










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.32 no.3 La Habana jul.-set. 2015
TECHNIQUE
Development of sandwich ELISA-based assays for screening Bordetella pertussis antigens expressed in the bacterial cell culture
Desarrollo de un ensayo ELISA sandwich para el pesquisaje de antígenos de Bordetella pertussis expresados en cultivos de células bacterianas
Gilda Lemos-Pérez1, Maite Delgado-Espina1, Diógenes Quintana-Vazquez1, Edelgis Coizeau-Rodríguez1, Emilio Carpio-Muñoz2, Evelin Caballero-Menéndez1, Anabel Alvarez-Acosta1
1 Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba.
2 Centro de Ingeniería Genética y Biotecnología de Sancti Spíritus, CIGBSS, Cuba.
ABSTRACT
Pertussis is a highly contagious respiratory disease caused by Bordetella pertussis, a Gram negative bacterium. Concerns about whole cell vaccine safety led to the development of acellular pertussis vaccines with a lower frequency of adverse reactions. But they require a quick and reliable method for antigen detection, its quantification,or both. For this aim, a sensitive capture (sandwich) enzyme-linked immunosorbent assay (ELISA) using monoclonal antibodies was established, to identify and quantification the acellular pertussis vaccine components: detoxified Pertussis toxin (Ptg), Pertactin 2 (Prn2) and filamentous hemagglutinin (FHA). The Ptg-capture ELISA uses Fetuin in the solid phase, and Mabs 2G3 and 6D4 against Ptg were evaluated as detection reagents, the best results attained with Mab 2G3. The linearity range of the ELISA was 0.78-50 ng/mL. The Prn capture ELISA was established by evaluating Mabs 9C6 and 6B3 indistinctly as capture or detection reagents. The best combination was Mab 9C6 for capture and 6B3 for detection, with a linearity range 0.15-10 ng/mL. For the evaluation of FHA, monoclonal antibodies 11E8, 437 and 5E12 were evaluated indistinctly as capture or detection reagents, the optimal combination using Mab 11E8 as capture and Mab 437 for detection, with a linearity range 3.25-200 ng/mL. Under optimized ELISA tests conditions, the limit of quantification were 0.78, 0.07 ng/mL and 3.25 ng/mL for the Ptg, Prn2 and FHA proteins, respectively. Given its performance, the ELISA tests developed could be efficaciously implemented for process control during the production of these three B. pertussis vaccine antigens.
Keywords: ELISA, Bordetella pertussis, pertussis toxin, Pertactin 2, filamentous hemagglutinin, vaccine antigen.
RESUMEN
La pertusis o tosferina es una enfermedad respiratoria altamente contagiosa causada por la bacteria Gram negativa Bordetella pertussis. Cuestionamientos sobre la seguridad de las vacunas basadas en células completas han conducido al desarrollo de vacunas acelulares que generan reacciones adversas con menor frecuencia, pero que requieren de métodos rápidos y confiables para la detección de los antígenos vacunales, su cuantificación, o ambas. Con tal propósito, se establecieron ensayos ELISA de tipo sándwich con el uso de anticuerpos monoclonales (Mabs) para la detección de los componentes de una vacuna acelular: la toxina pertúsica detoxificada (Ptg), la pertactina 2 (Prn2) y la hemaglutinina filamentosa (FHA). El ELISA de captura contra Ptg emplea la fetuina en fase sólida, y se estableció el Mab 2G3 contra Ptg como reactivo de detección con mejores resultados. La linealidad del ensayo se mantuvo en el rango 0.78-50 ng/mL. Para el ELISA de captura de la Prn2 se obtuvo como mejor combinación la que empleó el Mab 9C6 para la captura y Mab 6B3 para la detección, con una linealidad en el rango 0.15-10 ng/mL. Para la FHA, la mejor combinación se obtuvo con el Mab 11E8 para la captura y el Mab 437 para la detección, con un rango de linealidad 3.25-200 ng/mL. Los límites de detección en las condiciones óptimas de los ensayos fueron: Ptg 0.78 ng/mL, Prn2 0.07 ng/mL y FHA 3.25 ng/mL FHA. Estos ensayos ELISA se pueden implementar eficazmente para el control de procesos durante la producción de los tres antígenos vacunales de B. pertussis.
Palabras clave: ELISA, Bordetella pertussis, toxina pertúsica, Pertactina 2, hemaglutinina filamentosa, antígeno vacunal.
INTRODUCTION
Pertussis or whooping cough is a highly contagious respiratory disease caused by Bordetella pertussis, a Gram negative bacillus first described by Bordet and Gengou in 1906 [1]. The disease still represents a major threat to public health. Estimates suggest that, in 2008, about 16 million cases of pertussis occurred worldwide and that about 195 000 children died from the disease, making pertussis the fourth largest cause of vaccine-preventable death in children under five years of age [2]. The bacterium is transmitted directly from human to human, probably via aerosolized respiratory droplets. Severe pertussis can lead to apnea, pneumonia, pulmonary hypertension, hypoxemic respiratory failure, and ultimately death [3].
Whole-cell pertussis vaccine (wP), which is composed of a suspension of formalin-inactivated B. pertussis cells, was used widely in clinical practice by the mid-1940s, resulting in a rapid reduction in both the incidence of pertussis and the number of deaths caused by the infection [4]. The vaccine is still in use in most developing countries, as in Cuba.
In the late 1990s, concerns about safety led to the development of more pure (i.e., acellular) pertussis vaccines (aP), with successful clinical trials showing lower frequencies of adverse reactions [5-8].
Among the antigens considered, pertussis toxin or Pt, filamentous hemagglutinin (FHA), pertactin (Prn or P69k) and fimbriae induce antibodies contributing to protection in an animal model, and the natural infection with B. pertussis in humans induces antibodies to each of these antigens [9]. New formulations of acellular B.pertusis vaccines containing protective antigens in different combinations have been proved to be safer, less reactogenic and more reliable to introduce in mass vaccination campaigns [10-13]. Also replacing the wP vaccine with aP vaccines has resulted in an increase in pertussis vaccination coverage in newborns and infants.
Nevertheless, the number of pertussis cases has increased and epidemics returned in the acellular vaccine era, especially in adolescents and adults, in spite of the effectiveness attained with pertussis vaccination [2, 14]. It is unknown whether the re-emergence of pertussis is due to the waning of vaccine-induced immunity and/or pathogen adaptation [15], or to fundamental differences in the nature of the immune responses induced by aP vaccines as compared with wP vaccines or with natural infection [16].
For these reasons, vaccination strategies have changed during the past years and booster doses have been implemented in national vaccination programs. The development of new vaccines is aimed to improve control by increasing population coverage, allowing more frequent boosters by the use of acellular pertussis-only vaccines, or making the boosters unnecessary by inducing a more protective or long-lasting immune response, for a better match to circulating strains, and/or allowing the immunization of neonates.
Considering the antigens used in acellular vaccines, Pt is the main toxin of B. pertussis and one of the most important virulence factors [17]. This ADP-ribosylating protein is a globular protein with a molecular weight of 117 kDa. Pt has several biological activities and it is composed of five subunits, S1 to S5, which assemble into two functionally distinct moieties [18]. The S1 subunit is responsible for the enzymatic activity, while subunits S2 to S5 are involved in binding to the target cell [19]. Previous experiments using monoclonal antibodies against Pt have demonstrated that the antibody specific for the S1 subunit neutralized the leukocyte-promoting and islet-activating activities of Pt and protected against B. pertussis infection [20- 22]. Pt induces high levels of antibody after infection or vaccination with classical whole cell vaccine, and the toxin provides full protection against challenge in mouse models. For this reason, inactivated Pt is considered the major protective antigen in all the new acellular vaccines available today [23, 24].
Regarding Prn, this is an auto-transporter protein that undergoes auto-proteolytic processing [25]. The protein is processed from a 93 kDa large precursor to a 69 kDa and 22 kDa proteins, located at the cell-surface and in the outer membrane, respectively [26]. The 69 kDa product is used in aP vaccines. Prn contains an Arg-Gly-Asp (RGD) motif involved in mediated attachment of B. pertussis to mammalian cells [27], is a polymorphic protein and 13 variants (P.69 Prn1 to P.69 Prn13) have been identified so far [28]. Variation is mainly limited to two regions, designated region 1 and two, which are comprised of Gly-Gly- X-X-Pro (r1 repeat) and Pro-Gln-Pro (r2 repeat) re-peats, respectively, more variation found in region 1 near to the N-terminus and flanking the RGD sequence. Region 2 is located towards the C-terminus. It is reported that the r1 repeat induces type-specific antibodies which show little cross-reactivity between P.69 Prn1and P.69 Prn2 [29].
In the case of FHA, this is a large and complex molecule [30, 31] that is synthesized as a 367-kDa precursor (FHAB), translocated to the periplasm and exported through the outer membrane [32-34]. N-terminal processing [33] and cleavage of the C-terminal third of FHAB yield the 220-kDa mature FHA molecule [32, 34]. FHA is a multifunctional adhesin that is both cell-associated and secreted into the external milieu. FHA improves vaccine efficacy when included in multicomponent aP vaccines and in animal models, FHA alone elicits protective immunity [35, 36].
The variety of aP vaccines formulations approved for human use, which is a distinctive feature with respect to all other bacterial vaccines, should not be expected to confer equal levels of efficacy and long-term protection. Formulations with two (Pt and FHA) to five (Pt, FHA, pertactin, and fimbriae 1 and 2) immunizing antigens have been licensed for human use [13, 37, 38].
Specifically in Cuba, efforts are been made to reduce the reactogenicity attained with the whole-cell pertussis vaccine available, for use in booster doses in children aged above five years and in pregnant women after the third month, included in the National Immunization Program [39]. A new vaccine under development is an acellular vaccine composed of antigens from a B. pertussis strain genetically modified to produce a detoxified pertussis toxin (Ptg) and to express the Prn2 protein as the most frequent variant in clinical isolates. Antigens Ptg and FHA are isolated from the supernatant of the bacterial culture medium and Prn2 is obtained from heat inactivated cells precipitate.
Hence, this work was aimed to establish sensitive and simple capture (sandwich) enzyme-linked immunosorbent assays (ELISA) for the screening of pertussis antigens and readily transferable to process development units during the bioproduction scaling process. The tests developed was effectively used to identify and quantify the Ptg, Prn2 and FHA antigens, expressed by B. pertussis bacterium as acellular pertussis vaccine components.
MATERIALS AND METHODS
B. pertussis culture
B. pertussis strain BpCNIC0311 is a derivative of Bp165 strain, genetically modified to produce the pertussis toxoid PTg (ptxA2, 9K/129G) under the regulation of the ptxP1 promoter and the pertactin 2 protein (prn2). The fhaB gene encoding filamentous hemagglutinin (FHA) remained as a wild type [40]. Bacteria was grown for 18 h in the defined THIJS medium as described by Thalen et al. [41] and modified by the addition of aminoacids and 2,6-di-o-dimethyl-beta-cyclodextrin (CDX). For monitoring the antigens expression by ELISA tests, three experimental batches were produced in an INFORS HT bioreactor. Cultures were maintained at 35 °C with agitation from 100 to 150 rpm, at pH 7.3 and 20 ± 5 % oxygen. Samples were taken at 0, 3, 6, 9, 12, 15 and 18 h of culture.
Proteins and antibodies
Antigens were purified as previously described by Quintana-Vazquez et al. [40]. Pt and FHA were isolated from NaCl-washed cells and clarified culture medium supernatant. Prn was obtained from heat-inactivated and precipitated bacterial cells by Tris/NaCl extraction. Pertussis antigens were purified by sequential filtration, salt-precipitation and/or extraction, ultrafiltration and ion exchange chromatography [40].
Mabs and peroxidase-conjugates against pertussis antigens were supplied by the Center for Genetic Engineering and Biotechnology of Sancti Spiritus (CIGBSS). The specificities of the Mabs are the following: anti-Ptg: Mabs 6D4 and 2G3; anti-Prn2: Mabs 9C6 and 6B3; anti-FHA: Mabs 11E8, 5E12 and 437.
Mabs were biotinylated using EZ-Link® Sulfo- NHS-LC-Biotin (Cat. # 21335, Pierce Biotechnology, USA), according to manufacturer’s specifications. Fetuin (Cat. # F3004) and streptavidin-peroxidase were purchased from Sigma-Aldrich, USA.
B. pertussis proteins reference standards
NIBSC reference proteins were used as standards for the development of the ELISA tests: Pt (JNIH 90/518), Prn (Prn1, NIBSC 90/520) and FHA (NIBSC 90/654). Ampoules were completely dissolved in 500 μL of distilled water (40 µg/mL protein final concentration). Aliquots were stored at –70 ºC until use.
Protein quantification
Total protein quantification was determined with the BCA protein assay kit (Pierce/ThermoFisher, Rockford, USA), using bovine serum albumin (BSA) as calibrator. The regression coefficient (r2) was above 0.99. The concentration of purified antigens was determined by the BCA and UV absorbance method [42] using the formula: Protein (mg/mL) = 1.55 A280 – 0.76 A260.
Sandwich ELISA procedure
ELISA tests were established using a general sandwich-type procedure. Costar 3590 plates were coated with 100 µL of specific Mab or fetuin (this last for Pt-capture ELISA) in coating buffer (0.1 M carbonate/bicarbonate buffer, pH 9.6). A modification of Fetuin-capture method first described by Wong and Skelton for Pt capture ELISA [43]. Plates were coated with 4 μg of Fetuin per well, and incubated for 1 h at 37 °C. After washing three times with PBS (2.68 mM KCl, 1.47 mM KH2PO4, 136.89 mM NaCl, 8.1 mM Na2HPO4) and 0.05 % Tween 20, plates were blocked with 200 μL per well of blocking solution (PBS, 1 % BSA w/v, and 0.1 % Tween 20) for 1 h at 37 °C. Blocking solution was discarded by tapping the plate and 100 µL of test sample were added per well at the desired dilution in Sample solution (PBS, 0.1 % BSA w/v and 0.1 % Tween 20), and further incubated for 1 h at 37 °C. Calibration curves were prepared with purified proteins or NIBSC reference reagents in Sample solution at the suitable concentrations for each test. Baseline controls contained only Sample solution. After the washing step, the bound antigen was detected using 100 µL per well of a specific Mab, biotinylated or conjugated to peroxidase in Sample solution.
For ELISA setting up, tested Mabs used for detection were biotinylated and used at 10 µg/mL. Once chosen the best detection Mab for each antigen, they were conjugated and used as Mab-peroxidase conjugate. When Mab-biotin was used, after a washing step, a 1/10000 diluted streptavidin-peroxidase (streptavidin-HRP) was added followed by incubation for 30 min. After a final wash, 100 μL of Substrate solution (10 μg/mL of TMB- 3,3´,5,5´-Tetramethyl-benzidine, Cat. # Sigma-Aldrich, USA) was added per well in phosphate-citrate buffer (0.2 M phosphate, 0.1 M citrate, pH 5.0), followed by 0.006 % hydrogen peroxide and further incubated at room temperature for 10 min. The reaction was stopped by adding Stop solution (2 M H2SO4). Microtiter plates were read at a wavelength of 450 nm in a MultiScan microplate reader (SenseIdent Scan, Sweden).
Antigen was identified by considering the antigen present if absorbance was equal to or higher than 2 standard deviations (SD) of the average absorbance of baseline controls, regarded as the limit of detection (LD) of the test.
Antigen quantification was analyzed by using the GraphPad Prism 6.1 software. The mean for each duplicate or triplicate point of the calibration curve was computed and the obtained values fit to a straight line using linear regression. The best adjustment equation was determined and the regression coefficient (r2) should be equal or above 0.98. The concentration of samples was calculated by interpolation of the absorbance obtained for each sample, and further multiplied by their dilution factor.
Indirect Ptg ELISA procedure
Costar 3591 plates were coated with 100 µL of 10 µg/ mL of purified Ptg in coating buffer. The plates were incubated for 1 h at 37 °C. After washing three times with PBS and 0.05 % Tween 20, Mabs or Mab-conjugates were added at desired concentrations or dilutions, respectively, in Sample solution and were incubated for 1 h at 37 °C. After a washing step, in the case of non-conjugated Mabs, they were detected using 100 µL per well of a secondary antibody (1/10 000 dilution of mouse, Fc specific, goat peroxidase-conjugated (Cat. # A2554, Sigma-Aldrich, USA), for 30 min at 37 ºC. In both cases, after the final washing step, 100 µL per well of Substrate solution (TMB and hydrogen peroxide) was added, followed by incubation at room temperature for 10 min. The reaction was stopped by adding Stop solution. Microtiter plates were read at 450 nm. All solutions were the similar to the previously described sandwich ELISA procedure.
SDS-PAGE
Electrophoresis was performed as described [44], on 15 % gels. Culture supernatant samples were previously concentrated 10- to 18-fold by Centricon YM- 10 (Millipore, USA). Samples were boiled at 100 ºC for 10 min in the Sample buffer containing 2 % of 2-mercaptoethanol. Gels were visualized by the zinc-imidazole reverse staining technique [45]. Protein expression levels were estimated by densitometric analysis of images from SDS-PAGE stained gels.
RESULTS
Ptg-capture ELISA
Mabs 2G3 and 6D4 against Ptg were evaluated as detection reagents. In a previous western blot analysis carried out under denaturing conditions, Mab 2G3 showed reactivity against the S1 Pt subunit, whereas Mab 6D4 showed not reactivity at all (data not shown). In indirect ELISA (Ptg-coated plates), Mab 6D4 exhibited more sensitivity in ELISA (Figure 1 A and B) by using non-conjugated or peroxidase-conjugated Mab. On the other hand, when Mabs and Mab-conjugates were tested in the sandwich ELISA format (fetuin-coated plates), the outcome was completely different, with Mab 2G3 showing the best performance as detection reagent (Figure 1 C and D). Therefore, the 2G3-HRP conjugate was further used for Ptg-capture ELISA, and the titer was set up in the as 1:10 000 (results not shown).
LD of the Pt antigen using the modified Ptg-capture ELISA was determined with two-fold serial dilutions of Pt reference protein (NIBSC) at an initial concentration of 100 ng/mL (Figure 2). Under optimized ELISA conditions, the LD of the Ptg protein was 0.78 ng/mL (Figure 2A), and the linearity range of the Ptg-capture ELISA was obtained at the concentration range 0.78 to 50 ng/mL. The resulting linear equation was Y = 0.02528 X + 0.1742, with a coefficient regression coefficient (r2) above 0.99. The ELISA to identify and quantify the presence of Ptg in the supernatant of B. pertussis culture was performed using two-fold serial dilutions of Pt reference protein (NIBSC) as calibration curve. The minimum supernatant dilution (100×) having optical density (OD) value within the dynamic range was quantified as 0.055 µg/mL. The LD of the test was established as a 1/6400 dilution of the concentrated supernatant with a 0.17 OD450nm value corresponding to 0.8 ng/mL (0.052 µg/mL × 100/6400) of the Ptg protein. Furthermore, the linear formula was similar to that of Ptg (Figure 2C).
Prn-capture ELISA
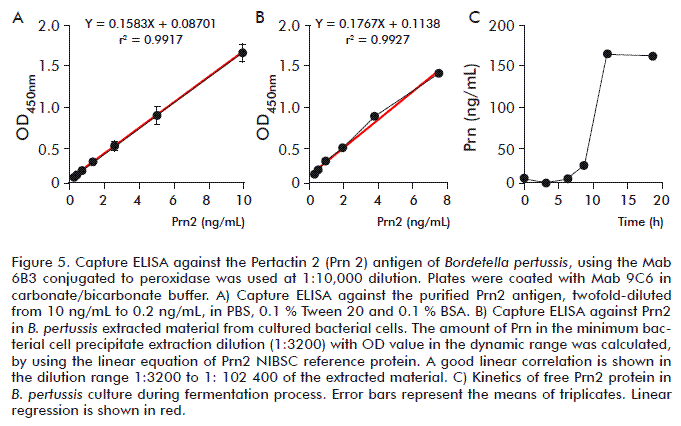
To establish the Prn capture ELISA, Mabs 9C6 and 6B3 were tested as capture or detection (biotin-conjugated) reagents. To determine the best antibodies combination, a two-fold diluted Prn1 (NIBSC reference protein) was used from 100 to 1.6 ng/mL, the best result achieved by combining Mab 9C6 as capture and 6B3 as detection antibodies (Figure 3). Mab 6B3 was conjugated to peroxidase and the optimal titer determined in ELISA as 1:10 000 (data not shown). When the recognition of Prn1 NIBSC reference protein was compared against purified Prn2 by the established Prn-capture ELISA (Figure 4), the test showed higher sensitivity for capturing Prn2 antigen, with the NIBSC antigen less recognized by the ELISA test. Then, to avoid inaccurate results, the purified Prn2 antigen was further used to prepare the calibration curve.
Under optimized Prn-capture ELISA conditions, the minimum detection of Prn2 protein was 0.07 ng/ mL and the linearity range of the ELISA was obtained at concentrations 0.15 to 10 ng/mL. The resulting linear equation was Y = 0.1583 X + 0.08701, with a linear regression coefficient (r2) above 0.99 (Figure 5A). To identify and quantify the presence of Prn2 in the B. pertussis culture, the material extracted from the heat-inactivated bacterial cell precipitate was diluted from 1:200 to 1:102 400 (Figure 5B). The lowest dilution of the extracted material (3200×) having an OD value within the dynamic range was established at 7.5 ng/ mL. The LD was a 1/102 400 dilution of the concentrated extraction material with a 0.121 OD450nm value corresponding to 0.23 ng/mL (7.5 ng/mL × 3200/102 400) of the Prn2 protein. Although the majority of Prn protein is bacterial cell surface-associated, the presence of free Prn2 was also monitored in the culture supernatant during bacterial growth (Figure 5C). Samples obtained at different time points during bacterial fermentation were evaluated by Prn-capture ELISA. At the end of the fermentation process (18 h), the free Prn2 concentration in the supernatant was quantified as 150 ng/mL. These results indicate that Prn-capture ELISA is sensitive enough for Prn2 detection in B. pertussis cultures, to monitor the expression of this protein in the culture supernatant as well as in the bacterial cell precipitate-extracted material.
FHA-capture ELISA
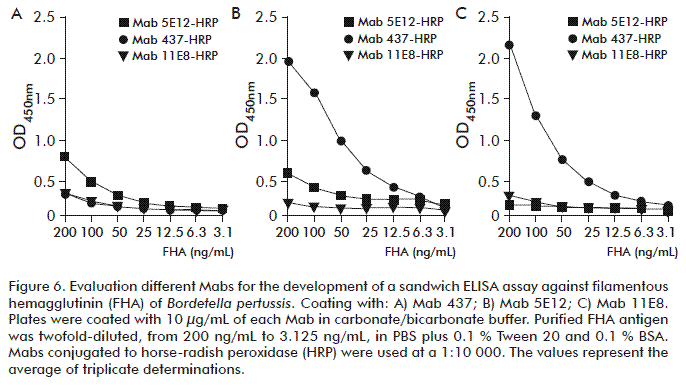
Monoclonal antibodies 11E8, 437 and 5E12 against FHA were evaluated as capture or detection reagents (peroxidase-conjugated) to establish an FHA-capture ELISA (Figure 6). The best antibodies combination was determined by using a twofold serial dilution of purified FHA in the range 200 to 3.12 ng/mL. Since FHA protein (NIBSC reference) was not available at the time of setting up the ELISA, purified FHA protein was used instead. The best ELISA results were obtained with the combination of Mab 11E8 as capture and Mab 437 peroxidase-conjugated as detection reagents (Figure 6B), with an optimal conjugate titer of 1:8000 (data not shown).
When comparing the recognition of NIBSC reference protein (FHA) and purified FHA by the established FHA-capture ELISA (Figure 7), the NIBSC antigen was poorly recognized with an average absorbance ratio (ODFHA/ODFHA NIBSC) of 5.1. Therefore, and to avoid inaccurate results, the purified FHA antigen was further used to prepare the calibration curve.
Under optimized FHA-capture ELISA conditions, the minimum detection of FHA protein was 3.25 ng/ mL and the linearity range of the ELISA was obtained in the range 3.25 to 200 ng/mL. The resulting linear equation was Y = 0.006726 X + 0.05578, with a regression coefficient (r2) above 0.99 (Figure 8A). To identify and quantify the presence of FHA in the B. pertussis culture, the supernatant was evaluated from non-diluted and a serial twofold dilution up to 1:128. The non-diluted supernatant showed OD values within the dynamic range of the assay and was quantified as 121 ng/mL. The LD was a 1/128 dilution of culture supernatant, with a 0.06 OD450nm value corresponding to 0.94 ng/mL (121 ng/mL/128) of the FHA protein.
Monitoring of B. pertussis antigens expression by ELISA
To assess the practical functionality of developed ELISA tests, three experimental cultures of B. pertussis were performed and samples were taken at three-hours intervals (0, 3, 6, 9, 12, 15 and 18 h) for monitoring the expression of Ptg, Prn2 and FHA bacterial antigens. The results (average) obtained in the three processes is shown in Figure 9. After 18 h, Ptg was detected by ELISA at 6.7 mg/L (Figure 9A). On the other hand, FHA exhibited values of 0.07 mg/L, not corresponding with the amount of protein estimated by SDS-PAGE (Figure 10, lane 1). This result suggested the underestimation of FHA by the ELISA test. The presence of aggregates was suspected and confirmed later on when a denaturing agent such as urea (2 M, final concentration) was added to the culture supernatant, the sample being further evaluated by FHA-capture ELISA. The urea-treated supernatant exhibited FHA values of approximately 8 mg/L in B. pertussis culture supernatants as determined by ELISA.
In the case of the Prn-capture ELISA, it was capable of detecting detached Prn2 in culture supernatants at a 0.15 mg/L concentration. Additionally, Prn was also quantified in extracted material since most of Prn2 was present in cells precipitate fraction (Figure 10, lane 2), the total amount of Prn2 in B. pertussis culture being estimated as 3 mg/L by ELISA.
DISCUSSION
The possibility of monitoring the expression of B. pertussis antigens during bacterial growths or to measure their quantity through purification steps, allows improving and making necessary modifications to the process to obtain vaccine antigens and to achieve more accurate recovery values. So, it is important to count with simple, specific and sensitive ELISA methods, capable of detecting and quantifying small amounts of proteins present in bacterial cell cultures and their results, which strengthened by methods like SDS-PAGE and total protein quantification, altogether, are powerful analytical tools required for any vaccine production process.
A previously reported Fetuin-capture ELISA test was modified [43] and a sandwich ELISA test developed, to identify and quantify Prn and FHA proteins from B. pertussis. In fact, ELISA-based methods have been reported for rapid screening of different antigens produced by B. pertussis, such as PT and FHA [46] or Prn and Fim [46-49]. Those methods are reliable, fast and cheap, but their results (expressed as OD) are not reproducible from culture to culture, and no antigen quantification is really made.
Ptg-capture ELISA showed good sensitivity and results, indicating that Ptg quantification in B. pertussis supernatants was consistent to Pt reference protein (NIBSC) quantification. FHA and Prn2 capture ELISA tests showed also good sensitivity, but results did not correlate with NIBSC reference antigens, most probably due to Mabs affinity, which is higher for BpCNIC0311 strain expressed antigens. Besides, the Prn expressed antigen is a pertactin type 2 variant whereas the Prn reference protein (NIBSC) is a pertactin 1. Moreover, Pt is a secreted protein, whereas Prn and FHA are surface-associated proteins [46].
Our results showed that the presence of Ptg and Prn2 in a chemically defined medium did not affect their quantification by ELISA. Moreover, the free Prn2 was quantified in the supernatant and also successfully extracted from precipitated cells without affecting its recognition by ELISA.
Nevertheless, FHA expression in the supernatant was underestimated by ELISA. In spite of putting CDX in the culture medium, FHA is a highly hydrophobic protein. Therefore, it is unstable in low salt solution and forms aggregates easily; therefore, hindering of Mabs specific epitopes is probably due to FHA-FHA or FHA-bacterial surface interactions [50] or to the conformation adopted by the expressed protein when bacteria is cultivated in vitro with chemically defined medium, or both. All these could cause blocking of the epitopes recognized by the Mab(s) used in the test. The possible presence of aggregates was corroborated when 2 M urea was added to the culture supernatant, and the sample was further evaluated by FHA-capture ELISA. At this urea molarity, the protein is not denatured and its stability and recovery in the purification steps is improved [14, 51]. So, accurate quantification of FHA in the supernatant by the established ELISA was only possible when the protein was disaggregated. Therefore, the ELISA should be used only for FHA identification in culture supernatants unless supernatants will be pre-treated with urea.
CONCLUSIONS
Easy, accurate and sensitive sandwich ELISA tests were developed, capable of identifying, quantifying or both the Pt, Prn and FHA antigens from B. pertussis in bacterial cultures. Their use in other steps throughout the vaccine antigen production process is under study.
REFERENCES
1. Bordet J, Gengou D. Le microbe de la coqueluche. Ann Inst Pasteur. 1906;20:731.
2. Black RE, Cousens S, Johnson HL, Lawn JE, Rudan I, Bassani DG, et al. Global, regional, and national causes of child mortality in 2008: a systematic analysis. Lancet. 2010;375(9730):1969-87.
3. Mata AF, Sarnaik AA. Bronchoscopy with N-acetylcysteine lavage in severe respiratory failure from pertussis infection. Pediatrics. 2013;132(5):e1418-23.
4. Edwards KM, Decker MD. Pertussis vaccines. In: Plotkin SA, Orenstein WA, Offit PA, eds. Vaccines. 6th ed. Edinburgh, Scotland: Elsevier Saunders; 2013. p. 447-92.
5. Greco D, Salmaso S, Mastrantonio P, Giuliano M, Tozzi AE, Anemona A, et al. A controlled trial of two acellular vaccines and one whole-cell vaccine against pertussis. Progetto Pertosse Working Group. N Engl J Med. 1996;334(6):341-8.
6. Gustafsson L, Hallander HO, Olin P, Reizenstein E, Storsaeter J. A controlled trial of a two-component acellular, a five-component acellular, and a whole-cell pertussis vaccine. N Engl J Med. 1996;334(6):349-55.
7. Simondon F, Preziosi MP, Yam A, Kane CT, Chabirand L, Iteman I, et al. A randomized double-blind trial comparing a two-component acellular to a whole-cell pertussis vaccine in Senegal. Vaccine. 1997;15(15):1606-12.
8. Preston A, Maskell DJ. A new era of research into Bordetella pertussis pathogenesis. J Infect. 2002;44(1):13-6.
9. Klein DL. Multicenter acellular pertussis vaccine trial: a National Institutes of Health perspective. Pediatrics. 1995;96(3 Pt 2):547-8.
10. Hewlett EL, Cherry JD. New and improved vaccines against pertussis. In: Myron ML, Graeme CW, James BK, Gary SC, editors. New generation vaccines 2nd Ed. New York: Marcel Dekker; 1997. p. 387-416.
11. Zhang L, Prietsch SO, Axelsson I, Halperin SA. Acellular vaccines for preventing whooping cough in children. Cochrane Database Syst Rev. 2011(1):CD001478.
12. Farizo KM, Burns DL, Finn TM, Gruber MF, Pratt RD. Clinical evaluation of pertussis vaccines: US Food and Drug Administration regulatory considerations. J Infect Dis. 2014;209 Suppl 1:S28-31.
13. Munoz FM, Bond NH, Maccato M, Pinell P, Hammill HA, Swamy GK, et al. Safety and immunogenicity of tetanus diphtheria and acellular pertussis (Tdap) immunization during pregnancy in mothers and infants: a randomized clinical trial. Jama. 2014;311(17):1760-9.
14. Cherry JD. The present and future control of pertussis. Clin Infect Dis. 2010;51(6):663-7.
15. van der Ark AA, Hozbor DF, Boog CJ, Metz B, van den Dobbelsteen GP, van Els CA. Resurgence of pertussis calls for re-evaluation of pertussis animal models. Expert review of vaccines. 2012;11(9):1121-37.
16. Ausiello CM, Cassone A. Acellular pertussis vaccines and pertussis resurgence: revise or replace? MBio. 2014;5(3):e01339-14.
17. Melvin JA, Scheller EV, Miller JF, Cotter PA. Bordetella pertussis pathogenesis: current and future challenges. Nat Rev Microbiol. 2014;12(4):274-88.
18. Ozcengiz E, Kilinc K, Buyuktanir O, Gunalp A. Rapid purification of pertussis toxin (PT) and filamentous hemagglutinin (FHA) by cation-exchange chromatography. Vaccine. 2004;22(11-12):1570-5.
19. Locht C, Coutte L, Mielcarek N. The ins and outs of pertussis toxin. FEBS J. 2011;278(23):4668-82.
20. Bigio M, Rossi R, Nucci D, Borri MG, Antoni G, Bartoloni A, et al. Monoclonal antibodies against pertussis toxin subunits. FEMS Microbiol Lett. 1988;51:7-12.
21. Sato H, Ito A, Chiba J, Sato Y. Monoclonal antibody against pertussis toxin: effect on toxin activity and pertussis infections. Infect Immun. 1984;46(2):422-8.
22. Sato H, Sato Y. Protective activities in mice of monoclonal antibodies against pertussis toxin. Infect Immun. 1990;58(10):3369-74.
23. Locht C. Molecular aspects of Bordetella pertussis pathogenesis. Int Microbiol. 1999;2(3):137-44.
24. Robbins JB, Schneerson R, Kubler-Kielb J, Keith JM, Trollfors B, Vinogradov E, et al. Toward a new vaccine for pertussis. Proc Natl Acad Sci USA. 2014;111(9):3213-6.
25. Henderson IR, Navarro-Garcia F, Nataro JP. The great escape: structure and function of the autotransporter proteins. Trends Microbiol. 1998;6(9):370-8.
26. Hijnen M, van Gageldonk PG, Berbers GA, van Woerkom T, Mooi FR. The Bordetella pertussis virulence factor P.69 pertactin retains its immunological properties after overproduction in Escherichia coli. Protein Expr Purif. 2005;41(1):106-12.
27. Leininger E, Ewanowich CA, Bhargava A, Peppler MS, Kenimer JG, Brennan MJ. Comparative roles of the Arg-Gly-Asp sequence present in the Bordetella pertussis adhesins pertactin and filamentous hemagglutinin. Infect Immun. 1992;60(6):2380-5.
28. Leininger E, Roberts M, Kenimer JG, Charles IG, Fairweather N, Novotny P, et al. Pertactin, an Arg-Gly-Asp-containing Bordetella pertussis surface protein that promotes adherence of mammalian cells. Proc Natl Acad Sci USA. 1991;88(2):345-9.
29. He Q, Makinen J, Berbers G, Mooi FR, Viljanen MK, Arvilommi H, et al. Bordetella pertussis protein pertactin induces type-specific antibodies: one possible explanation for the emergence of antigenic variants? J Infect Dis. 2003;187(8):1200-5.
30. Locht C, Bertin P, Menozzi FD, Renauld G. The filamentous haemagglutinin, a multifaceted adhesion produced by virulent Bordetella spp. Mol Microbiol. 1993;9(4):653-60.
31. Makhov AM, Hannah JH, Brennan MJ, Trus BL, Kocsis E, Conway JF, et al. Filamentous hemagglutinin of Bordetella pertussis. A bacterial adhesin formed as a 50-nm monomeric rigid rod based on a 19-residue repeat motif rich in beta strands and turns. J Mol Biol. 1994;241(1):110-24.
32. Arico B, Nuti S, Scarlato V, Rappuoli R. Adhesion of Bordetella pertussis to eukaryotic cells requires a time-dependent export and maturation of filamentous hemagglutinin. Proc Natl Acad Sci USA. 1993;90(19):9204-8.
33. Jacob-Dubuisson F, Buisine C, Mielcarek N, Clement E, Menozzi FD, Locht C. Amino-terminal maturation of the Bordetella pertussis filamentous haemagglutinin. Mol Microbiol. 1996;19(1):65-78.
34. Renauld-Mongenie G, Cornette J, Mielcarek N, Menozzi FD, Locht C. Distinct roles of the N-terminal and C-terminal precursor domains in the biogenesis of the Bordetella pertussis filamentous hemagglutinin. J Bacteriol. 1996;178(4):1053-60.
35. Brennan MJ, Burns DL, Meade BD, Shahin RD, Manclark CR. Recent advances in the development of pertussis vaccines. In: Ellis R, editor. Vaccines: new approaches to immunological problems. Shoreham, Mass: Butterworth; p. 23-52.
36. Brennan MJ, Shahin RD. Pertussis antigens that abrogate bacterial adherence and elicit immunity. Am J Respir Crit Care Med. 1996;154(4 Pt 2):S145-9.
37. Cherry JD. 2012. Why do pertussis vaccines fail? Pediatrics. 129:968-70.
38. Plotkin SA. The pertussis problem. Clin Infect Dis. 2014;58(6):830-3.
39. Reed G, Galindo MA. Cuba’s National Immunization Program. MEDICC Rev. 2007;9(1):5-7.
40. Quintana-Vázquez D, Delgado M, Lemos G, Coizeau E, Ramos Y, Támbara Y, et al. Assessment of the Bordetella pertussis BpCNIC0311 STRAIN as a producing strain of genetically detoxified toxoid (PTg), filamentous hemagglutinin (FHA) and type 2 Pertactin (Prn2). J Infect Dis Ther. 2015;3:8-20.
41. Thalen M, van der Ark A, van den Ijssel J, van Straaten I, Jansen D, Beuvery C, et al. Improving the cellular pertussis vaccine: increased potency and consistency. Vaccine. 2008;26(5):653-63.
42. Layne E. Spectrophotometric and turbidimetric methods for measuring proteins. Methods Enzymol. 1957;3:447-54.
43. Wong KH, Skelton SK. New, practical approach to detecting antibody to pertussis toxin for public health and clinical laboratories. J Clin Microbiol. 1988;26(7):1316-20.
44. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227(5259):680-5.
45. Fernandez-Patron C, Castellanos-Serra L, Rodriguez P. Reverse staining of sodium dodecyl sulfate polyacrylamide gels by imidazole-zinc salts: sensitive detection of unmodified proteins. Biotechniques. 1992;12(4):564-73.
46. Barkoff AM, Guiso N, Guillot S, Xing D, Markey K, Berbers G, et al. A rapid ELISA-based method for screening Bordetella pertussis strain production of antigens included in current acellular pertussis vaccines. J Immunol Methods. 2014;408:142-8.
47. Tsang RS, Sill ML, Advani A, Xing D, Newland P, Hallander H. Use of monoclonal antibodies to serotype Bordetella pertussis isolates: comparison of results obtained by indirect whole-cell enzyme-linked immunosorbent assay and bacterial microagglutination methods. J Clin Microbiol. 2005;43(5):2449-51.
48. Heikkinen E, Xing DK, Olander RM, Hytonen J, Viljanen MK, Mertsola J, et al. Bordetella pertussis isolates in Finland: serotype and fimbrial expression. BMC Microbiol. 2008;8:162.
49. Barkoff AM, Mertsola J, Guillot S, Guiso N, Berbers G, He Q. Appearance of Bordetella pertussis strains not expressing the vaccine antigen pertactin in Finland. Clin Vaccine Immunol. 2012;19(10):1703-4.
50. Menozzi FD, Boucher PE, Riveau G, Gantiez C, Locht C. Surface-associated filamentous hemagglutinin induces autoagglutination of Bordetella pertussis. Infect Immun. 1994;62(10):4261-9.
51. Wu T, Bi J, Huang Y, Zhang Y, Sun L, Sun C, et al. Effect of solution environment on the purification of pertussis toxin. Sheng Wu Gong Cheng Xue Bao. 2008;24(7):1279-84.
Received in January, 2015.
Accepted in July, 2015.
Gilda Lemos-Pérez. Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba. E-mail: gilda.lemos@cigb.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}