










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Orthezia praelonga Douglas, 1891 (Hemiptera: Sternorrhyncha: Ortheziidae), conocida como piojo blanco, es una cochinilla que causa pérdidas económicas importantes en cultivos citrícolas (1,2) y plantas ornamentales (3). Los perjuicios pueden ser directos cuando se alimenta de savia de las hojas, o indirectos por la excreción de una sustancia azucarada “honeydew”, que cuando está acumulada promueve el desarrollo de hongos; tal es el caso de Capnodium sp., que cuando cubre gran cantidad de hojas reduce el área fotosintética de la planta (4) y cuando alcanza los frutos afecta su valor comercial (5).
Algunas características, como longevidad, fecundidad, prolificidad, diversidad de plantas hospederas (6,7,8) y gran capacidad de dispersión (9), favorecen la presencia de esta plaga durante todo el año (10). En condiciones adecuadas, como baja temperatura, precipitación y humedad relativa, surgen picos poblacionales de O. praleonga (11), lo que la convierte en una plaga de importancia económica, principalmente en estas temporadas del año.
El control de O. praleonga en huertos citrícolas se realiza con el uso de insecticidas organofosforados, piretroides y neonicotinoides (6,12); sin embargo, la cochinilla adulta posee una estructura abdominal (ovisaco), donde almacena sus huevos y los protege de agentes externos (13). En campo, las estrategias para superar esta barrera pueden conllevar a incrementar la dosis recomendada o usar productos altamente tóxicos, lo que contribuye a la contaminación ambiental, surgimiento de organismos resistentes y desequilibrio biológico en el agroecosistema (14,15,16). Algunos organismos reguladores de O. praleonga, como Gitona brasiliensis Lima, 1950 (Diptera: Drosophilidae), Scymnus sp., Azya luteipes Mulsant, 1850 (Coleoptera: Coccinellidae), Heza insignis Stal, 1859 (Hemiptera: Reduviidae) y varias especies dos géneros Chrysopodes spp. y Ceraeochrysa spp (Neuroptera: Chrysopidae), pueden ser reducidos drásticamente por insecticidas de uso común (2). Entre estos predadores, crisopas es un grupo considerado importante, debido a su alto potencial biótico, voracidad y plasticidad ecológica (17,18). Se deben priorizar, en el manejo integrado de esta plaga, los productos alternativos con efecto insecticida y que presenten selectividad a enemigos naturales.
Los aceites vegetales son derivados de plantas oleaginosas y se caracterizan por contener en su composición abundantes ácidos grasos; se destacan los ácidos esteárico, palmítico, oleico, linoleico y linolénico (19). Los estudios demostraron bioactividad de estos compuestos a insectos (20,21,22), ácaros (23,24) y bacterias (25,26); además, su alto rendimiento en la extracción, el bajo precio y su disponibilidad en el mercado los convierte en una potencial alternativa para el manejo de O. praelonga. El presente estudio tuvo como objetivo evaluar la toxicidad y la repelencia de cuatro aceites vegetales a O. praelonga, así como su selectividad a su predador Ceraeochrysa caligata Banks, 1945 (Neuroptera: Chrysopidae).
MATERIALES Y MÉTODOS
Insectos
Las hembras de O. praelonga procedieron de plantaciones citrícolas exentas de la aplicación de agroquímicos, al menos por un año, ubicadas en el municipio Boquim (11°08’35”S; 37°38’35”O, 165 msnm) Sergipe - Brasil. Las cochinillas adultas recién emergidas se recolectaron en plantaciones de naranja (var. Pera), máximo 48 horas previo a cada bioensayo. El predador C. caligata proviene de colonias de laboratorio en la Empresa Brasileira de Pesquisa Agropecuaria (EMBRAPA, Tabuleiros Costeiros) en Aracaju, Sergipe - Brasil.
Aceites vegetales
Se usaron cuatro aceites vegetales presentes en el mercado nacional (Brasil): aceite de algodón (HG Comercial LTDA), soja desgomado (ABOISSA), coco y dendé (Aracaju-Sergipe-Brasil). Los procesos de extracción y análisis cromatográfico se describen para cada aceite vegetal: Oliveira et al. (23) para soja desgomado y coco, Freitas (27) para dendé y Teodoro et al. (24) para algodón. Estos aceites contienen ácido linoleico (C18:2) en 58,65 %, 19,77 %, 49,61 % y 47,97 % para el aceite de soja desgomado, coco, algodón y dendé, respectivamente; en ese mismo orden, el ácido palmítico (C16:0) aportó con 16,15 %, 15,21 %, 10,04 % y 34,79 % para cada aceite. El aceite de coco posee, además, los ácidos láurico (C12:2) y mirístico (C14:0) en 27,23 y 19,90 %; en tanto, los ácidos oleico (C18:1) y linolénico (C18:3) representan el 23,50 y 6,64 % en el aceite de algodón.
Bioensayo de toxicidad
Se realizaron los bioensayos de concentración de mortalidad para determinar concentraciones letales (CL) de los cuatro aceites vegetales a O. praelonga. Las arenas consistieron en hojas frescas de naranja ubicadas perfectamente en una placa Petri (78,5 cm2) con el envés expuesto hacia arriba. Se colocaron cinco cochinillas adultas recién emergidas en cada arena. Las concentraciones de cada aceite vegetal, previamente diluidos en agua destilada y detergente neutro (1 %), se aplicaron a través de una torre Potter (Burkard, Rickmansworth, UK) con presión de 0,34 bar (34 KPa), alícuota de 1,7 ml y deposición de 1,8 ± 0,1 mg/cm². Cada concentración y el control (agua más detergente neutro) tuvieron seis repeticiones (n = 30). Las arenas pulverizadas permanecieron a temperatura ambiente por 30 min. Enseguida se cubrieron con film plástico perforado y se mantuvieron en ambiente controlado (26 ± 2 °C; 70 ± 2 % de HR y 12 h de fotofase) por 24 h. Posteriormente, se evaluó la mortalidad; las hembras de O. praelonga se consideraron muertas cuando no respondieron al toque con un pincel.
Repelencia
El efecto repelente de los aceites vegetales se evaluó adaptando la metodología de área preferida (28). Se usaron las CL50 y CL80 de los aceites de soja desgomado, coco, algodón y dendé, respectivamente. Las hojas frescas de naranja, con el envés expuesto hacia arriba, se ubicaron en placas Petri (área libre 25 cm2) y se delimitaron con agar (3 %). Las arenas se dividieron en dos partes iguales y una mitad (control) se cubrió con doble camada de papel de alta densidad (Kraft ©). Posterior a la aplicación de cada concentración (conforme descrito en la toxicidad), se retiró el papel que cubría la mitad de la arena. Transcurridos 30 min de la aplicación, se transfirió una cochinilla adulta recién emergida al centro de cada arena, sobre una cinta adhesiva transparente (7 mm2). Posteriormente, las arenas se cubrieron con film plástico perforado y se mantuvieron a 26 ± 2 °C, 70 ± 2 % de HR, 12 h de fotofase. Las evaluaciones se realizaron transcurridas 1, 24 y 48 h, donde el número de cochinillas presente en cada área fue registrado. Cada concentración contó con 30 repeticiones.
Selectividad al predador Ceraeochrysa caligata
Las larvas del primer estadio de C. caligata (<12 horas de vida) se trataron con cada aceite vegetal a las CL50 y CL80, previamente estimadas para O. praelonga. En una placa Petri (15 cm de diámetro x 0,7 cm de altura), que contenía agua en su interior, se introdujeron diez tapas plásticas (arenas) de 2 cm de diámetro x 1 cm de altura. El interior de cada arena se cubrió con papel filtro (3,14 cm2) y luego se introdujo una larva; cada concentración tuvo tres repeticiones (n = 30). La aplicación de cada aceite vegetal, así como las condiciones ambientales, siguieron los procedimientos citados anteriormente. Posterior a la aplicación de los tratamientos, en cada arena se adicionaron huevos de Anagasta kuehniella Zeller, 1879 (Lepidoptera: Pyralidae) como alimento al predador. El número de insectos vivos se registró 24 h después de la de exposición; se consideraron muertos los insectos que no responden al toque con pincel.
Análisis estadístico
Las concentraciones letales CL50 y CL80 de cada aceite vegetal se obtuvieron por análisis Probit. La razón de toxicidad (RT) y sus intervalos de confianza (95 %) se obtuvieron y se consideraron diferentes cuando estas estimativas no incluyen el valor de 1 (29). Los datos de repelencia se sometieron a análisis de frecuencia mediante la prueba chi-quadrado; mientras que, el porcentaje de predadores vivos se comparó mediante análisis de varianza Kruskal-Wallis One Way ANOVA (p<0,05). La prueba T se usó para estimar la diferencia significativa entre las dos concentraciones letales dentro de un mismo aceite vegetal. Todos los análisis se realizaron con el paquete estadístico SAS versión 9.4 (30).
RESULTADOS Y DISCUSIÓN
Toxicidad
Todos los aceites vegetales resultaron tóxicos a O. praelonga cuando se expuso al contacto (Tabla 1). La razón de toxicidad para la CL50 varió entre 1,32-2,61 y mostró que en esta concentración no existe diferencia al existir sobreposición de intervalos (RT=1). Para la CL80, la razón de toxicidad varía de 1,71-3,75; no existió sobreposición para el aceite de coco (RT≥2), lo que revela que con la CL80 este aceite es el menos tóxico a O. praelonga (Tabla 1).
TABLA 1 Concentraciones letales estimadas (µl/ml) y sus intervalos de confianza (95 % IC) de cuatro aceites vegetales a hembras adultas de O. praelonga. / Estimated lethal concentrations (μl/ml) and their confidence intervals (95 % CI) of four plant oils to adults females of O. praelonga.
| Aceites vegetales | Número de insectos | Inclinación ± EE | CL50 (95% IC) | RT CL50 (95% IC) | CL80 (95% IC) | RT CL80 (95% IC) | χ² | |
|---|---|---|---|---|---|---|---|---|
| Algodón | 300 | 1,70 ± 0,17 | 1,92 (1,53 - 2,41) | - | 5,99 (4,54 - 8,64) | - | 1,83 | 0,99 |
| Dendé | 330 | 1,39 ± 0,13 | 2,54 (1,92 - 3,29) | 1,32 (0,98 - 1,78) | 10,22 (7,50 - 15,22) | 1,71 (1,35 - 2,15) | 6,94 | 0,64 |
| Soja desgomado | 360 | 1,42 ± 0,14 | 3,18 (2,46 - 4,02) | 1,65 (1,25 - 2,19) | 12,50 (9,37 - 18,28) | 2,09 (1,68 - 2,59) | 5,62 | 0,85 |
| Coco | 450 | 1,29 ± 0,12 | 5,02 (3,94 - 6,31) | 2,61 (1,99 - 3,42) | 22,45 (16,79 - 32,80) | 3,75 (2,98 - 4,72) | 12,51 | 0,49 |
CL: Concentración letal; χ²: chi-cuadrado; EE: Error estándar; P: probabilidad; RT: razón de toxicidad (CLs de cada aceite vegetal /CLs de aceite de algodón). Letras minúsculas identifican diferencias por los IC.
La toxicidad de los aceites vegetales se atribuye a la elevada cantidad de compuestos con cadenas de 18 carbonos (ácido linolénico, linoleico y oleico) en su composición (23,24). Aunque el análisis de la estructura y la actividad de los ácidos grasos no son bien comprendidos, existen estudios que demostraron que la toxicidad a artrópodos por estos compuestos depende de la longitud de la cadena. Las moléculas con cadenas de C10, C12 y C18 presentan mayor toxicidad que las moléculas con C14 y C16 indistintamente, sean saturadas o insaturadas (20,21).
Los resultados del presente estudio muestran al aceite de algodón y de dendé como los más tóxicos a O. praelonga, propiedad atribuida al ácido linoleico presente en alta cantidad en estos aceites. Este compuesto mostró actividad insecticida sobre Helicoverpa zea Boddie, 1850 y Spodoptera frugiperda Smith, 1797 (Lepidoptera: Noctuidae); Lymantria dispar Linnaeus, 1758 y Orgyia leucostigma Smith, 1797 (Lepidoptera: Lymantriidae) y Malacosoma disstria Hübner, 1820 (Lepidoptera: Lasiocampidae) (31,32). Los estudios realizados por Kannathasan et al. y Rahuman et al. (33,34) muestran al ácido oleico, también mayoritario en el aceite de algodón (23,5 %), como tóxico a Aedes aegyptii Linnaeus, 1762, Anopheles stephensi Liston, 1901 y Culex quinquefasciatus Say, 1823 (Diptera: Culicidae), lo que sugiere un efecto aditivo entre estos compuestos, que resulta en una elevada toxicidad a insectos cuando están expuestos al contacto. Contrariamente a lo ocurrido con el aceite de algodón y dendé, la toxicidad del aceite de soja desgomado y coco fue menor a estos; por lo que es posible los efectos antagónicos de compuestos característicos presentes en estos últimos.
Repelencia
El efecto repelente con las CL50 y CL80 de cada aceite vegetal mostró que el aceite de algodón fue repelente a O. praelonga en un corto periodo (1 h) de exposición CL50 (P = 0,0036) (Fig. 1A) y su efecto disminuyó después de 24 h (Figs. 1 B, C). Con una mayor concentración (CL80), los aceites de algodón y de dendé lograron repeler a O. praelonga por 1 h (P = 0,0377; P= 0,0391, respectivamente) (Fig. 2A). El aceite de algodón presenta mayoritariamente ácido linoleico y oleico en su composición; estos compuestos, combinados o aislados, son conocidos por sus propiedades repelentes a insectos (35,36); sin embargo, el efecto repelente del aceite de algodón disminuye con el tiempo. Bánsági y Taylor (37) sugieren que en algunos ácidos y compuestos glicolíticos es común que ocurra hidrólisis de enlaces éster, que resultan en monómeros solubles no tóxicos a plagas (38). Contrario a lo expuesto, el aceite vegetal de soja desgomado (CL80) se tornó repelente después de 24 horas (P = 0,0029) y mantuvo tal efecto durante todo el periodo de evaluación (P = 0,0047) (Figs. 2 B-C). Es conocido que la insaturación ocurre con frecuencia en ácidos grasos y que en compuestos menores juegan un rol importante en cada molécula. Esta reacción puede dar origen a metabolitos (aldeídos, ácidos carboxílicos) que son biológicamente activos a organismos (39), por lo que tal actividad es atribuida al aceite de soja. El aceite de coco con el ácido láurico (27,23 %) y mirístico (19,90 %), como compuestos mayoritarios, no presentó repelencia a O. praelonga. Resultados similares obtuvieron Dani et al. (35), pues estos compuestos no fueron repelentes a tres especies de hormigas.
Selectividad al predador Ceraeochrysa caligata
Los aceites vegetales tóxicos para O. praelonga presentaron selectividad para su enemigo natural C. caligata. Con cada CL50 estimada para la cochinilla, la sobrevivencia del predador fue mayor al 80 % (Tabla 2) y no se observó diferencia estadística entre todos los aceites (F = 5,15; gl= 4; P = 0,272). Al ser expuestos a la CL80, la sobrevivencia del predador fue reducida a un mínimo de 69 % (Tabla 2), sin existir diferencias entre todos los tratamientos (F = 7,48; gl= 4; P = 0,113). De igual forma, no se evidenció diferencia en sobrevivencia del predador al compararse los dos tratamientos dentro de un mismo aceite. (Tabla 2)
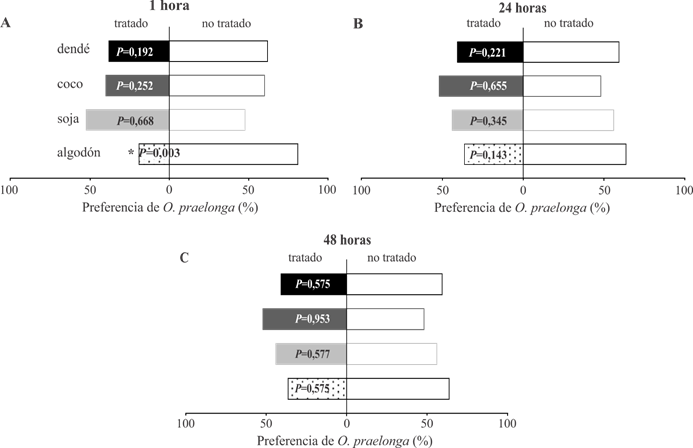
FIGURA 1 Porcentaje de O. praelonga en área tratada (CL50) y no tratada con aceites vegetales, después de 1, 24 y 48 h de exposición. Las barras corresponden a medias con nivel de significancia (p<0,05). / Percentage of O. praelonga in treated area (LC 50 ) and not treated with plant oils, after 1, 24 and 48 h of exposure. Bars correspond to means with significance level (p<0,05).

FIGURA 2 Porcentaje de O. praelonga en área tratada (CL80) y no tratada con aceites vegetales, después de 1, 24 y 48 h de exposición. Las barras corresponden a medias con nivel de significancia (p<0,05). / Percentage of O. praelonga in treated area (LC 80 ) and not treated with plant oils, after 1, 24 and 48 h of exposure. Bars correspond to means with significance level (p<0,05).
TABLA 2 Porcentaje media de C. caligata que sobrevivieron después de 24 h expuestos a concentraciones letales (CL50 y CL80) de cuatro aceites vegetales estimados para O. praelonga. / Average percentage of C. caligata that survived after 24 h exposed to lethal concentrations (LC 50 and LC 80 ) of four plant oils estimated for O. praelonga.
| Aceite vegetal | Número de vivos (%) | ||||
|---|---|---|---|---|---|
| CL50 | CL80 | ||||
| Algodón | 87,09 ± 13,80 | 74,16 ± 10,10 | 57 | 1,19 | 0,24 |
| Dendé | 85,24 ± 5,01 | 69,05 ± 11,45 | 52 | 1,31 | 0,19 |
| Soja desgomado | 86,67 ± 23,09 | 74,24 ± 5,17 | 56 | 1,02 | 0,31 |
| Coco | 80,00 ± 26,45 | 76,30 ± 10,91 | 57 | 0,38 | 0,70 |
Diferencias entre las CL50 y CL80 dentro de cada aceite vegetal fueron analizadas por el test t (p<0,05).
Varias especies de crisopas se conocen como predadores generalistas y contribuyen al control natural de varias plagas en cítricos; O. praelonga es una de estas. El elevado número de predadores vivos (C. caligata), cuando se expone a concentraciones letales de estos aceites vegetales, confirma su selectividad a este enemigo natural. Las larvas de crisopas se caracterizan por ser relativamente tolerantes a pesticidas de origen sintético (40,41,-42) y natural (43). Por otro lado, también se demostró la inocuidad de un aceite mineral y un compuesto derivado de aceite de neen Azadirachta indica (Meliaceae) a C. cubana (44). Así, los aceites vegetales se suman a otros productos naturales que presentan selectividad a enemigos naturales y son promisorios en programas de manejo integrado.
Los resultados de este estudio revelan que los aceites vegetales representan una alternativa para el control de la cochinilla O. praelonga. Se destacan los aceites de algodón, dendé y soja desgomado por su toxicidad, repelencia, selectividad a su enemigo natural y facilidad de acceso en el mercado.

