










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
En Cuba, Chloridea (= Heliothis) virescens (Fabricius) (Lepidoptera: Noctuidae), conocida como el cogollero del tabaco, se reconoce como la plaga clave de este cultivo (1,2,3,4). Sin embargo, recientemente se notificó la presencia de Chloridea tergemina (Felder y Rogenhofer) en la provincia Las Tunas (5,6), la que también se informó en Colombia (7) y Venezuela (8), donde se especificó que esta especie siempre estuvo asociada al cogollo y C. virescens a las partes reproductivas de las plantas de tabaco (8).
En el cultivo del tabaco negro está establecida la realización del desbotonado (eliminación de la yema terminal del tallo, cogollo) como una labor fitotécnica importante en la obtención de una hoja de elevada calidad para la industria. Con esta labor y los deshijes (eliminación de las yemas axilares) sucesivos, se deberá impedir que el cultivo florezca (4). El incumplimiento de estas dos actividades incurre en una violación tecnológica del proceso productivo, pues la floración trae consigo cambios en la composición química de las hojas que influyen en los parámetros de calidad.
Sin embargo, en áreas dedicadas al cultivo del tabaco en la provincia Pinar del Río, se observaron larvas de C. virescens asociadas al cogollo con características morfológicas diferentes a otras que se localizaban en las estructuras reproductivas (flores y cápsulas) de plantas donde no se realizó el desbotonado.
Las larvas del cogollo, en contraste con las ubicadas en las flores y cápsulas, presentaban una coloración verde-amarillenta, sin la presencia de manchas y franjas longitudinales oscuras a lo largo del cuerpo. Por otra parte, no se encontraron diferencias marcadas en el estado adulto, lo que condujo a la búsqueda de variabilidad biológica, partiendo de la variabilidad fenotípica encontrada en las larvas.
En Cuba, se realizaron algunos estudios sobre la biología de C. virescens en tabaco (3,9,10,11); sin embargo, no se tuvo en cuenta la variabilidad morfológica y la ubicación de las larvas en la planta, elementos importantes en el establecimiento de estrategias de manejo de plagas.
El estudio tuvo como objetivo evaluar el comportamiento de los parámetros biológicos y la tabla de vida en dos poblaciones de C. virescens asociadas al cogollo y a las cápsulas de plantas de tabaco negro.
MATERIALES Y MÉTODOS
El estudio biológico se desarrolló a partir de larvas recolectadas en el cogollo (designado como Grupo A) y en los frutos (cápsulas, designado como Grupo B) de plantas de tabaco negro del cultivar de (Habana-92( (12), que se cultivaron en áreas de la Estación Experimental del Tabaco de San Juan y Martínez (N22o16´46,488¨, W83o49´35,04¨), provincia Pinar del Río.
Las larvas se individualizaron en placas Petri y se les dio seguimiento hasta la formación de las pupas, las que se sexaron e individualizaron en frascos de cristal; se sellaron con malla antiáfidos y banda elástica, hasta la emergencia de los adultos. Cuando emergieron, se formaron parejas de cada grupo en frascos con capacidad de 500 ml. Para la alimentación, se colocó algodón embebido en solución de agua destilada con miel de abeja (3:1), el cual se renovó cada 24 horas; también se colocaron dos tiras de papel de 25 x 2 cm, en forma de acordeón, para facilitar la puesta de huevos.
Iniciada la oviposición, se seleccionó una cohorte de 50 larvas por grupo, emergidas el mismo día. Se individualizaron en placas Petri (150 mm X 30 mm) y se les siguió el desarrollo hasta la muerte de los adultos, en condiciones de temperatura media de 24,02 ± 0,24°C y 82,52 ± 0,72 % de humedad relativa. Los datos de temperatura y humedad fueron registrados en un higrotermómetro digital.
Teniendo en cuenta que la hoja es el órgano más importante en este tipo de tabaco, las larvas se dispusieron sobre hojas del cogollo de plantas de tabaco negro del cultivar ‛Habana-92’, sanas y libres de aplicaciones de plaguicidas, cultivadas en la Estación Experimental. Las placas Petri se revisaron cada 24 horas para la limpieza, cambio de alimentación y determinar el momento de muda de la cápsula cefálica.
Se evaluaron la duración del periodo de incubación, la duración de la fase larval y los estadios larvales, la prepupa y pupa, así como el número de huevos.hembra-1.día-1, la fecundidad media/♀, la duración de los periodos prerreproductivo, reproductivo y posreproductivo, la longevidad de los adultos y la duración del ciclo biológico.
A partir de los datos de sobrevivencia de la hembra y la ovoposición diaria, se calculó la tasa neta de reproducción (R0), tasa intrínseca de incremento natural (rm), tiempo medio generacional (T), doble tiempo generacional (TD) y tasa finita de incremento (λ) a través de las fórmulas siguientes (13):
 (1)
(1)
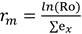 (2)
(2)
 (3)
(3)
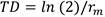 (4)
(4)
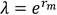 (5)
(5)
Los datos de la biología en ambos grupos se procesaron a través de un análisis de varianza (ANOVA) y las medias se compararon según rangos múltiples de Duncan para p≤0,05, utilizando el Paquete estadístico InfoStat-2016 (14).
RESULTADOS Y DISCUSIÓN
Biología
La duración del periodo de incubación en los Grupos A y B fue de tres días, que se encuentra dentro del intervalo de dos a cinco días informado, por diversos autores, para C. virescens en tabaco, algodón y garbanzo (2,3,11,15,16,17).
Las larvas de cada grupo completaron el estado larval en seis instares, lo que se confirmó a través de las exuvias y cápsulas cefálicas encontradas, resultados que concuerdan con los informados para la especie C. virescens en el cultivo del tabaco (3,17) y garbanzo (16,18).
Asimismo, se encontraron diferencias significativas entre los grupos con relación a la duración del estado larval (Tabla 1). Las larvas del Grupo B culminaron la fase larval en menor tiempo, aun cuando estuvieron bajo las mismas condiciones de temperatura y humedad relativa que las del Grupo A; lo cual pudo estar influenciado por el cambio de alimento y de cápsulas (progenitores) a hojas del cogollo (descendencia).
TABLA 1 Parámetros biológicos de los Grupos A y B de Chloridea virescens F. sobre el cultivar (Habana-92(. / Biological parameters of the Groups A and B of Chloridea virescens F., estimated on the black tobacco, cultivar (Habana-92(.
| Parámetros Biológicos | Grupo A | Grupo B |
|---|---|---|
| X± DS | X± DS | |
| Duración de la fase larval (días) | 21,27± 0,35 a | 18,36± 0,36 b |
| L1 | 2,34± 0,05 b | 3,00± 0,05 a |
| L2 | 3,24± 0,08 a | 2,24± 0,08 b |
| L3 | 2,68± 0,10 b | 3,38± 0,10 a |
| L4 | 3,04± 0,07 a | 2,22± 0,08 b |
| L5 | 2,30± 0,06 a | 2,05± 0,07 b |
| L6 | 2,37± 0,09 b | 2,31± 0,10 b |
| Duración de la pre-pupa (días) | 2,02 ± 0,09 a | 1,64 ± 2,02 b |
| Duración de la pupa (días) | 11,51± 0,17 b | 12,11± 0,18 a |
Medias con letras diferentes, en una misma fila, difieren significativamente (p≤0,05)
Sin embargo, la duración del estado larval en ambos Grupos se encuentra dentro de los valores informados en Cuba para C. virescens sobre tabaco, con una duración de 17 a 22 días con temperaturas de 27,3 °C a 29,5 °C (3,11).
Del mismo modo, los grupos A y B se diferenciaron, significativamente, en cuanto a la duración de las fases de prepupa y pupa (Tabla 1). Al respecto solo se cuenta con la información de Méndez (3), quien señaló que el estado de pupa transcurrió en 10 y 11 días a temperaturas de 27,3°C a 29°C, cuando las larvas se alimentaron de tabaco; por otra parte, Capinera (17) informó que la duración en esta fase fue de 11 días a 30°C, 13 días a 25°C y 22 días a 20 °C.
Los dos grupos fueron semejantes en cuanto a la duración de los periodos reproductivos y la longevidad de los adultos. Sin embargo, la duración del ciclo de vida tuvo diferencias significativas entre las hembras y machos de ambos grupos, con una mayor duración en los del Grupo A. Estos resultados fueron superiores a los notificados por diferentes autores para C. virescens (1,3,9,19,20). (Tabla 2)
TABLA 2 Duración del periodo reproductivo, ciclo de vida y longevidad de los Grupos A y B de Chloridea virescens F. sobre el cultivar de tabaco negro (Habana-92(. / Duration of the reproductive period, life cycle, and longevity in the Groups A and B of Chloridea virescens F. estimated on the black tobacco, cultivar (Habana-92(.
| Parámetros Biológicos (Días) | Grupo A | Grupo B | |
|---|---|---|---|
| X±DS | X±DS | ||
| Prerreproductivo | 4,25± 0,67 b | 4,67± 0,70 b | |
| Reproductivo | 5,00±0,66 b | 4,67± 0,76 b | |
| Posreproductivo | 3,75± 1,17 b | 3,33± 1,35 b | |
| Duración ciclo vida | ♀ | 50± 0,6 a | 44± 0,2 b |
| ♂ | 42,5± 0,2 a | 40,0± 0,1 b | |
| Longevidad | ♀ | 13,00± 1,65 b | 12,67± 1,91 b |
| ♂ | 10,25± 1,37 b | 9,67± 1,58 b | |
| Relación Sexual | 0,5 | 0,6 | |
Medias con letras diferentes, en una misma fila, difieren significativamente (p≤0,05)
El coeficiente sexual mostró un equilibrio entre la producción de hembras y machos en el Grupo A; mientras que en el Grupo B este coeficiente fue de 60 %, lo que evidencia una mayor producción de hembras en este Grupo.
Es importante señalar que, en el presente estudio, solo se discuten los resultados con C. virescens por no disponer de informaciones sobre este tema de otras especies del género con relación al cultivo del tabaco, tanto en Cuba como en otros países.
Tabla de vida
La fecundidad total de las hembras del Grupo A fue de 833 huevos y dos veces superior a las del Grupo B con 384 huevos; mientras que la viabilidad fue similar en ambos grupos con un 90,3± 1,48 % y 90,5± 2,10 %, respectivamente.
El máximo número de huevos en el Grupo A se alcanzó en la segunda puesta con 241 huevos; en tanto, en el Grupo B se alcanzó en la cuarta puesta con un máximo de 128 huevos. A partir de la máxima fecundidad y hasta el término de las puestas, el número de huevos descendió en ambos grupos. (Figura 2)

FIGURA 1 Fecundidad media (mx) de los Grupos A y B de Chloridea virescens F. sobre el cultivar de tabaco negro (Habana-92(. / Average fertility (mx) of the Groups A and B of Chloridea virescens F., estimated on the black tobacco cultivar (Habana-92(.
La curva de supervivencia de las poblaciones de los Grupos A y B, sobre hojas del cultivar de tabaco negro (Habana-92( (Figura 1), se asemeja a una curva de tipo I, desde el inicio de la cohorte hasta aproximadamente el día ocho en el Grupo A y el siete en el Grupo B, donde la probabilidad de sobrevivir a las edades tempranas es igual a uno; mientras que, a partir de ese mismo momento, se describe una curva de Tipo II, donde la tasa de mortalidad del Grupo B varía poco, ya que la curva presenta una forma diagonal descendente, mientras que en el Grupo A mostró una curva de forma sigmoidea, donde el número de individuos que muere en cada tramo se considera más o menos constante.
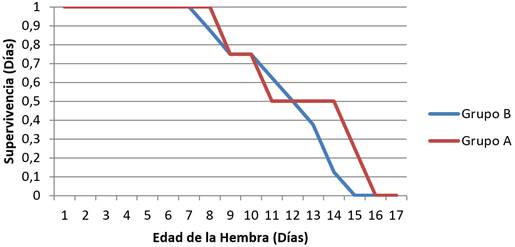
FIGURA 2 Sobrevivencia (lx) de los Grupos A y B de Chloridea virescens F. sobre el cultivar de tabaco negro (Habana-92(. / Survival (lx) of the Groups A and B of Chloridea virescens F., estimated on the black tobacco cultivar (Habana-92(
La tasa neta de reproducción (R0) fue dos veces superior en el Grupo A con respecto al Grupo B. Este resultado indicó la alta capacidad reproductiva que tienen las poblaciones de los dos grupos; o sea, que por cada hembra de la generación del Grupo A, habrá dos hembras más en la generación siguiente, que las hembras que aportaría cada hembra del Grupo B (Tabla 3).
No obstante, este resultado se debe tener en cuenta en el manejo de esta plaga, pues se evidenció que, sobre las hojas del cogollo, la población del Grupo B es capaz de lograr poblaciones importantes y provocar daños al cultivo.
TABLA 3 Parámetros poblacionales de los Grupos A y B de Chloridea virescens F. sobre el cultivar de tabaco negro (Habana-92(/ Population parameters of the Groups A and B of Chloridea virescens F., estimated on the black tobacco cultivar (Habana-92(.
| Grupo | R0 | rm | T (días) | TD (días) | λ |
|---|---|---|---|---|---|
| A | 826,50 | 0,17 | 39,14 | 4,30 | 1,19 |
| B | 378,00 | 0,16 | 36,84 | 4,04 | 1,17 |
Tasa neta de reproducción: (Ro), Tasa intrínseca de crecimiento: (rm), Intervalos medio entre generaciones: (T), Tiempo de duplicación: (TD) y Tasa finita de incremento (λ).
Por otro lado, la tasa intrínseca de crecimiento (rm) tuvo una diferencia del uno por ciento entre las poblaciones de los dos grupos y predice un crecimiento poblacional de 17 % en el Grupo A y de 16 % en el Grupo B de un día al siguiente, cuando las larvas se alimentaron con hojas del cogollo (Tabla 3). Este resultado confirmó la potencialidad de ambas poblaciones como plagas en este cultivo
El tiempo generacional fue menor en la población del Grupo B, lo que se puede explicar a través de los factores que afectaron el ciclo de vida, como el cambio de alimentación. Sin embargo, las dos poblaciones requieren aproximadamente el mismo tiempo para duplicar una generación. Elementos de gran importancia a tener en cuenta en el manejo de estas poblaciones. (Tabla 3)
La tasa finita de crecimiento (λ) fue similar para los dos grupos e indicó que, por cada hembra presente en un día, habrá aproximadamente 1,2 hembras al día siguiente, lo que predice un incremento sostenido de estas poblaciones, al ser λ>1 y refuerza la idea de que ambos grupos pueden mantener un alto nivel de sus poblaciones sobre hojas de tabaco.
Los parámetros biológicos y poblacionales caracterizan la dinámica potencial de una población en condiciones de laboratorio, donde el recurso alimenticio se encuentra en exceso y no hay hacinamiento ni causas externas de mortalidad (parasitoidismo y depredación), es de esperar que los organismos expresen su mayor potencial de supervivencia que sirve para discutir sus efectos en escenarios más complejos (21).
Por tanto, los resultados alcanzados sugieren que las poblaciones, tanto del cogollo como de las cápsulas, están relacionados con C. virescens. Sin embargo, el cambio de alimentación influyó en la duración del ciclo de vida y en los parámetros poblacionales del Grupo B; lo que sugiere que esta población está más adaptada a las estructuras reproductivas de la planta, lo cual es posible que esté condicionado por un compartimento de ambas poblaciones por el nicho trófico y relacionado con las diferencias de la composición química entre cápsulas y hojas de tabaco.

