










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.33 no.4 La Habana oct.-dic. 2016
RESEARCH
Changes in peptide and protein concentrations during the ontogenesis of honeybee (A. mellifera) drone larvae is associated to variations in protease activity
Cambios en las concentraciones de péptidos y proteínas durante la ontogénesis de larvas de zángano de abeja (A. mellifera) asociados con variaciones en la actividad proteolítica
Zhanna Grishina, Michael Gengin†
Penza State University, Penza, 440026, Russia.
ABSTRACT
The development of a bee drone larvae comprises a period of seven days, in which the larva mass increases 1500-fold. There is no comparable increase during the development of any other species, the in metabolic activity is not observed in any other species development. Rapid development of larvae of insects is mediated by key regulators such as regulatory peptides or protease activity acting on those peptides. Therefore, we studied the dynamics of peptide turn-over during larvae ontogenesis by purifying and measuring the peptide amount in larvae, depending on their age, using size-exclusion chromatography and spectrophotometric measurement. Additionally, we measured the activity of proteolytic enzymes such as a cathepsin D and the trypsin-like proteases, by measuring the conversion rate of enzyme specific substrates, and the effect of pH on the enzyme activity was studied using specific substrates, to corroborate the differences to be found in the studied proteolytic enzymes. We demonstrated correlation between the activity of trypsin-like proteases and peptide amount during larvae ontogenesis. This could indicate that these enzymes participate in the production and cleavage of regulatory peptides during larvae ontogenesis, whereas no correlation was observed for cathepsin D activity and peptide amount.
Keywords: honeybee, drone larvae, ontogenesis, peptides, cathepsin D, trypsin-like proteases.
RESUMEN
La larva de zángano de abeja se desarrolla en un periodo de siete días, durante el cual la masa de la larva aumenta mil quinientas veces. Tal aumento en la actividad metabólica es único entre las especies del Reino Animal. El rápido desarrollo de las larvas de los insectos está mediado por reguladores clave, como son los péptidos reguladores o las diferentes actividades de proteasas que actúan sobre dichos péptidos. Por tales razones, en este trabajo se estudió la dinámica del recambio de péptidos durante la ontogénesis de las larvas de zángano de abeja, mediante la purificación y la medición de los niveles de péptidos en diferentes edades del desarrollo de las larvas. Para ello se empleó la cromatografía de exclusión molecular y la espectrofotometría. También se midió la actividad de enzimas proteolíticas como la catepsina D y las proteasas similares a tripsina, a las que se les evaluó la tasa de conversión de los sustratos específicos. Además, se evaluó el efecto del pH sobre la actividad de estas enzimas proteolíticas, con el uso de sustratos específicos. Se demostró la correlación existente entre la actividad de las proteasas similares a tripsina y los niveles de péptidos durante la ontogénesis. Estos resultados pueden indicar que dichas enzimas participan en la producción y clivaje de los péptidos reguladores durante la ontogénesis de las larvas de zángano de abeja. No se observó dicha correlación entre los niveles de péptidos y la actividad catepsina D.
Palabras clave: abeja mielífera, larva de zángano, ontogénesis, péptidos, catepsina D, proteasas similares a tripsina.
INTRODUCTION
The evolutionary development of organisms is mediated by relevant regulatory processes and mechanisms during the different ontogenesis stages. One of such processes is honeybee drone larvae development, in which the drone increases its body mass 1500-fold in just 7 days, what poses a huge metabolic pressure, not paralleled in the rest of the Animal kingdom [1]. Among the said regulatory processes involved, the control exerted by proteolytic enzymes on protein turnover during development is essential, which could be particularly relevant in honeybee drone larvae, as in other living organism.
All structural proteins, enzymes and peptides with numerous physiological actions are substrates of proteolytic enzymes in a varying degree [2-5]. Proteases are not only responsible for protein degradation for the aim of protein de novo synthesis, but it is also considered as a major metabolism regulatory factor. In fact, the hydrolysis of a peptide bond in an active or inactive protein can lead to a protein or peptide with new properties [6].
Recently, the regulatory role of peptides in the regulation of ontogenesis was recognized [7]. Peptides can act on the re-organization of proteins during phases of development of an organism including tissue and organ formation. According to the modern idea, regulatory action of peptides in an organism of animals is carried out by particular enzymatic systems [8-10].
Our previous work focused on the ontogeny of honeybees because different members of this family spend different periods for embryonal and post-embryonal development, and differences in their life expectancy were shown in spite of carrying an identical genome [11]. However, the mechanisms mediating those differences remained to be characterized. Therefore, the aim of this work was to study the turnover of regulatory peptides during the development of honeybee drone larvae, as measuring the activity of proteolytic enzymes such as a cathepsin D and the trypsin-like proteases. Our results evidence a possible direct link between proteolytic activity and regulatory peptides during the development of honeybee drone larvae.
MATERIALS AND METHODS
Reagents
Pepstatin A and hemoglobin (Methemoglobin) substrate, Sephadex G-25, Kunitz STI protease inhibitor, trichloroacetic acid (TCA), NaCl, HCl and sodium acetate were from Sigma-Aldrich (USA). Nα-Benzoyl- D,L-arginine 4-nitroanilide hydrochloride (BAPNA) was from Sigma (Germany).
Samples
The experimental material was a larvae drone and worker of the A. mellifera, collected in the second half of May-June 2016. The brood originated from a 30-colony private apiary of Dr. Michael Gengin located 50 km outside Penza. To acquire larvae of known ages, a queen was isolated on an empty frame with honey and pollen for 16 h inside a nucleus colony with several hundred worker bees. The brood frame with newly laid eggs was then placed into the original hive, along with the queen, workers, and two supporting frames. Three days after reintroducing the eggs into the colonies, larvae were collected for five consecutive days. Before proceeding with protein collection, all larvae were washed in 0.9 % NaCl to reduce royal jelly contamination.
Thereafter, the larvae was isolated from honeycombs and separated into developmental stages based on morphological features according to Jay (1962, 1963) [12]: one, two, three, four, five and six-seven days-old drones larvae and one to six days-old workers larvae.
Sample preparation
For each time-point, 1 g larvae was macerated in a glass homogenizer and resuspended in 10 mL of 0.9 % NaCl (10 %) for protein extraction, and centrifuged at 10 000 g for 30 min to remove cell debris. Then, the supernatant was used for further gel-filtration to obtain the fraction of peptides (Mr ≤ 10 kDa) and proteins (Mr ≥ 10 kDa). Protein concentration was measured by Lowry method [13]. All procedures were performed on ice and the extract was stored at –40 °C. All the experiments were run using 4-6 homogenates for each stage of larval development. Homogenates were analyzed independently.
Size-exclusion chromatography fractionation of extracts
Proteins and peptides were fractionated from the initial 10 % extract by gel-filtration, using a 35 × 1.5-cm column filled with Sephadex G-25 (Sigma-Aldrich, USA): 400-500 μL of protein solutions with concentration of 4-5 mg/mL in 0.9 % of NaCl were used for gel-filtration. Proteins were eluted from the column at a 30 mL/h flow rate and fractions were collected in a final volume of 2 mL. Concentration of the eluted proteins was measured spectrophotometrically at a wavelength of 280 nm.
Enzyme assay
The cathepsin D assay for cathepsin activity measurement was carried out according to Barrett’s method with the following modifications: 40 μL of 8 % hemoglobin solution was mixed with 120 μL of 100 mM sodium acetate buffer pH 4.5, to which 40 μL of the enzyme (~20 μg protein) was added and incubated for 40 min at 37 °C [14]. Then 200 μL of 5 % TCA was added and the samples were centrifuged for 20 min at 4000 g. Afterwards, 100 μL of supernatant were collected, containing the TCA soluble peptide fraction. Hemoglobin hydrolysates were quantified using the Lowry protein assay. The activity of the enzyme was expressed as the amount of μmol of tyrosine formed during 40 min incubation at 37 °C, estimated using 1 mg of protein. Activity was measured by determining the optical density (O.D.) at 750 nm.
To determine the enzyme activity of trypsin-like proteases, 0.5 mL BAPNA and 0.05 M of Phosphate buffer (pH 10) was added to 250 μL of each sample and incubated at 37 °C for 30 min. Subsequently, the reaction was stopped with 350 μL of 1 M HCl. The supernatant containing the peptides was collected and O.D. was determined at 383 nm against blank. The enzymatic activity was expressed as the amount of μmol of p-nitroaniline formed during 30 min of incubation at 37 °C, estimated using 1 mg of protein.
All enzyme assays were analyzed in triplicates, the average values and standard deviations are presented.
Analysis of enzyme optimal pH and activity
To determine the pH optimum of cathepsin D, enzymes containing samples were incubated with hemoglobin in different buffers of the pH range 3.0–5.0, and the activities were measured as described above. To determine the pH optimum of trypsin-like proteases, enzymes containing samples was incubated with BAPNA in different buffers of the pH range 7.0-11.0, and the enzymatic activities were measured as described above.
Inhibition analysis was performed using specific inhibitors of proteases: pepstatin (0.1 mg/mL) for cathepsin D and Kunitz STI protease inhibitor (0.1 mg/ mL) for trypsin-like proteases.
Statistical analysis
The experiment was done in three replicates, each containing four to six samples (all stages of development) of the studied stages. Data was expressed as average values ± standard deviations. ANOVA (single-tailed) and the Student’s t-test were employed to evaluate the differences between means of the enzymatic activities (p < 0.05) [15]. For identification of interrelation between activity of enzymes and amount of proteins and peptides correlation analysis was carried out (Pearson’s test). All statistic estimations were done by using the Microsoft Exel-2007 software.
RESULTS AND DISCUSSION
The fast development of insect larvae is mediated by a highly active metabolism, of which key regulators are peptides [16], their levels defined by the activity of proteases. Therefore, we studied the dynamics of peptide turn-over during larvae ontogenesis in honeybee drone larvae and the activity of proteolytic enzymes such as a cathepsin D and the trypsin-like proteases. The effect of pH on the enzyme activity was studied using specific substrates to confirm the difference in the studied proteolytic enzymes [17-20]. It has been described that the pH for the optimum enzymatic activity of Capthesin D and trypsin-like proteases differs between vertebrate animals and insects [5, 21]. As far as we know, the optimum pH of honeybee Capthesin D and trypsin-like proteases remained unknown. Therefore, the effect of pH on the enzymatic activity of these enzymes was studied using specific substrates. It was found that the optimum pH for cathepsin D was pH 4.5, and pH 10.1 for trypsin-like proteases (Figure 1).
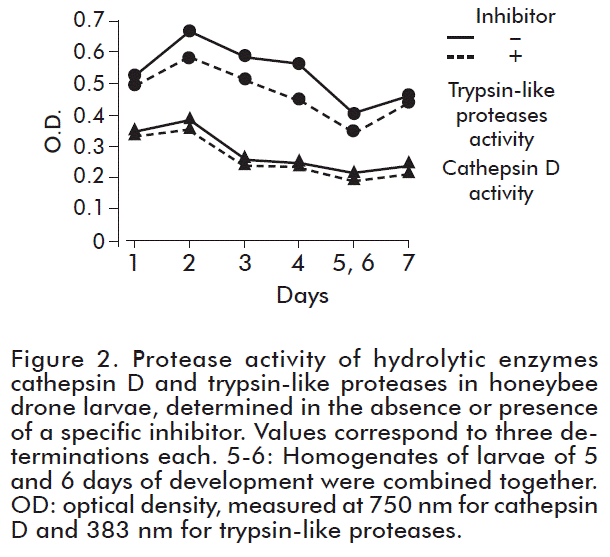
Additionally, we analyzed the protease class using specific inhibitors for proteolytic enzymes. These substrates (pepstatin for cathepsin D and Kunitz STI protease inhibitor for trypsin-like proteases) inhibited the activity of the enzymes in 7-10 % (Figure 2). There was a low inhibition rate because the proteases in insects show different substrate specificity [3, 5, 19].
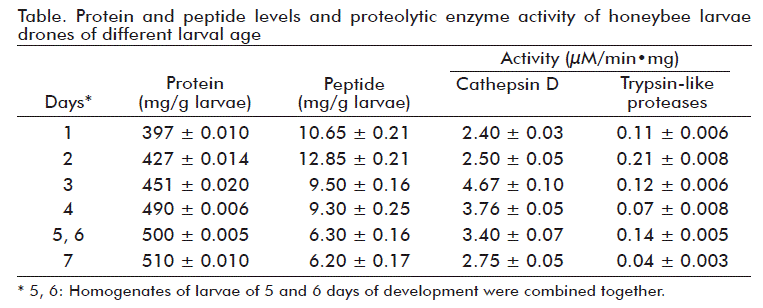
Results of protein and peptide concentration, and enzyme activity in larvae extracts, are presented in the table. There was observed an increase in protein concentration along with the development stages of drone larvae. Dynamics of peptides during ontogenesis of larvae differs from dynamics of proteins. The maximal abundance of peptides in larvae was detected on day 2 (Table).
On day 1, 1 g of larvae contained an average of 397 mg of protein, whereas, on day 4, protein content reached 490 mg. In contrast, the maximal abundance of peptides is observed on day 2 (12.9 mg) of larvae development. Its minimum was reached after 4-5 days (6.3 mg), which is 2-fold less than the amount on day 2. Thus, early stages of larvae development include protein biosynthetic processes, which are regulated by the turn-over of regulatory peptides [2, 8-11].
The dynamics of cathepsin D and trypsin-like protease activity at different stages of larvae development is presented in Figure 3. Activity of cathepsin D reaches a maximum on day 3 and decreases subsequently in larvae with their age (Table). In contrast, the activity of trypsin-like proteases shows its maximum on day 2 followed by an inconsistent pattern of lower enzyme activity.
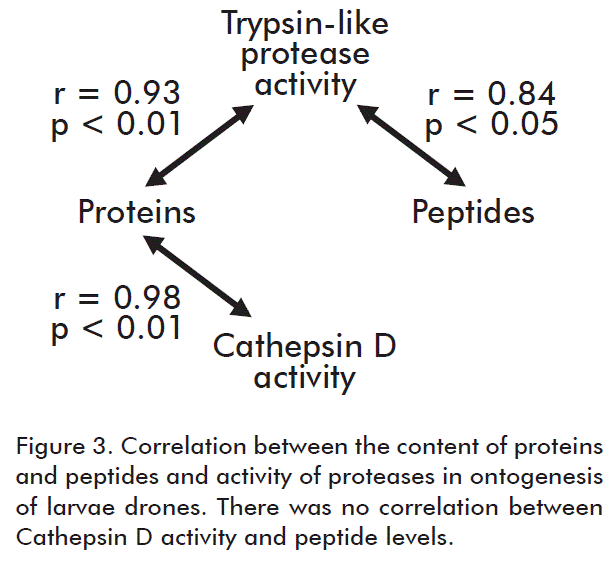
According to the correlation analysis, we demonstrated a correlation between the activity of trypsin-like proteases and peptide amount during ontogenesis of larvae (Table). Therefore, we assumed that these enzymes participate in production and cleavage of regulatory peptides. Besides, correlation between the activity of trypsin-like proteases and concentration of proteins is shown (Figure 3), indicative of their activity during protein metabolism in insects. This dependence has been observed also in vertebrate animals [2, 4, 19- 22]. No reliable correlation of cathepsin D activity and peptides was observed (Figure 3) [15, 21].
Cathepsin D is known to proteolytically cleave and modify lysosomal proteins. This coincides with the correlation observed in this study between cathepsin D activity and protein amounts, and the lack of correlation with the concentration of peptides. Trypsin-like proteases are commonly active in the cytoplasm and their activity correlates with protein as well as peptide amounts in larvae. Thus, we propose that trypsin-like proteases are involved in the synthesis of regulatory peptides whereas cathepsin D is not connected with this process. Regulatory peptides have their function in the cytoplasm during biosynthesis of regulatory protein needed during early stages of larvae development [2, 8-11].
Notably, these enzymes usually participate in the degradation of proteins to free amino acids, contributing too homeostasis of proteins and peptides with functions other than regulation [2, 4, 5, 19, 21, 23].
CONCLUSIONS
Studying the dynamics of regulatory peptides and proteolytic enzymes during ontogenesis of larvae drones opens the view on evolutionary mechanisms in bee families. Our data reveals the correlation between the activity of our studied enzymes and amount of proteins or peptides, thus, confirming the functional dependence of proteolytic enzymes and cellular substrates.
By studying proteolytic activity with specific substrates we examined the dynamics in the activity change of cathepsin D and the trypsin-like proteases in ontogenesis of larvae drone. The correlation of cathepsin D activity and protein during ontogenesis indicates an important role of cathepsin D in protein metabolism regarding the degradation of spare proteins as well as the catabolism to deliver structural proteins for building the developing larva organism. Due to the correlation of trypsin-like protease activity and peptide dynamics, participation of these proteases in the synthesis of regulatory peptides is suggested. Thus, trypsin-like proteases have regulatory function during ontogenesis of larva drone additionally to their role during protein homeostasis.
REFERENCES
1. Chan QW, Foster LJ. Changes in protein expression during honey bee larval development. Genome Biol. 2008;9(10):R156.
2. Gengin M. Features structural - functional organization and physicochemical properties not lysosomal peptide-hydrolases of animal brain (PhD Thesis). Penza: Penza University; 2002.
3. Zótowska K, Lipinski Z, Farjan M. Activity of selected hydrolases in ontogeny of drone Apis mellifera carnica. J Apicult Sci. 2011;51(1):95-100.
4. Zótowska K, Fraczek R, Lipinski Z. Hydrolases of developing worker brood and newly emerged worker of Apis mellifera carnica. J Apicult Sci. 2011;51(1):27-37.
5. Iarygin DV, Min’kova NO, Filippovich Iu B. Study of intracellular localization of the proteolytic enzyme complex and its protein inhibitors in bombyx grain. Ontogenez. 2012;43(5):325-32.
6. Gomazkov OA. Functional biochemistry of regulatory peptides. Moscow: Nauka; 1993.
7. Fang Y, Feng M, Han B, Qi Y, Hu H, Fan P, et al. Proteome Analysis Unravels Mechanism Underling the Embryogenesis of the Honeybee Drone and Its Divergence with the Worker (Apis mellifera lingustica). J Proteome Res. 2015;14(9):4059-71.
8. Nassel DR, Homberg U. Neuropeptides in interneurons of the insect brain. Cell Tissue Res. 2006;326(1):1-24.
9. Havinson V, Kvetnaja T. Regulatory peptides and homeostasis. Russ Chem J. 2005;XLIX(1):112-7.
10. Ashmarin IP, Obukhova MF. Regulatory peptides. A functionally continuous collection. J Biochem. 1986;51(4):531-42.
11. Mishukovskaya G. Ultrastructural and histochemical changes in the isolated muscle of a flying musculation of bees during ontogenesis. Bee J. 2007; 7:18-20.
12. Jay SC. The Development of Honeybees in their Cells. Journal of Apicultural Research. 1963;2(2):117-34.
13. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951;193(1):265-75.
14. Barrett AJ. Cathepsin D. Purification of isoenzymes from human and chicken liver. Biochem J. 1970;117(3):601-7.
15. Ivanter EV, Korosov AV. Elementary Biometrics: A Tutorial. Petrozavodsk: Izd. PetrGU; 2010.
16. Crailsheim K. The protein balance of the honeybee worker. Apidologie. 1990;21: 417-29.
17. Blanco-Labra A, Martinez-Gallardo NA, Sandoval-Cardoso L, Delano-Frier J. Purification and characterization of a digestive cathepsin D proteinase isolated from Tribolium castaneum larvae (Herbst). Insect Biochem Mol Biol. 1996;26(1):95-100.
18. Saikhedkar N, Summanwar A, Joshi R, Giri A. Cathepsins of lepidopteran insects: Aspects and prospects. Insect Biochem Mol Biol. 2015;64:51-9.
19. Wielkopolan B, Walczak F, Podlesny A, Nawrot R, Obrepalska-Steplowska A. Identification and partial characterization of proteases in larval preparations of the cereal leaf beetle (Oulema melanopus, Chrysomelidae, Coleoptera). Arch Insect Biochem Physiol. 2015;88(3):192-202.
20. Shrimpton CN, Smith AI. Soluble neutral metallopeptidases: physiological regulators of peptide action. J Pept Sci. 2000;6(6):251-63.
21. Gui ZZ, Lee KS, Kim BY, Choi YS, Wei YD, Choo YM, et al. Functional role of aspartic proteinase cathepsin D in insect metamorphosis. BMC Dev Biol. 2006;6:49.
22. Volpicella M, Ceci LR, Cordewener J, America T, Gallerani R, Bode W, et al. Properties of purified gut trypsin from Helicoverpa zea, adapted to proteinase inhibitors. Eur J Biochem. 2003;270(1):10-9.
23. Fusek M, Vetvicka V. Dual role of cathepsin D: ligand and protease. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2005;149(1):43-50.
Received in October, 2016.
Accepted in December, 2016.
Zhanna Grishina. Penza State University, Penza, 440026, Russia. E-mail: grinzanetk@icloud.com.


{kind=link}
{kind=link}
{kind=link}
{kind=link}