










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkFitosanidad
versión On-line ISSN 1818-1686
Fitosanidad v.13 n.2 Ciudad de la Habana abr.-jun. 2009
Obtención de aislados de Bacillus thuringiensis Berliner autóctonos de Cuba
Determination of Bacillus thuringiensis BerlinerIsolates Autochthonous in Cuba
Bertha Carreras Solís
Instituto de Investigaciones de Sanidad Vegetal. Calle 110 no. 514 e/ 5.a B y 5.a F, Playa, Ciudad de La Habana, CP 11600, carreras@inisav.cu
RESUMEN
La presencia de aislados nativos de Bacillus thuringiensis se determinó mediante el análisis de 87 muestras colectadas de las diferentes regiones de Cuba. El proceso de pasteurización se utilizó para la selección de colonias esporuladas crecidas en medio nutriente. Las colonias se seleccionaron por su aspecto y morfología mediante observación al microscopio estereoscópico. Se observó la típica forma bacilar, posición y morfología de la endospora por microscopía óptica, características del grupo cereus del género Bacillus. La especie B. thuringiensis se reconoció básicamente debido a la presencia del cristal paraesporal. El índice B. thuringiensisobtenido se determinó por la relación entre el número de colonias de B. thuringiensis seleccionadas y el número total de colonias examinadas en la muestra. Colonias de 1-1,5 cm de diámetro, planas, opacas, de color blanco-grisáceo, de consistencia ligeramente costrosa, bordes irregulares, células vegetativas en forma de bacilos y esporas elipsoidales en posición terminal dentro de la célula vegetativa permitieron la selección de 609 colonias esporuladas. De estas, 341 colonias se clasificaron como B. thuringiensis sobre la base de la producción del cristal paraesporal. Los aislados de B. thuringiensis mostraron tres morfologías del cristal: bipiramidales predominantemente amorfos y dos aislados produjeron cuerpos paraesporales pequeños con aparienciade gorro sobre la espora. Los índices de B. thuringiensis variaron de 0,37 a 0,74, y el índice medio en el total de colonias observadas fue de 0,56.
Palabras claves: Bacillus thuringiensis, aislamiento, cristal paraesporal
ABSTRACT
The presence of native isolates of Bacillus thuringiensis was determinedby the analysis of 87 samples collected from different regions of Cuba. The pasteurization was utilized for the selection of sporulating colonies growed in nutrient medium. The colonies were selected because of their aspect and morphology by stereoscopic observation. It was observed the typical bacilar shape, position and morphology of the endospore by light microscopy, characteristics of cereus group, Bacillus genus. The bacterium was recognized basically due to the presence of a paraesporal crystal. The B. thuringiensis index obtainedwas determined by the relation between the numbers of B. thuringiensis colonies found with respect to the total number of colonies examined in the sample. 1-1,5 cm diameter colonies, plane, opaque, white-gray, lightly scabby consistence, irregular border, vegetative cells the typical bacilar shape and elipsoidal terminal endospores inside the vegetative cell allowed the selection of 609 sporulating colonies. From these, 341 colonies were classified as B. thuringiensis based on the production of parasporal crystal. B. thuringiensis isolates showed three crystal morphologies: bipyramidal mainly, amorphous, two isolates produced small parasporal bodies which appeared as a «cap» on the spore. The B. thuringiensis index varied from 0.37 to 0.74 and the B. thuringiensis index total was 0.56.
Key words: Bacillus thuringiensis, isolates, parasporal crystal
INTRODUCCIÓN
Los productos que se obtienen a partir de la bacteria Bacillus thuringiensis Berliner dominan el mercado de los bioplaguicidas, y el uso constante y creciente de sus productos por cuatro décadas se debe fundamentalmente a su alta especificidad, así como a su inocuidad para insectos benéficos, plantas y mamíferos, incluidos los humanos [Ghelardi et al., 2002; Támez et al., 2007]. El componente tóxico principal de B. thuringiensis es la proteína cristal (Cry) o ä-endotoxina, la cual tiene un espectro de actividad biológica definido, usualmente restringido a pocas especies dentro de un grupo particular de organismos.
La búsqueda de nuevas cepas de esta bacteria en los hábitats naturales, donde se encuentra con relativa frecuencia, se ha convertido en una práctica internacional que ha permitido aumentar las posibilidades de uso a partir de nuevas potencialidades de control sobre organismos que constituyen plagas. Mediante la prospección y aislamiento en diferentes regiones geográficas se ha logrado el enriquecimiento de los ceparios que conservan esta especie con un elevado potencial para el control de insectos y otros organismos plaga [Martin y Travers, 1989; Ruiz de Escudero et al., 2004; Armengol et al., 2007; Rosas et al., 2008].
El Instituto de Investigaciones de Sanidad Vegetal (Inisav) ha tenido la responsabilidad de conservar y suministrar las cepas para la producción de bioplaguicidas en Cuba, y cuenta con un grupo de cepas de B. thuringiensis cuyo origen no está totalmente definido debido a que no se ha realizado un estudio orientado a determinar la presencia de B. thuringiensis en diferentes hábitats de las regiones del país y que permita conocer el verdadero potencial de esta especie. De manera que el objetivo principal de la investigación fue la obtención de aislados de Bacillus thuringiensis, Berliner, autóctonos de Cuba.
MATERIALES Y MÉTODOS
Se analizaron 87 muestras colectadas de las diferentes regiones de Cuba (Tabla 1). Las muestras de suelo se recogieron en prospecciones de campo realizadas en las diferentes provincias del país de abril a junio y de septiembre a noviembre del 2004 y del 2005. Se tomaron muestras tanto de suelos de hábitats naturales (zonas boscosas) como de suelos cultivados, donde los biopreparados de B. thuringiensis nunca se han aplicado. También se hicieron aislamientos de rizosfera, semillas, raíces y hojas de plantas. En cada punto de muestreo, con un área estimada de 9 m2, se tomaron varias submuestras, en una superficie de 0,10 m2 cada una.

De cada una de las muestras procesadas se tomó 1 g que se mezcló con 10 mL de agua destilada estéril en tubos de ensayo, y se sometió a agitación vigorosa. Los tubos se incubaron en un baño de María a 65°C durante 30 min con el objetivo de seleccionar las esporas presentes en la mezcla. Luego se dejó en hielo por igual tiempo y se procedió a realizar diluciones decimales. Cien microlitros de las diluciones se depositaron en placas con medio nutriente, se esparcieron con espátula de Drygalski y se incubaron a 28-30°C durante 48-72 h hasta visualizar colonias individuales. Se examinaron las colonias crecidas y se seleccionaron por su aspecto y morfología mediante observación al microscopio estereoscópico. Se determinó la forma bacilar, distensión del esporangio, localización y morfología de la espora mediante observación de la tinción de Wirtz al microscopio óptico 1000 X [Valenzuela, 2003], características descritas para el grupo cereus del género Bacillus [Boone et al., 2001; Porcar y Juárez, 2004].
Se utilizó como criterio de selección de B. thuringiensis respecto a B. cereus la presencia de un cuerpo paraesporal (cristal) [Boone et al., 2001] mediante observación de la tinción simple con violeta cristal al microscopio óptico 1000 X.
Se determinó el índice de B. thuringiensis obtenido por la relación del número de colonias a las que estuvo asociado el cuerpo paraesporal y el número de colonias examinadas con las características culturales de Bacillus:

Para la conservación de los aislados se preparó una suspensión de células muy densa de estos y se añadieron 100 ìL a una placa de Petri estéril. El inóculo se distribuyó en la superficie de la placa y se le colocaron unos discos de papel de filtro estériles. Se dejó secar en flujo laminar. Los discos de papel de filtro secos se guardaron en tubos eppendorf estériles y se conservaron a 4°C hasta su uso.
RESULTADOS Y DISCUSIÓN
Las colonias de 1-1,5 cm de diámetro, de color blancogrisáceo, planas, opacas, de consistencia ligeramente costrosa y que mostraron indistintamente dos tipos de bordes irregulares, lobulados o ramificados, ambos incluidos en las características descritas para el grupo cereus del género Bacillus (Fig. 1), se consideraron para seleccionar 609 colonias como posibles B. thuringiensis, criterio utilizado por Porcar y Juárez (2004).

La posición y la morfología de la endospora en el interior de la bacteria constituyen un carácter taxonómico útil para diferenciar especies dentro de un mismo género.Bacillus thuringiensis tiene localizada la endospora elipsoidal en posición terminal dentro de la célula vegetativa. La Fig. 2 muestra esta posición y morfología en una de las colonias seleccionadas. Se observa la endospora teñida de color verde y las células vegetativas de color rojo.
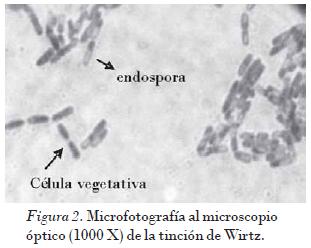
La presencia de las células vegetativas en forma de bacilos de 1,0-1,2 ìm de ancho por 3-5 ìm de largo (Fig. 2), así como la formación de esporas ovales a las cuales se encontró asociado un cristal paraesporal de forma bipiramidal, esférico o amorfo teñido de color azul (Fig. 3), permitió confirmar la presencia de B. thuringiensis. La aparición del cristal es una característica distintiva de B. thuringiensis, la cual permite separarla de otras especies cercanas filogenéticamente como Bacillus cereus [Boone et al., 2001; Porcar y Juárez, 2004].
En los aislados seleccionados predominó la morfología bipiramidal de las inclusiones cristalinas, aunque también se observó la morfología esférica/amorfa y en muy pocos aislados un cristal muy pequeño adherido a la espora (Fig. 3). De manera que, sobre el criterio de la capacidad de producción del cristal paraesporal, de 609 colonias analizadas 341 se identificaron como B. thuringiensis.

Mediante la utilización de esta técnica de identificación muchos autores han realizado trabajos de aislamiento de B. thuringiensis y se han establecido grandes colecciones de esta especie, como los de Ruiz de Escudero et al. (2004), quienes seleccionaron 1230 colonias con las características morfológicas de B. thuringiensis, y de estas 684 presentaron cristal paraesporal.
De manera similar Arrieta et al. (2004), de 641 muestras procesadas lograron aislar 300 colonias con morfología similar a B. thuringiensis, y de ellas un total de 202 mostraron cristales. Se presentó una gran diversidad de cristales por cepa, y entre las cepas donde las morfologías más frecuentes fueron la redondeada e irregular. Los resultados del presente trabajo coincidieron más con los de Ruiz de Escudero et al. (2004) y Rosas et al. (2008), en que hubo un predominio de la morfología bipiramidal. Los cristales bipiramidales se han asociado a B. thuringiensis desde su descubrimiento en 1901 por Shigetane Ishiwata [Aizawa, 2001].
La eficiencia del aislamiento de cepas de B. thuringiensis con cristales del 56% con respecto a los bacilos esporulados acristalíferos resultó similar a lo reportado por otros autores en España y Costa Rica [Bel et al., 1997; Arrieta et al., 2004], lo cual confirma la validez del método empleado.
Con relación al total de muestras analizadas, B. thuringiensis se encontró en el 74,7%, y con las muestras de suelo estuvo en el 82,5%. Se dispone de información en los que el porcentaje de muestras con presencia deB. thuringiensis oscila entre el 20 y el 30% [Iriarte et al., 1998], o hasta el 92% [Bel et al., 1997]. La mayor o menor riqueza de muestras con B. thuringiensis puede depender del número de colonias identificadas en cada muestra. Para tener en cuenta esto se utilizó el índice B. thuringiensis que se considera como una medida del éxito en el aislamiento de esta especie.
El índice B. thuringiensis obtenido para suelo resultó de 0,6 (Tabla 2). En muestras de suelo este índice puede tomar valores muy bajos de 0,0060-0,0092 [Quesada etal., 2004; Rosas et al., 2008], valores medios de 0,5 [Ruiz de Escudero et al., 2004], o valores altos de 0,85 cuando se usan medios selectivos (medio con acetato) [Martin y Travers, 1989].

El valor de 0,6 para las muestras de suelo analizadas indica una buena incidencia de la especie en este hábitat. Esta eficiencia puede atribuirse a que cuando las muestras se colectaron, se rechazó la superficie, y el material de la muestra pertenece a la capa del suelo que está por debajo de 5-10 cm de la superficie, a niveles en que los daños causados por la luz UV no son posibles y la temperatura es más estable [Bel et al., 1997]. De manera general los resultados sugieren que los suelos de Cuba son una importante fuente de B. thuringiensis, ya que el índice obtenido fue comparable con los valores informados previamente por otros autores en muestreos de suelos hechos en distintas regiones de los cinco continentes [Martin y Travers, 1989; Ruiz de Escudero et al., 2004; Chatterjee et al., 2007].
En el caso de rizosfera, hoja, semilla y raíz, mostraron índices de incidencia de B. thuringiensis más bajos (Tabla 2), aunque se debe considerar que el número de muestras analizadas fue mucho menor.
Los más bajos índices B. thuringiensis en rizosfera respecto a suelo puede atribuirse a que, como plantean Medrano et al. (2000), B. thuringiensis no se considera como parte de su flora nativa, pues está sujeto a la competencia y depredación de los microorganismos autóctonos de este ambiente, y a su incapacidad para competir por los exudados de las raíces.
En las hojas de pinos se señala que las esporas persisten más de siete meses, pero la experiencia con la aspersión de las esporas y cristales en las hojas de plantas no forestales muestra una limitada viabilidad de esas esporas, principalmente por la acción germicida de la luz visible y UV. De manera general, B. thuringiensis se considera un pobre colonizador de la superficie de las hojas [Maduell et al., 2008]. Esto pudiera dar explicación al índice de 0,39 encontrado en las muestras de hojas analizadas, lo que está en correspondencia con Maduell et al. (2002), quienes informaron índices de 0,2 en especies del género Piper (Piperaceae).
Al analizar la presencia de B. thuringiensis en las diferentes regiones de Cuba se encontraron índices más elevados en las regiones occidental y oriental (Tabla 3).
Aunque en el trabajo no se realiza un análisis de la presencia de B. thuringiensis en los suelos, pues no se recogen sus características y se analizan otras fuentes además de suelo, se observa una correspondencia con los resultados de Ruiz de Escudero et al. (2004). De acuerdo con lo planteado por estos autores, el lugar de procedencia de las muestras, el tipo de suelo, el cultivo implantado y el resto de las propiedades físico-químicas del suelo no mostraron diferencias notables en la riqueza en B. thuringiensis. Los índices variaron desde 0,7 a 0,9, y el índice medio de B. thuringiensis en el total de colonias observadas fue 0,55; sin embargo, Maduell et al. (2005) observaron que el pH del suelo fue una de las variables más importantes en la presencia de B. thuringiensis. Él no mostró un efecto por sí solo. La combinación del pH con otras variables, como la disponibilidad de nutrientes (Ca, Mg, K), el contenido de arcillas y la textura del suelo, tuvieron un efecto sobre la biología de B. thuringiensis.
De manera general, y de acuerdo con la clasificación genética de los suelos de Cuba [Hernández y Ascanio, 2005], estos son mayormente pardo sialíticos en las regiones central y oriental, y ferralíticos en la occidental. En estos suelos existe un predominio de minerales, y el contenido de hierro libre no sobrepasa el 3%; no presentan además pH mayor que 7, condiciones que favorecen la presencia de las esporas de B. thuringiensis [Maduell et al., 2005]. Se considera que la temperatura sí pudiera ejercer una influencia sobre la presencia de esta especie bacteriana por la acción germicida de la luz visible y UV sobre B. thuringiensis. En Cuba las diferencias no son tan marcadas entre las diferentes regiones, por lo que los índices de B. thuringiensis se consideran aceptables.

La diferencia en la frecuencia de aislamiento entre los hábitats se asume que se debe a la sensibilidad de las esporas a la radiación solar y la desecación, hecho asociado con los plásmidos que codifican para la síntesis de la ä-endotoxina, los que causan cambios en la composición química de la cubierta de las esporas y que reducen su tolerancia a estos factores físicos del ambiente [Sánchez y Peña, 2000].
Independientemente de la mayor o menor riqueza encontrada, actualmente se admite que B. thuringiensis es una bacteria distribuida por todo el mundo y se halla prácticamente en todos los hábitats muestreados. Estos resultados claramente revelan la presencia de B. thuringiensis en todo el territorio, y por tanto permiten confirmar que Cuba no es una excepción en este sentido.
CONCLUSIONES
• Se obtuvieron 341 aislados de Bacillus thuringiensis de las diferentes regiones y hábitats de Cuba.
REFERENCIAS
Aizawa, K.: «Shigetane Ishiwata: His Discovery of Sottokin (Bacillus thuringiensis) in 1901 and Subsequent Investigations in Japan», Proceedings of a Centennial Symposium Commemorating Ishiwata's Discovery of Bacillus thuringiensis, Kurume, Japón, 2001, pp. 1-14.
Armengol, G.; M. C. Escobar; M. E. Maldonado; S. Orduz: «Diversity of Colombian Strains of Bacillus thuringiensis with Insecticidal Activity Against Dipteran and Lepidopteran Insects», J. Appl. Microbiol. 102(1):77-88, EE.UU., 2007.
Arrieta, G.; A. Hernández; A. M. Espinoza: «Diversity of Bacillus thuringiensis Strains Isolated from Coffee Plantations Infested with the Coffee Berry Borer Hypothenemus hampei», Rev. Biol. Trop. 52(3):757-764, Costa Rica, 2004.
Bel, Y.; F. Granero; T. Alberola; M. J. Martínez; J. Ferré: «Distribution, Frequency and Diversity of Bacillus thuringiensis in Olive Tree Environments in Spain», System Appl. Microbiol. 20:652-658, EE.UU., 1997.
Boone, D. R.; G. M. Garrity; R. W. Castenholz: Bergy's Manual of Systematic Bacteriology, vol. 1, 2nd ed., Spring, Spring-Verlag, Nueva York, 2001.
Chatterjee, S. N.; T. Bhattacharya; T. K. Dangar; G. Chandra: «Ecology and Diversity of Bacillus thuringiensis in Soil Environment», African Journal of Biotechnology 6(13):1587-1591, 2007.
Ghelardi, E.; F. Celandroni; S. Salvetti; D. J. Beecher; M. Gominet; D. Lereclus; A. C. L. Wong; S. Senesi: «Requirement of flh A for Swarming Differentiation, Flagellin Export, and Secretion of Virulence Associated Proteins in Bacillus thuringiensis», J. Bacteriol. 184:6424-6433, EE.UU., 2002.
Hernández, J. A.; G. M. O. Ascanio: «The Development of Soil Classification in Cuba», Eurasian Soil Science 38 (12):1342-1346, 2005.
Iriarte, J.; Y. Bel; M. D. Ferrandis; R. Andrew; J. Murillo; J. Ferré; P. Caballero: «Environmental Distribution and Diversity of Bacillus thuringiensis in Spain», System. Appl. Microbiol. 21:97-106, EE.UU., 1998.
Maduell, P.; R. Callejas; K. R. Cabrera; G. Armengol; S. Orduz: «Distribution and Characterization of Bacillus thuringiensis on the Phylloplane of Species of Piper (Piperaceae) in Three Altitudinal Levels», Microb. Ecol. 44(2):144-53, EE.UU., 2002.
Maduell, P.; A. Armengol; S. Orduz: «Can Soil Phisicochemical Characteristics Affect the Presence of Bacillus thuringiensis Spores?», Microb. Ecol. 27(82):35-42, EE.UU., 2005.
Maduell, P.; G. Armengol; M. Llagostera; S. Orduz; S. Lindow: «Bacillus thuringiensis Is a Poor Colonist of Leaf Surfaces», Microb. Ecol. 55:212-219, EE.UU., 2008.
Martin, P. A. W.; R. S. Travers: «Worldwide Abundance and Distribution of Bacillus thuringiensis Isolates», Appl. Environ. Microbiol. 55:2437- 2442, EE.UU., 1989.
Medrano, O. M.; O. H. A. Luna; J. J, Peña; J. M. Sánchez: «Supervivencia de células vegetativas de Bacillus thuringiensis en la espermosfera-rizosfera de frijol», Terra 18:333-337, 2000. (falta país)
Porcar, M.; V. Juárez: «Aislamiento y establecimiento de una colección de Bacillus thuringiensis», Bacillus thuringiensis en el control biológico, Universidad Nacional de Colombia, Bogotá, 2004, pp. 69-100.
Quesada E.; E. García; P. Valverde; C. Santiago: «Solation, Geographical Diversity and Insecticidal Activity of Bacillus thuringiensis from Soils in Spain», Microbiological Research 159:59-71, 2004. (falta país)
Rosas, N. M.; M. Mireles; J. L. Hernández; J. E. Ibarra: «Screening of Cry Gene Contents of Bacillus thuringiensis Strains Isolated from Avocado Orchards in Mexico, and Their Insecticidal Activity Towards Argyrotaenia sp. (Lepidoptera: Tortricidae) Larvae», J. Appl. Microbiol. 104:224-230, EE.UU., 2008.
Ruiz de Escudero, I.; M. A. Padilla; A. A. Ibáñez; P. Caballero: «Aislamiento y caracterización de nuevas cepas de Bacillus thuringiensis procedentes de muestras de tierra de Canarias», Bol. San. Veg. Plagas, 30:703-712, España, 2004.
Sánchez, J. M.; J. J. Peña: «Supervivencia de esporas de Bacillus thuringiensis en el filoplano de maíz, frijol y en suelo», Terra 18:325- 331, Brasil, 2000.
Támez, P.; M. M. Iracheta; B. Pereira; L. J. Galán; R. Gómez; R. Támez; C. Rodríguez: «Caracterización de cepas mexicanas de Bacillus thuringiensis tóxicas para larvas de lepidópteros y coleópteros», Ciencia UANL 8 (4):477-482, México, 2007.
Valenzuela, E.: «Guía Pasos prácticos. Microbiología» 112, Instituto de Microbiología, Universidad Austral de Chile, 2003.

