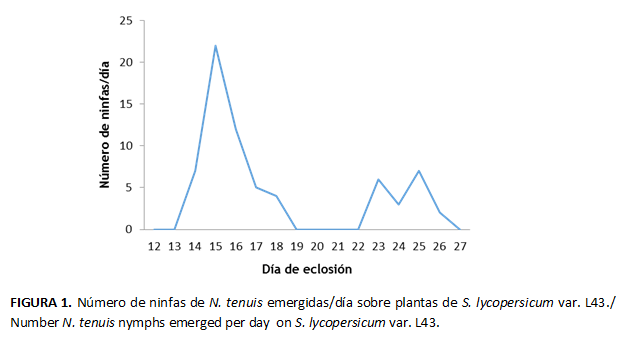
La fecundidad fue de 4,5 ± 4,8 ninfas/♀ y con 4,3 ± 5,9 ninfas/día. La curva (Fig. 1) muestra que existen tres momentos donde ocurre un mayor número de eclosiones; el primero de estos con los mayores valores de ninfas emergidas por día (23 ninfas/día), el mismo ocurre a los primeros 15 días de la oviposición. Esto se puede explicar porque es el momento en que se encuentra vivo un mayor número de hembras aptas para ovopositar.
Ocho días después ocurren dos picos, segundo y tercero, con valores superiores a la media. Ambos ocurren entre los 23 y 25 días de ovoposición. En estos momentos la sobrevivencia de las hembras disminuye hasta hacerse cero.
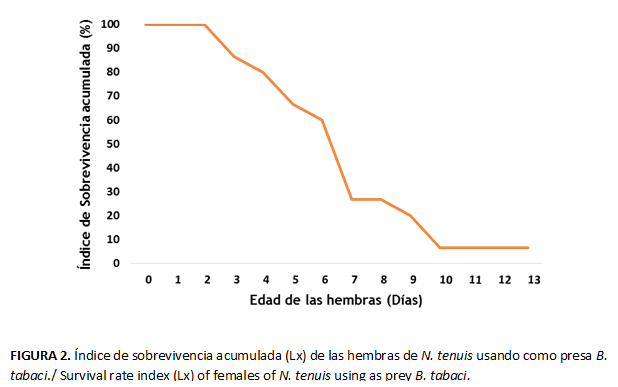
Al analizar la proporción de individuos sobrevivientes durante su tiempo de vida, los resultados mostraron una reducción significativa de la sobrevivencia a medida que aumentaba la edad de las hembras, simulando una curva de tipo II. (Fig. 2)
Sánchez et al. (14) obtuvieron un periodo embrionario de 8,9 ± 0,09 días para esta misma especie a 25ºC; mientras que para Macrolophus pygmaeus Rambur (15) y Macrolophus caliginosus Wagner (16), a igual temperatura, fue de 11,08 ± 0,19 y 11,4 ± 0,49 respectivamente. En ambos casos, los huevos de N. tenuis se desarrollaron más rápido que las especies antes mencionadas; esto pudiera estar dado por la diferencia de temperatura (2ºC), lo cual sugiere que, al parecer, a medida que aumenta la temperatura el tiempo de incubación de los huevos se hace más corto.
Según Martínez et al. (17), N. tenuis es capaz de completar su desarrollo de huevo a adulto a temperaturas constantes y mostrar una sobrevivencia superior a 80 %; se reduce así su tiempo de desarrollo a 12 días a temperaturas superiores de 30ºC. Bajo la temperatura utilizada en este estudio, el tiempo de desarrollo coincidió con el intervalo planteado por estos autores. No obstante, los resultados obtenidos por ellos deben tenerse en cuenta al realizar las crías masivas del depredador.
Según Brakefield y Kerbeke (18), existen diferencias en el desarrollo para los insectos, entre sistemas con temperaturas constantes y fluctuantes, así como que es superior la tasa de crecimiento cuando se utilizan condiciones de temperaturas controladas.
Las condiciones evaluadas durante el estudio no representan las verdaderas condiciones presentes en la naturaleza, pues los insectos se encuentran aislados de otros factores climáticos como son las lluvias o la radiación solar, entre otros. No obstante, los resultados se aproximan más a la realidad que cuando se realizan estudios bajo condiciones controladas.
Otro factor que pudo haber influido en la duración del periodo de incubación pudiera ser el tipo de presa suministrada. A los adultos no se les proporcionó presa, condición que afecta la producción de huevos, pues los nutrientes solo se pudieron obtener de la alimentación fitófaga, fuente secundaria de alimentación para estos insectos (19).
Por otro lado, en cuanto a la calidad del alimento, el contenido proteico en huevos de Ephestia kuehniella Zeller ofrecidos en otros estudios es en promedio 16,6 % de su peso fresco (20); mientras, en los huevos de B. tabaci el contenido proteico es de alrededor 10,8 % (21). Según Urbaneja et al. (22), la variedad de la planta hospedante pudiera ser otro factor que influye en el desarrollo y la sobrevivencia de N. tenuis, lo que se evidencia en los trabajos antes citados.
Además, Urbaneja et al. (6) plantearon que una adición de sacarosa (0,5 M) ad libitum a una dieta de huevos de E. kuehniella puede incrementar, significativamente, el número de descendientes de N. tenuis, sin afectar su desarrollo ninfal, aunque puede reducir el consumo de la presa. Al analizar los resultados, es posible que la no utilización de recursos azucarados pudiera ser otro factor que influyó, de forma directa, en el número de descendientes obtenidos durante el estudio. Estos resultados demuestran la necesidad del depredador de recursos azucarados en su dieta para incrementar el número de puestas por hembra. Elemento que se debe tener en cuenta durante el desarrollo de crías masivas y liberación del depredador (23).
A pesar de las diferencias antes mencionadas, Sánchez et al. (14) obtuvieron valores de relación entre sexos (0,53 para 25ºC) similares a los de este estudio (0,56) y no encontraron diferencias significativas en el tiempo de desarrollo entre hembras y machos; lo que demuestra que, quizás, las variaciones de temperatura no sean un factor que influye en la proporción de sexos de esta especie.
Tabla de vida de N. tenuis
Al estimar los parámetros de tabla de vida se obtuvo una tasa neta de reproducción (R0=23,8), definida como el número de hembras a las que da lugar cada hembra de la generación actual. El tiempo promedio entre dos generaciones sucesivas fue de 28,7 días y tiempo en el cual se duplica la generación fue de 6,3, con lo cual se ratifica el resultado descrito en la biología, con respecto al número de generaciones por año que se podrían lograr.
La tasa intrínseca de crecimiento (rm) fue de 0,11; lo que muestra la capacidad de multiplicación de la población sobre el cultivo con B. tabaci como presa. El número de individuos que se agregan a una población por día fue de 1,12, el cual se traduce como un crecimiento poblacional de un 11,2 % diario.
Los resultados de este trabajo no coincidieron con los alcanzados para otras especies pertenecientes a la misma familia, como M. pygmaeus alimentado con T. vaporariorum sobre tomate a 27,5ºC (15). En este sentido, N. tenuis tuvo una tasa neta de reproducción (R0=23,8) menor que M. pygmaeus (Ro=59,38). Sin embargo, en este mismo estudio, el tiempo medio generacional fue mayor (43,88 días) que el obtenido para N. tenuis (28,7 días).
El tiempo medio generacional, que se plantea corto para este tipo de especie, al ser menor de un mes se considera una ventaja para el incremento poblacional del depredador; por lo que los efectos a largo plazo para las dos especies podrían ser similares.
En el caso del crecimiento (rm=0,111), capacidad de multiplicación de una población, la tasa finita de crecimiento (λ=1,12), así como el doble tiempo generacional (DT=6,3), fueron ligeramente superiores a los informados por Perdikis y Lykouressis (15) para M. pygmaeus (rm=0,104; λ=1,11; DT=6,67).
Estos resultados aseguran que las poblaciones de N. tenuis son capaces de multiplicarse y reproducirse cuando se alimentan de B. tabaci sobre plantas de tomate. Lo que, unido a otros estudios, pudieran avalar su potencialidad como un posible agente de control biológico para B. tabaci en las condiciones de Cuba.
Tomando en cuenta que la temperatura para M. pygmaeus es la más favorable según estos mismos autores y que los depredadores pertenecen a la misma familia al igual que las especies de presas utilizadas en ambos estudios, los resultados para N. tenuis pueden ser considerados como indicadores de un buen candidato para el control biológico de especies fitófagas (24).
En el estudio realizado por Kakimoto et al. (25) de la especie B. tabaci biotipo B sobre tomate (25ºC), se evidenció una tasa intrínseca de crecimiento (rm) de 0,11 en un tiempo de desarrollo de 25,6 días.
Aunque se conoce que la variedad de las plantas pudiera influir en el comportamiento de los parámetros biológicos de una especie de plaga (26), podemos inferir que el depredador posee una capacidad de multiplicación y un tiempo de desarrollo muy similar al fitófago, lo que confirma la potencialidad del depredador como agente de control biológico de esta presa.
Es conveniente recordar que el valor de rm, en condiciones controladas, está regido por la natalidad y la mortalidad de los individuos de la población, mientras que en condiciones naturales existen otros factores que afectan el equilibrio, como la inmigración y emigración de los individuos, además de los cambios ambientales y la presencia de enemigos naturales, lo cual es un aspecto que hay que considerar en la aplicación práctica del depredador (23).
Se pudo observar una correspondencia entre el periodo de mayor fecundidad con el de mayor sobrevivencia (Fig.1 y 2), lo que puede traducirse como el periodo de mayor número de hembras vivas y aptas para ovopositar.
En los estudios reproductivos realizados para M. pygmaeus y N. tenuis (27) se encontraron diferencias significativas en el comportamiento sexual de las hembras de estas dos especies. Las hembras de M. pygmaeus fueron monándricas, con un periodo de cortejo obligatorio, mientras que las de N. tenuis se comportaron como poliándricas, con un alto porcentaje de cópulas repetidas, luego de 24 horas de un llamado periodo refractario (tiempo entre cópulas) para ambos sexos. En ambos casos los machos fueron poligínicos.
Durante esta investigación se observa que parece suficiente una sola cópula para abastecer de esperma a las hembras de M. pygmaeus durante al menos 21 días; mientras que en el caso de N. tenuis el 60 % de las hembras se quedaron sin espermatozoides solo una semana después del apareamiento (27).
Los resultados de Franco (27) pudieran explicar el periodo donde la sobrevivencia de las hembras desciende. Este comportamiento pudiera estar dado por el hecho de haber sometido la hembra a un macho durante el experimento, lo cual no le brindó la posibilidad de interactuar con otros machos. Si se toma en consideración lo anterior, se puede inferir que las hembras de N. tenuis necesitan más de un apareamiento para reponer el reservorio de esperma con el que fertilizan sus huevos.
Esta situación hace pensar en replantear una serie de experimentos y metodologías usadas de forma general para los depredadores sin tomar en cuenta particularidades tan esenciales para las especies, como es su vínculo sexual. Forzar a los machos polígamos a la monogamia durante varias generaciones causa una reducción en el tamaño de los testículos con relación a los machos control en varias especies de insectos (i.e, moscas amarillas (28), escarabajos del estiércol (29), además de que resulta una desventaja para la fertilización.
Según estos resultados y los obtenidos por Franco (27), es importante considerar y prestar especial cuidado en la relación entre sexos del depredador N. tenuis para el desarrollo de crías masivas y durante su empleo como agente de control biológico. En este sentido será necesario mantener en las colonias un número superior de machos que de hembras, ya que estas últimas necesitan acoplarse de forma periódica para reabastecerse de esperma.
-
Miller GT. Sustaining the Earth, 6th edition. Thompson Learning, Inc. Pacific Grove, California. Chapter 9, 2004: 211-216.
-
Gilden RC, Huffling K, Sattler BB. Pesticides and health risks. J. Obstet. Gynecol. Neonatal. Nurs. 2010, 39(1): 103–109.
-
Andorno, A.V. Evaluación del sistema planta hospedera-huésped alternativo como estrategia para el control biológico de pulgones (Hemíptera: Aphididae) en sistemas de producción hortícola en cultivos protegidos. Tesis de Doctorado. Universidad de Buenos Aires, Argentina. 2012. 171 pp
-
Gurr, GM, Wratten SD, Barbosa P. Success in Conservation Biological Control of Arthropods Chapter 4. En: G. Gurr y S. Wratten (Eds.) Biological Control: Measures of success. Kluwer Academic Publishers. 2000: 105- 132
-
Bueno VHP, van Lenteren JC. Predatory bugs (Heteroptera). En: A.R. Panizzi, J.R.P. Parra (Eds.). Insect Bioecology and Nutrition for Integrated Pest Management. CRC Press, Boca Raton, FL. 2012: 539-569.
-
Urbaneja PB , Alonso M, Tena A, Bolckmans K, Urbaneja A. Sugar as nutritional supplement for the zoophytophagous predator Nesidiocoris tenuis. BioControl. 2013; 58: 57–64.
-
Martínez MA, Duarte L, Baños HL, Rivas A, Sánchez A. Predatory mirids (Hemiptera : Heteroptera : Miridae) in tomato and tobacco in Cuba. Rev. Protección Veg. 2014; 29:204-207.
-
Baños HL, Louzada E, Moura N, Martínez M, Bueno VE. Life table of Macrolophus basicornis (Hemiptera: Miridae) preying on Myzus persicae (Sulzer) and Macrosiphum euphorbiae (Thomas) (Hemiptera: Aphididae). Rev. Protección Veg. 2014; 29(2): 94-98.
-
Pérez M, Urbaneja A. Prospects for predatory mirid bugs as biocontrol agents of aphids in sweet peppers. J Pest Sci. 2015; 88:65–73.
-
Itou M, Watanabe M, Watanabe E, Miura K. Gut content analysis to study predatory efficacy of Nesidiocoris tenuis (Reuter) (Hemiptera: Miridae) by molecular methods. Entomological Science. 2013; 16: 145–150.
-
Bueno VHP, van Lenteren JC, Lins JCJr, Calixto AM, Montes FC, Silva DB, Santiago LD, Pérez LM. New records of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) predation by Brazilian Hemipteran predatory bugs. J. Appl. Entomol. 2013; 137(1-2):29-34
-
Baños HL., Ruiz TG, del Toro MB, Miranda I. Consumo y respuesta funcional de Nesidiocoris tenuis Reuter (Hemiptera: Miridae) frente a estadios inmaduros de Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae). Rev. Protección Veg. 2016: 31 (3): 217-223
-
Lins JC. Search capacity, prey preference, predation rates and reaction to prey and predator induced volatiles of predatory mirids of two tomato pests, Tuta absoluta (Lep.: Gelechiidae) and Bemisia tabaci (Hem.: Aleyrodidae). Tese (doutorado), Universidade Federal de Lavras, Lavras. 2014; 116pp.
-
Sánchez JA., A. Lacasa, J. Arnó, C. Castane y O. Alomar. Life history parameters for Nesidiocoris tenuis (Reuter) (Het., Miridae) under different temperature regimes. J. Appl. Entomol. 2009; 133: 125–132.
-
Perdikis DC, Lykouressis DP. Life table and biologi- cal characteristics of Macrolophus pygmaeus when feeding on Myzus persicae and Trialeurodes vaporariorum. Entomol. Exp. Appl. 2002; 102: 261–272.
-
Martínez GH, Román FR, Sáenz MG, Pérez IM, Marco VS. Optimizing Nesidiocoris tenuis (Hemiptera: Miridae) as a biological control agent: mathematical models for predicting its development as a function of temperature. Bulletin of Entomological Research. 2016; 106: 215–224.
-
Fauvel, G, Malausa J., Kaspar B. Etude en laboratoire des principales characteristiques biologiques de Macrolophus caliginosus (Heteroptera: Miridae). Entomophaga. 1987; 32: 529–543.
-
Brakefield PM, Kesbeke F. Genotype−environment interactions for insect growth in constant and fluctuating temperature regimes. Proc. R. Soc. Lond. B. 1997; 264:717-723.
-
Hamdi F, Chadoeuf J, Bonato O. Functional relationships between plant feeding and prey feeding for a zoophytophagous bug. Physiol Entomol 2013; 38(3): 241-245.
-
Ferkovich SM, Venkatesan T, Shapiro JP, Carpenter JE. Presentation of artificial diet: effects of composition and size of prey and diet domes on egg production by Orius insidiosus (Heteroptera: Anthocoridae). Flor. Entomol. 2007; 90(3):502-508.
-
Cohen AC, Byrne DN. Geocoris punctipes as a predator of Bemisia tabaci: a laboratory evaluation. Entomol. Exp. Appl. 1992; 64 (2):195-202.
-
Urbaneja A, Tapia G, Stansly P. Influence of host plant and prey availability on developmental time and surviorship of Nesidiocoris tenuis (Het. Miridae). Biocon- trol Sci. Technol. 2005; 15: 513–518.
-
Maselou DA, Perdikis DCh, Sabelis MW, Fantinou AA. Use of plant resources by an omnivorous predator and the consequences for effective predation. Biological Control 2014; 79: 92–100.
-
Baños HL. Tamarixia radiata Waterson (Hymenoptera: Eulophidae ): agente de control biológico para la regulación de Diaphorina citri Kuwayama (Hemiptera: Liviidae). Tesis de Doctorado, Universidad Agraria de la Habana, Mayabeque. 2013: 89pp
-
Kakimoto K, Inoue H, Yamaguchi T, Ueda S, Honda K, Yano E. Host plant effect on development and reproduction of Bemisia argentifolii Bellows and Perring (B. tabaci [Gennadius] B-biotype) (Homoptera: Aleyrodidae). Appl. Entomol. Zool. 2007; 42: 63-70.
-
Calvo J, Bolckmans K, Stansly PA, Urbaneja A. Predation by Nesidiocoris tenuis on Bemisia tabaci and injury to tomato. Bio Control. 2009; 54:237–246.
-
Franco KV. Biología de la reproducción de los míridos depredadores Macrolophus pygmaeus y Nesidiocoris tenuis. 2010; 132pp
-
Hosken DJ, Ward PI. Experimental evidence for testis size evolution via sperm competition. Ecology Letters. 2001; 4:10-13.
-
Simmons LW, García FG. Evolutionary reduction in testes size and competitive fertilization success in response to the experimental removal of sexual selection in dung beetles. Evolution. 2008; 62: 2580-2591.
Recibido: 31/3/2017
Aceptado: 6/7/2017
* Autor para correspondencia: Heyker L. Baños-Díaz. E-mail: hlellani@censa.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}