










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Plantas Medicinales
versión On-line ISSN 1028-4796
Rev Cubana Plant Med v.16 n.1 Ciudad de la Habana ene.-mar. 2011
ARTÍCULO ORIGINAL
Efecto protector de los polifenoles de Rhizophora mangle L. sobre el daño oxidativo a proteínas y ADN
Protective effect of polyphenols from Rhizophora mangle L. on oxidative damage to proteins and DNA
Janet Sánchez Calero,I Gregorio Martínez Sánchez,II Roberto Faure GarcíaIII
I Doctora en Medicina Veterinaria. Investigadora Agregada. Departamento de Química-Farmacología-Toxicología, Centro Nacional de Sanidad Agropecuaria. San José de Las Lajas, La Habana, Cuba.
II Doctor en Ciencias Farmacéuticas. Investigador Auxiliar. Centro de Investigaciones y Evaluaciones Biológicas, Instituto de Farmacia y Alimentos. Lisa, La Habana, Cuba.
III Doctor en Medicina Veterinaria. Investigador Titular. Departamento de Química-Farmacología-Toxicología, Centro Nacional de Sanidad Agropecuaria. San José de Las Lajas, La Habana, Cuba.
RESUMEN
Introducción: al extracto acuoso de la corteza de Rhizophora mangle L. se le ha demostrado un amplio espectro de usos medicinales: en el tratamiento de la mastitis bovina, la curación de heridas, las infecciones uterinas y las úlceras gastroduodenales; debido a sus propiedades antiséptica, cicatrizante, antiinflamatoria y antioxidante. Sin embargo, no se han completado los estudios de la actividad antioxidante a todos los niveles de complejidad para dilucidar los mecanismos de acción involucrados en este efecto farmacológico.
Objetivo: determinar si el extracto acuoso de R. mangle y su fracción polifenólica protegen a las principales biomoléculas del daño oxidativo.
Metodos: la evaluación de la actividad antioxidante del extracto de R. mangle y su fracción polifenólica sobre las principales biomoléculas se determinó mediante ensayo de daño oxidativo a la albúmina de suero bovino expuesta a los radicales hidroxilo generados en el sistema Fenton y ensayo de degradación oxidativa del ADN inducido por el sistema bleomicina-Fe3+.
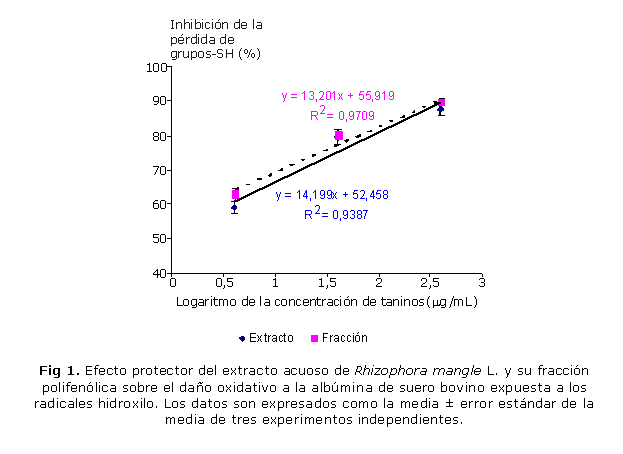
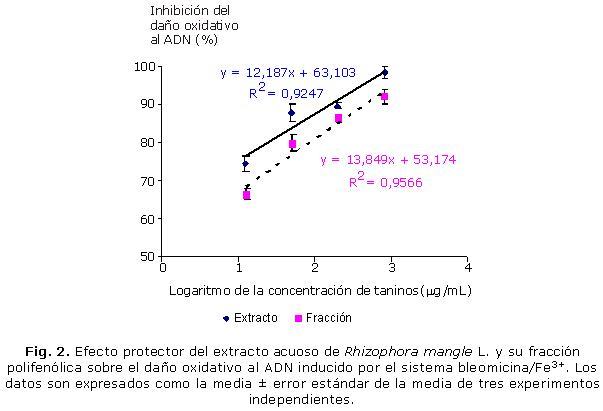
Resultados: el extracto de R. mangle y su fracción polifenólica, a la máxima concentración ensayada, disminuyeron la oxidación de los grupos sulfidrilos en 87,3 y 89,1 %; e inhibieron la degradación del ADN en 98,4 y 91,9 %, respectivamente. El análisis de regresión mostró que ambos efectos fueron dependientes de la concentración de taninos en el extracto y su fracción. La comparación de las líneas de regresión reveló que el extracto y su fracción resultaron igualmente eficaces en proteger a la albúmina de suero bovino de la oxidación por los radicales hidroxilos, sin embargo, el extracto fue más eficaz en proteger al ADN de la degradación oxidativa que su fracción.
Conclusiones: el extracto acuoso de R. mangle mostró un efecto protector a las principales biomoléculas del daño oxidativo, evidenciado por la inhibición de la pérdida de grupos sulfidrilos en la albúmina de suero bovino y la disminución de la degradación del ADN. A su vez se demostró que los compuestos polifenólicos presentes en el extracto fueron los principales responsables de los efectos antioxidantes observados en este estudio.
Palabras clave: radicales hidroxilo, bleomicina, biomoléculas, compuestos polifenólicos, Rhizophora mangle (L.), actividad antioxidante.
ABSTRACT
Introduction: the aqueous extract from Rhizophora mangle (L.) bark has demonstrated a broad spectrum of medicinal uses; for example, in treating bovine mastitis, wound healing, uterine infections and gastroduodenal ulcers, due to its antiseptic, healing, anti-inflammatory and antioxidant properties. However, the antioxidant activity in its whole complexity has not been fully studied in order to elucidate the mechanisms of action involved in this pharmacological effect.
Objective: to determine if R. mangle bark aqueous extract and its polyphenolic fraction protect the main biomolecules from oxidative damage.
Methods: the antioxidant activity of R. mangle extract and its polyphenolic fraction on the main biomolecules was determined by the following methods: oxidative damage trial on bovine serum albumin exposed to hydroxyl radicals generated in the Fenton system and the bleomycin-Fe3+ system-induced DNA oxidative degradation trial.
Results: the R. mangle bark extract and its polyphenolic fraction, at the highest tested concentration, reduced the sulfhydryl group oxidation by 87,3 % and 89,1 % and they also inhibited the DNA degradation by 98.4 % and 91.9 % respectively. The regression analysis demonstrated that both effects depended on tannin concentration in the extract and its fraction. The comparison of regression lines revealed that the extract and its fraction were equally effective in protecting bovine serum albumin from oxidation by hydroxyl radicals; however, the extract was more effective when protecting DNA from oxidative degradation than its fraction.
Conclusions: the R. mangle aqueous extract showed a protective effect on the main biomolecules from the oxidative damage, evidenced by inhibiting loss of sulfhydryl group in bovine serum albumin and decreasing the DNA degradation. At the same time, it was shown that polyphenolic compounds present in the extract were the main responsible for the antioxidant effects observed in this study.
Key words: hydroxyl radicals, bleomycin, biomolecules, polyphenolic compounds, Rhizophora mangle (L.), antioxidant activity.
INTRODUCCIÓN
El estrés oxidativo surge en sistemas biológicos después de una prolongada exposición a oxidantes, o debido a una disminución de la capacidad antioxidante del sistema; y está asociado con la generación de especies reactivas del oxígeno (ERO), las cuales están implicadas en la patología de numerosas enfermedades como el cáncer, enfermedades cardiacas, arterosclerosis, enfermedades cerebrales y el envejecimiento.1 Cuando un exceso de ERO se forma, puede causar efectos letales en las células por la oxidación de lípidos, proteínas, ADN y carbohidratos; que ocasionan alteraciones en sus funciones biológicas.2 Sin embargo, los compuestos antioxidantes tienen la capacidad de inhibir o interrumpir las reacciones de oxidación que causan daños a las mencionadas biomoléculas.3
Varios estudios han demostrado que las plantas producen potentes antioxidantes y representan una importante fuente de antioxidantes naturales. En tal sentido, la caracterización química del extracto acuoso de la corteza de Rhizophora mangle L. (mangle rojo) reveló la presencia de polifenoles (54,78 %), representados en su mayoría por taninos poliméricos (80 %) y taninos hidrolizables (20 %); en estos últimos se destaca la presencia de epicatequina, catequina, ácido clorogénico, ácido gálico y ácido elágico, además se encontraron galotaninos y elagitaninos. De las estructuras no tánicas, se refiere la presencia de carbohidratos (17,5 %) libres y enlazados; ácidos grasos (4,0 %) de cadena larga, saturados e insaturados; fitoesteroles (0,0285 %); componentes volátiles o semivolátiles (70 compuestos) (0,0205 %) y aromas o aceites esenciales no volátiles.4
A esta planta se le atribuyen varias propiedades en medicina tradicional. Se ha utilizado para el tratamiento de enfermedades de la garganta y en la tuberculosis pulmonar;5 así como contra la lepra, el asma, el envenenamiento con pescados contaminados, la úlcera péptica, los trastornos digestivos, las infecciones de la piel y las enfermedades venéreas.6 Además se reportó que tiene actividad antifúngica.7
En la última década se demostraron varias propiedades farmacológicas del extracto acuoso de la corteza de R. mangle, que incluyen, prevención de la mastitis bovina,8 eficacia en la curación de las heridas,9,10 así como propiedades antimicrobianas;11,12 a su vez resultó exitoso en el tratamiento de las infecciones uterinas13 y las úlceras gastroduodenales,14 también se demostraron sus propiedades antiinflamatorias.15
Algunas propiedades antioxidantes han sido demostradas previamente, tanto para el extracto acuoso de R. mangle como para su fracción mayoritaria constituida por compuestos polifenólicos, como son: actividad secuestradora de radicales hidroxilo, propiedades quelantes de iones de hierro,16 inhibición de la peroxidación lipídica en cerebro de rata, reducción del riesgo de hemólisis inducida por radicales libres;17 y en un modelo de curación de heridas abiertas asépticas en ratas, durante el proceso de reparación de las lesiones, incrementó los indicadores antioxidantes: superóxido dismutasa, catalasa, glutatión reducido y redujo los biomarcadores prooxidantes: malonildialdehído.18 Aún así no existe información acerca de sus efectos sobre otras biomoléculas, como: ADN y proteínas, expuestas a daño oxidativo, un aspecto que es importante conocer para completar el estudio de las propiedades antioxidantes de esta planta a todos los niveles de complejidad y dilucidar los mecanismos de acción involucrados en este efecto farmacológico. De ahí que el objetivo del presente trabajo consistió en determinar si el extracto acuoso de R. mangle y su fracción polifenólica protegen a las principales biomoléculas del daño oxidativo.
MÉTODOS
Material vegetal
Las cortezas de Rhizophora mangle L. se colectaron en la zona de la Playa Caimito, La Habana, en junio de 2007. A un espécimen se le realizó la identificación taxonómica en el Herbario del Jardín Botánico Nacional (La Habana, Cuba).
Preparación del extracto
El material vegetal fresco se secó al aire, a temperatura ambiente (25 °C). El extracto de R. mangle se obtuvo a partir de la decocción acuosa de las cortezas previamente molinadas, en una proporción 1:7,5 (p/v) durante 30 min a 95 °C. El extracto se concentró hasta 24 mg/mL de materia seca y mostró un contenido de taninos superior a 12 mg/mL.
Preparación de la fracción polifenólica
La fracción polifenólica se obtuvo a partir del extracto acuoso de R. mangle, a una concentración de 24 mg/mL, mediante la precipitación con NaCl, en una proporción 1:2,8 (p/v).19 El NaCl se añadió lentamente al extracto, en condiciones de agitación y calentamiento a 70 °C durante 10 min. El sobrenadante se decantó y las muestras se centrifugaron a 3 500 rpm durante 15 min a temperatura ambiente. El precipitado obtenido se disolvió en agua desionizada y se le realizaron tres extracciones con n-butanol 1:2 (v/v) para eliminar las sales contaminantes. La fase butanólica se colectó y se rotoevaporó hasta sequedad para la eliminación del solvente orgánico. El material obtenido se liofilizó y se conservó a -20 °C.
Determinación del contenido de taninos
El contenido de taninos en el extracto acuoso de R. mangle y su fracción polifenólica se determinó como antes se describió.20 Brevemente, este método colorimétrico mide la cantidad de taninos precipitados por una proteína estándar, la albúmina de suero bovino (BSA). La mezcla de reacción contenía: extracto de R. mangle, su fracción polifenólica o ácido tánico como estándar (1 mL) y BSA (1 mg/mL), los cuales inicialmente se mezclan y posteriormente se dejan en reposo durante 15 min a temperatura ambiente. A continuación, se centrifuga a 3 000 g durante 15 min, el sobrenadante se desechó y el pellet se disolvió en 15 mL de laurilsulfato de sodio (1 %) y a 1 mL se le añadieron 3 mL de una solución que contenía laurilsulfato de sodio (1 %) y trietanolamina (7 %) y 1 mL de FeCl3 (0,01 M en HCl 0,1 N) e inmediatamente la mezcla se agitó. Después de 15 min, se midió la absorbancia a 510 nm y la concentración de taninos se calculó usando una curva patrón de ácido tánico.
Ensayo de daño oxidativo a proteínas
Se empleó la BSA como proteína modelo. El daño oxidativo a la BSA se logró por su exposición a radicales hidroxilos generados en el sistema Fenton.21 La mezcla de reacción para la oxidación de la proteína contenía en un volumen final de 1 mL, los siguientes reactivos en las concentraciones finales indicadas: BSA (1 mg/mL) en solución amortiguadora fosfato 50 mM pH 7,4, extracto de R. mangle (4,0; 40,5; 405,2 µg taninos/mL) o su fracción polifenólica (4,1; 41,2; 411,8 µg taninos/mL), EDTA (3,0 mM), FeCl3 (1,0 mM), H2O2 (2,5 mM) y ácido ascórbico (1,0 mM) y se incubó a 25 ºC durante 45 min. Se utilizó como control la solución de BSA, en un volumen igual de solución amortiguadora fosfato 50 mM pH 7.4. A las muestras incubadas se les añadió 1,0 mL de ácido tricloroaético (TCA) 10 % (p/v) para precipitar las proteínas, a continuación se centrifugó a 937 g durante 10 min a 4 °C y se decantó el sobrenadante. El precipitado se lavó con 1,0 mL de TCA 10 % y se removió una vez más el sobrenadante. El precipitado se disolvió en 1,0 mL de solución amortiguadora fosfato 50 mM pH 7,4 y se le determinó el contenido de grupos sulfidrilos,22 para evaluar la magnitud del daño oxidativo a esta proteína. Brevemente, este ensayo consistió en hacer reaccionar las muestras con el ácido 5,5' ditio-bis (2-nitrobenzoico) (reactivo de Ellman's) 4 mg/mL para obtener un cromógeno y después se midió la absorbancia a 412 nm. Se utilizó una curva patrón de glutatión. Los resultados fueron expresados como porcentaje de inhibición de la pérdida de grupos sulfidrilos en la BSA.
Ensayo de daño oxidativo al ADN
El daño oxidativo sobre el ADN fue inducido por el sistema bleomicina-Fe3+.23 La mezcla de reacción contenía en un volumen final de 1 mL, los siguientes reactivos en las concentraciones finales indicadas: ADN de esperma de salmón (1 mg/mL) en solución amortiguadora fosfato 20 mM pH 7,4, extracto de R. mangle (12,6; 50,6; 202,6; 810,3 µg taninos/mL) y su fracción polifenólica (12,7; 51,5; 205,9; 823,6 µg taninos/mL), bleomicina (0,05 mg/mL), FeCl3 (50 µM). Estos reactivos se mezclaron previo a la adición de la solución amortiguadora fosfato 20 mM pH 7,4 conteniendo MgCl2 (5 mM) y ácido ascórbico (0,2 mM). La mezcla de reacción se incubó durante 30 min a 37°C, a continuación se adicionó 0,1 mL de EDTA (0,1 M) para detener la reacción y se tomaron 500 µL para la determinación de los productos reactivos al ácido tiobarbitúrico (PRATB).24 A 500 µL de muestra se le añadieron 1,95 mL de una mezcla que contiene ácido tiobarbitúrico (0,5 %) y ácido tricloroacético (15 %) en ácido clorhídrico (0,25 N) y después se añadieron 50 µL de BHT (1 %). La mezcla de reacción se calentó 15 min a 100 °C, se centrifugó a 1 000 g durante 15 min. La densidad óptica del sobrenadante se midió a 535 nm. La formación de PRATB se cuantificó empleando el coeficiente de extinción del MDA e535= 1,56×105 M-1cm-1. Los resultados se expresaron como porcentaje de inhibición de la degradación del ADN.
Análisis estadístico
Los resultados experimentales son expresados como la media ± EEM (error estándar de la media). Todas las mediciones se realizaron por triplicado. Los datos se analizaron mediante un análisis de regresión lineal para obtener las curvas de regresión que relacionan la concentración de taninos con el efecto de inhibición del daño oxidativo a proteínas y al ADN, ejercido por el extracto de R. mangle y su fracción polifenólica; después se realizó la comparación de estas, con el paquete estadístico Statgraphics versión 4.1 para Windows. Se consideraron valores estadísticamente significativos para p< 0,05.
RESULTADOS
Efecto del extracto de R. mangle y su fracción polifenólica sobre el daño oxidativo a proteínas
En la figura 1 se muestra el análisis de regresión de la inhibición de la pérdida de grupos sulfidrilos en la BSA en función de la concentración de taninos del extracto acuoso de R. mangle y su fracción polifenólica. La incubación del extracto o su fracción polifenólica con la BSA, expuesta a los reactantes del sistema Fenton, resultó en una disminución del daño oxidativo a esta biomolécula, evidenciado en la inhibición de la pérdida de grupos sulfidrilos en la proteína. A la máxima concentración ensayada, el extracto y su fracción polifenólica disminuyeron la oxidación de los grupos SH en 87,3 y 89,1 %, respectivamente. Este efecto fue dependiente de la concentración de taninos con valores de índice de determinación (r2) de 0,939 y 0,971 para el extracto y la fracción respectivamente. Los valores de la concentración inhibitoria media (CI50) del extracto y la fracción fueron 0,67 y 0,36 µg taninos/mL, respectivamente. La comparación de las líneas de regresión reveló que el extracto y su fracción resultaron igualmente eficaces en proteger a la BSA de la oxidación por los radicales hidroxilos.
Efecto del extracto de R. mangle y su fracción polifenólica sobre el daño oxidativo al ADN
En la figura 2 se muestra el análisis de regresión de la inhibición del daño oxidativo al ADN en función de la concentración de taninos del extracto acuoso de R. mangle y su fracción polifenólica. La incubación del extracto o su fracción polifenólica con el ADN, expuesto al sistema bleomicina-Fe3+, resultó en una inhibición del daño oxidativo a esta biomolécula, evidenciado por la disminución de la formación de PRATB. A la máxima concentración ensayada, el extracto y su fracción polifenólica, disminuyeron la oxidación del ADN en 98,4 y 91,9 %, respectivamente. Este efecto fue dependiente de la concentración de taninos con valores de índice de determinación (r2) de 0,925 y 0,957 para el extracto y la fracción respectivamente. Los valores de la CI50 del extracto y la fracción fueron 0,084 y 0,59 µg taninos/mL, respectivamente, que demuestra la superioridad del extracto en este efecto. La comparación de las rectas de regresión reveló diferencias significativas (p< 0,05) entre estas, lo cual indica que en el intervalo de concentraciones estudiadas el extracto fue más eficaz en proteger al ADN de la degradación oxidativa que su fracción.
DISCUSIÓN
Los resultados del presente trabajo indican que el extracto acuoso de R. mangle y su fracción polifenólica disminuyeron el daño oxidativo de la BSA ocasionado por los radicales hidroxilo, medido por la inhibición de la pérdida de grupos sulfidrilos en la proteína.
La literatura refiere que los aminoácidos también son sensibles al daño oxidativo.25 En este sentido, un estudio revela que todos los aminoácidos de la BSA fueron modificados por la exposición de esta proteína a los radicales hidroxilo,26 y resultan más vulnerables los residuos de triptófano, histidina y cisteína, lo cual está en concordancia con las constantes de velocidad de reacción de estos aminoácidos libres con los radicales hidroxilo.27 Otros trabajos también informan acerca de la pérdida de grupos sulfidrilo en la BSA expuesta a radicales hidroxilo, generados por radiolisis gamma, xantina oxidasa + EDTA-Fe3+ o Fe2+ + H2O2.28-30
Los valores de inhibición de la pérdida de grupos sulfidrilos obtenidos para la fracción, aportan más de 95 % al efecto antioxidante del extracto en el intervalo de concentraciones estudiadas, lo cual sugiere que los compuestos polifenólicos presentes en la fracción son los responsables de la disminución del daño oxidativo a la BSA. En concordancia con estos resultados, otros autores han encontrado efectos protectores sobre esta y otras proteínas con el empleo de otros extractos de plantas que contienen polifenoles o evaluando los compuestos polifenólicos puros. Así, se ha probado que el extracto acuoso de la corteza de la planta Mangifera indica L. atenuó la pérdida de grupos sulfidrilos tanto en la BSA expuesta al mismo sistema de estudio del presente trabajo,31 como en un homogenado de hígado de ratón tratado con TPA; efectos similares fueron obtenidos al emplear la mangiferina, principal polifenol presente en este extracto.32 En otro trabajo se indica que las formas radicalarias de los aminoácidos aromáticos triptófano y tirosina, los cuales están entre los aminoácidos más oxidables en las proteínas, son reparadas por los compuestos polifenólicos quercetina, catequina y rutina mediante reacciones de transferencia de electrones para generar radicales de compuestos polifenólicos semioxidados.33
Los posibles mecanismos que pueden explicar el efecto protector de los polifenoles sobre la pérdida de grupos sulfidrilos en la BSA ocasionada por los radicales hidroxilo, son las propiedades secuestradoras de esta ERO y la actividad quelante de iones de hierro previamente demostrada para los compuestos polifenólicos que conforman la fracción mayoritaria del extracto acuoso de R. mangle.16 En presencia de compuestos polifenólicos, la disminución de la pérdida de grupos SH en la BSA está dada porque la capacidad de donar átomos de hidrógeno descrita para los polifenoles hace más probable la reacción ·OH-polifenoles que ·OH-grupos SH.
En el presente trabajo también se demostró que tanto el extracto como su fracción disminuyeron el daño oxidativo al ADN inducido por el complejo bleomicina-Fe3+. Previamente se informó que el ADN de células tratadas con bleomicina resulta degradado.34 Se ha comprobado que la bleomicina provoca la ruptura de las cadenas dobles y simples en preparaciones purificadas de ADN 35 y que la escisión de cadena ocurre preferencialmente en la secuencia de bases guanina-pirimidina, en la dirección 5' ® 3'.36 Un estudio del mecanismo responsable de la escisión de cadena del ADN inducida por bleomicina concluyó que los productos de la reacción primaria resultante incluyen: oligonucleótidos y bases de propenal, estas últimas con una estructura común: base-CH=CH-CHO,37 las cuales forman cromóforos con el ácido tiobarbitúrico y pueden ser detectadas por el ensayo de PRATB.38 Estos autores plantearon que el grupo propenal es derivado de los primeros tres carbonos de la desoxirribosa, mientras los carbonos restantes C-4' y C-5' de este azúcar permanecen como un residuo de ácido glicólico esterificado a través del grupo hidroxilo al fosfato 3' terminal en el oligonucleótido residual. Estos investigadores sugieren que esas observaciones son consistentes con un mecanismo de ruptura de cadena del ADN, que involucra la captura de un átomo de hidrógeno del C-4' de la desoxirribosa por radicales libres, lo cual provoca la escisión del enlace (C-3')-(C-4'), otros trabajos coinciden en considerar estos eventos como el posible mecanismo que explica la escisión de cadena en esta biomolécula inducida por radicales libres;36 los azúcares del ADN resultan el principal blanco de daño oxidativo por el sistema bleomicina-Fe3+. Varios investigadores han concordado en sugerir que las especies radicalarias requeridas para este mecanismo pueden ser generadas a partir de un complejo bleomicina-Fe (II)-O2.36,38 Se ha propuesto un modelo acerca de este complejo, su principal rasgo consiste en la coordinación del oxígeno con el metal y la inserción preferencial de los anillos de bitiazol de la bleomicina entre los pares de base del ADN, en los cuales una cadena contiene una secuencia de bases GT o GC. La unión al ADN puede ser estabilizada a través de interacciones electrostáticas que se establecen entre las cadenas laterales terminales cargadas positivas de la bleomicina y los grupos fosfato cargados negativos del ADN. En este modelo el O2 está estratégicamente posicionado, de tal forma que la reducción de una especie radicalaria conlleve a que un átomo de hidrógeno unido al C-4' del anillo de la desoxirribosa sea capturado de manera selectiva, guiando al clivaje oxidativo del enlace (C-3')-(C-4') en este azúcar. Si el complejo bleomicina-Fe (III)-O2 es reducido in situ, los ·OH pueden ser generados en la vecindad inmediata de los grupos adyacentes sobre el ADN y provocar su ruptura.37 Los parámetros cinéticos de la reacción de escisión del anillo de la desoxirribosa han sido reportados y la rápida ruptura observada en este azúcar, sugiere que los radicales hidroxilo pueden ser la especie directamente involucrada en la fragmentación del ADN inducida por bleomicina.38
Los elevados porcentajes que aporta la fracción a la actividad antioxidante del extracto en este ensayo indican que los compuestos polifenólicos presentes en esta son de vital importancia para obtener los efectos mostrados por el extracto. Existen reportes en la literatura acerca del efecto protector que ejercen los polifenoles puros o aquellos integrados a extractos vegetales, sobre el daño oxidativo al ADN ocasionado por la bleomicina. En correspondencia con esta idea, el extracto acuoso de cortezas de Mangifera indica L. (Vimang®) protegió del daño oxidativo al ADN inducido por bleomicina- Fe3+.39 Un estudio reportó que el extracto de semillas de la uva, en el que se destaca la presencia de compuestos polifenólicos como: catequina, epicatequina, trans-resveratrol y ácido gálico, disminuyó la actividad mutagénica de la bleomicina en la cepa TA102 de Salmonella typhimurium.40 Empleando esta misma cepa como modelo de estudio, otros investigadores demostraron que el mecanismo que explica la actividad antimutagénica de los polifenoles es su capacidad secuestradora de radicales libres.41 Coincidiendo con estos hallazgos, otros científicos encontraron una buena correlación entre la actividad antioxidante de la catequina, la epicatequina y un extracto de té verde, determinada por sus propiedades secuestradoras de radicales DPPH, O2·- y H2O2, y su acción antimutagénica.42 En otro estudio se indica que el extracto de uvas inhibió la ruptura de cadenas del ADN inducida por mitomicina-c,43 lo cual sugiere que este extracto puede inhibir el daño al ADN ocasionado por ERO. Otro reporte informa que cuando células del bazo de ratones son preincubadas con compuestos polifenólicos aislados de la semilla de la uva, como: procianidina B4, catequina y ácido gálico; resultaron menos susceptibles al daño al ADN inducido por el peróxido de hidrógeno según los resultados del ensayo cometa.44
Estos resultados permiten concluir que el extracto acuoso de R. mangle mostró un efecto protector a las principales biomoléculas del daño oxidativo, evidenciado por la inhibición de la pérdida de grupos sulfidrilos en la BSA y la disminución de la degradación del ADN. A su vez se demostró que los compuestos polifenólicos presentes en el extracto fueron los principales responsables de los efectos antioxidantes observados en este estudio.
REFERENCIAS BIBLIOGRÁFICAS
1. Kohen R, Nyska A. Oxidation of biological systems: Oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. Toxicologic Pathology. 2002;30(6):620-50.
2. McCord JM. The Evolution of free radicals and oxidative stress. Am J Med. 2000;108:652-59.
3. Halliwell B. Antioxidants in human health and disease. Annual Review Nutrition. 1996;16(1):33-50.
4. Sánchez LM, Melchor G, Álvarez S, Bulnes C. Caracterización química y toxicológica de una formulación cicatrizante de Rhizophora mangle L. Rev Salud Anim. 1998;20(2):69-72.
5. Roig JT. Plantas medicinales, aromáticas y venenosas de Cuba. 2da ed. La Habana Editorial Científico-Técnica; 1988. p. 606-7.
6. Rojas N, Coto O. Propiedades antimicrobianas de extractos de Rhizophora mangle L. Rev Cubana Med Tropical. 1978;30(3):181-7.
7. Cáceres A, López B, Juárez X, del Aguila J, García S. Plants used in Guatemala for the treatment of dermatophytic infections 2: Evaluation of antifungal activity of seven American plants. J Ethnopharmacol. 1993;40:207-13.
8. Armenteros M. Evaluación de un desinfectante mamario post-ordeño de origen natural [Tesis doctoral]. La Habana: Centro Nacional de Sanidad Agropecuaria, Universidad Agraria de La Habana; 1998.
9. Bulnes C, Fernández O, Navarro D, Marrero E, Rueda D, Figueroa O, et al. Healing effect of a red mangrove extract in open aseptic wounds in rat. Rev Salud Anim. 2001;23(2):102-8.
10. Fernández O, Capdevila JZ, Dalla G, Melchor G. Efficacy of Rhizophora mangle aqueous bark extract in the healing of open surgical wounds. Fitoterapia. 2002;73:564-8.
11. Montes de Oca N, Reverón Y, González R. Evaluation of antimicrobial activity of different extracts of Rhizophora mangle L using five methods. Rev Salud Anim. 2001;23:1-11.
12. Melchor G, Armenteros M, Fernández O, Linares E, Fragas I. Antibacterial activity of Rhizophora mangle bark. Fitoterapia. 2001;72:689-91.
13. Agüero F. Evaluación de un producto natural, a base de Rhizophora mangle L. en la terapia de la endometritis bovina [Tesis doctoral]. La Habana: Centro Nacional de Sanidad Agropecuaria, Universidad Agraria de La Habana. 2004.
14. Sánchez LM, Rueda D, Gómez BC. Gastric antiulcer effect of Rhizophora mangle L. J Ethnopharmacol. 2001;77:1-3.
15. Marrero E, Sánchez J, de Armas E, Escobar A, Melchor G, Abad MJ, et al. COX-2 and sPLA2 inhibitory activity of aqueous extract polyphenols of Rhizophora mangle (red mangrove). Fitoterapia. 2006;77:313-5.
16. Sánchez J, Melchor G, Martínez G, Escobar A, Faure R. Antioxidant activity of Rhizophora mangle bark. Fitoterapia. 2006;77:141-3.
17. Sánchez J, Melchor G, Martínez G, Sánchez LM, Faure R, Vinardel P. Protective effect of Rhizophora mangle bark on lipid peroxidation and erythrocyte hemolysis. Phcog Mag. 2005;1(3):101-4.
18. Sánchez J, Faure R, Martínez G, Vega E, Fernández O. Propiedades antioxidantes de Rhizophora mangle L. y su relación con el proceso de curación de heridas en ratas. Rev Salud Anim. 2009;31(3):170-5.
19. Matsuo T, McCord Itoo SA. Simple and rapid purification method of condensed tannins from several young fruits. Agric Biol Chem. 1980;45:1885-7.
20. Hagermann AE, Butler LG. Protein precipitation method for the determination of tannins. J Agric Food Chem. 1978;26:809-12.
21. Kingu CS, Wei R. Alcohol deshydrogenase catalyzed reduction of protein carbonyl derivatives. J Biochem Biophys Meth. 1997;34:61-8.
22. Sedlak J, Lindsay RH. Estimation of total protein-bound and nonprotein sulfhydryl groups in tissue with Ellman's reagent. Anal Biochem. 1968;25:192-205.
23. Aruoma OI. Use of DNA damage as a measure of pro oxidant actions of antioxidant food additives and nutrient components. In: Halliwell B Aruoma OI, editors. DNA and free radical. Ellis Horwood: London; 1993. p. 315 27.
24. Buege JA, Aust SD. Microsomal lipid peroxidation. Methods Enzymol. 1978;30:301-10.
25. Davies KJA. Protein damage and degradation by oxygen radicals. I. General aspects. Modification of aminoacids. J Biol Chem. 1987;262(20):9895-901.
26. Davies KJ, Delsignore ME, Lin SW. Protein damage and degradation by oxygen radicals. II. Modification of amino acids. J Biol Chem. 1987;262(20):9902-7.
27. Dorfman LM, Adams GE. Reactivity of the hydroxyl radical in aqueous solution. National Standards Reference Series 46, Washington, D.C.: National Bureau of Standards; 1973.
28. Di Simplicio P, Cheeseman KH, Slater TF. The reactivity of the SH group of bovine serum albumin with free radicals. Free Radic Res Commun. 1991;14(4):253-62.
29. Miura T, Muraoka S, Ogiso T. Oxidative damage to bovine serum albumin induced by hydroxyl radical generating systems of xanthine oxidase + EDTA-Fe3+ and ascorbate + EDTA-Fe3+. Chem Biol Interact. 1992;85(2-3):243-54.
30. Davies MJ, Gilbert BC, Haywood RM. Radical-induced damage to bovine serum albumin: role of the cysteine residue. Free Radic Res Commun. 1993;18(6):353-67.
31. Martínez G, Giuliani A, León OS, Pérez G, Nuñez Sellés AJ. Effect of Mangifera indica L. Extract (QF808) on Protein and Hepatic Microsome Peroxidation. Phytother Res. 2001;15:581-5.
32. Martínez G, Giuliani A, Nuñez AJ, Pérez G, León OS. Protective effects of mangifera indica L. Extract, mangiferin and selected antioxidants against TPA-induced biomolecules oxidation and peritoneal macrophage activation in mice. Pharmacological Research. 2000;42(6):565-73.
33. Filipe P, Morliere P, Patterson LK, Hug GL, Maziere JC, Maziere C, et al. Mechanisms of flavonoid repair reactions with amino acid radicals in models of biological systems: a pulse radiolysis study in micelles and human serum albumin. Biochimica et Biophysica Acta. 2002;1572:150-62.
34. Moore CW. Degradation of DNA and Structure-Activity Relationship between Bleomycins A2 and B2 in the Absence of DNA Repair. Biochemistry. 1990;29:1342-7.
35. Solomon LR, Beerelli RD, Moseley PL. Bleomycin-iron can degrade DNA in the presence of excess ethylenediaminetetraacetic acid in vitro. Biochemistry. 1989;28(26):9932-7.
36. Takeshita M, Grollman AP, Ohtsubo E, Ohtsubo E. Interaction of Bleomycin with DNA. PNAS. 1978;75(12):5983-7.
37. Giloni L, Takeshita M, Johnson F, Iden C, Grohant AP. Bleomycin-induced strand-scission of DNA. Mechanism of deoxyribose cleavage. J Biological Chemistry. 1981;256(16):8608-15.
38. Povirk LF. Catalytic release of deoxyribonucleic acid bases by oxidation and reduction of an iron-bleomycin complex. Biochemistry. 1979;18:3989-95.
39. Martínez G, Delgado R, Pérez G, Garrido G, Núñez AJ. Evaluation of the in vitro Antioxidant Activity of Mangifera indica L. Extract (Vimang). Phytother Res. 2000;14:424-7.
40. Stagos D, Kazantzoglou G, Theofanidou D, Kakalopoulou G, Magiatis P, Mitaku S, et al. Activity of grape extracts from Greek varieties of Vitis vinifera against mutagenicity induced by bleomycin and hydrogen peroxide in Salmonella typhimurium strain TA102. Mutat Res. 2006;609(2):165-75.
41. Edenharder R, Grunhage D. Free radical scavenging abilities of flavonoids as mechanism of protection against mutagenicity induced by tert-butyl hydroperoxide or cumene hydroperoxide in Salmonella typhimurium TA102. Mutat Res. 2003;540(1):1-18.
42. Geetha T, Garg A, Chopra K, Kaur I. Delineation of antimutagenic activity of catechin, epicatechin and green tea extract. Mutat Res. 2004;556(1-2):65-74.
43. Stagos D, Kazantzoglou G, Magiatis P, Mitaku S, Anagnostopoulos K, Kouretas D. Effects of plant phenolics and grape extracts from Greek varieties of Vitis vinifera on Mitomycin C and topoisomerase I-induced nicking of DNA. Int J Mol Med. 2005;15(6):1013-22.
44. Fan P, Lou H. Effects of polyphenols from grape seeds on oxidative damage to cellular DNA. Mol Cell Biochem. 2004;267(1-2):67-74.
Recibido: 1 de agosto de 2010.
Aprobado: 10 de octubre de 2010.
Janet Sánchez Calero. Departamento de Química-Farmacología-Toxicología, Centro Nacional de Sanidad Agropecuaria. San José de Las Lajas, La Habana, Cuba. Correo electrónico: jsanchez@censa.edu.cu


{kind=link}
{kind=link}