










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkFitosanidad
versión On-line ISSN 1818-1686
Fitosanidad v.14 n.2 Ciudad de la Habana abr.-jun. 2010
Control biológico
Evaluación microbiológica in vitro de mezclas de especies de hongos entomopatógenos ingredientes activos de bioplaguicidas cubanos
In vitro Microbiological Evaluation of Entomopathogenic Fungi Mixtures Used as Active Ingredients in Cuban Biopesticides
Orestes Elósegui Claro y Ana Ibis Elizondo Silva
Instituto de Investigaciones de Sanidad Vegetal. Calle 110 no. 514 e/ 5.a B y 5.a F, Playa, Ciudad de La Habana, C. P. 11600
RESUMEN
Se ha demostrado in vitro que la mezcla de cepas puede traer efectos beneficiosos o perjudiciales en el biocontrol de plagas agrícolas. En este trabajo se estudió el efecto in vitro sobre la germinación de mezclas duales de esporas en cocultivo de Beauveria bassiana, Metarhizium anisopliae y Lecanicillium lecanii, así como la interacción de estos hongos en cultivos duales. Las cepas usadas son ingrediente activo de bioplaguicidas comerciales cubanos. Las mezclas no afectaron la germinación de los conidios, donde se obtuvo un porcentaje de germinación superior al 95% para cada especie en cocultivo. En los cultivos duales la tasa de crecimiento radial de la colonia no se afectó en los primeros días; pero a partir del 13 al 15 comenzó una disminución, con un cese del crecimiento en la tercera semana y una distancia entre las márgenes de cada colonia mayor de 4-6 mm. Este comportamiento permitió clasificar el tipo de interacción como inhibición mutua a distancia. Estos resultados permiten iniciar trabajos para evaluar in vivo las cepas de estos hongos entomopatógenos en mezcla a través de bioensayos de laboratorio en una primera etapa, con el propósito de conocer si es factible su uso práctico sobre el control de determinadas plagas diarias.
Palabras claves: control biológico, Metarhizium anisopliae, Beauveria bassiana, Lecanicillium lecanii, bioplaguicidas
ABSTRACT
It has been demonstrated that in vitro strain mixtures may have beneficial or detrimental effects on crop pest biocontrol. In this paper in vitro effect of Beauveria bassiana, Metarhizium anisopliae and Lecanicillium lecanii spores dual mixtures on conidia germination in co-culture was assessed as well as interaction between these fungi in dual cultures. The fungal strains used were active ingredients of commercial Cuban biopesticides. Conidial mixtures did not change the conidial percentage germination with a percentage higher than 95% for every species in co-culture. In dual cultures colony radial growth rate in the first days did not change but from day 13 to day 15 a decrease in growth rate was visible. The colony growth also stopped by the third week when the margins of both colonies were about 4-6 mm away from each other. This behaviour allowed consider the type of interaction between both fungi as a mutual inhibition at distance. These results are basic for starting research about in vivo assessment of in-mixture entomopathogenic fungi through bioassays in a first stage, in order to know their feasible practical use on the control of certain target pests.
Keywords: biological control, Metarhizium anisopliae, Beauveria bassiana, Lecanicillium lecanii, biopesticides
INTRODUCCIÓN
El control biológico constituye una alternativa de gran importancia en los programas de manejo integrado de plagas (MIP). Los hongos entomopatógenos pueden estar en armonía con las diferentes técnicas usadas en esquemas de manejo integrado [Butt et al., 2001; Lacey et al., 2001].
En los últimos años se ha reforzado la posibilidad de mezcla de cepas o especies de hongos biocontroladores para mejorar su actuación sobre la plaga. Tigano-Milani et al. (1995) propusieron la hipótesis de que más de un haplotipo (clon) se debía requerir para iniciar y mantener una epizootia _en referencia a los hongos entomopatógenos_ en una población de insectos heterogénea y natural; sin embargo, la mezcla in vitro de más de una cepa puede traer efectos beneficiosos o perjudiciales en el biocontrol de plagas agrícolas. En Colombia, Cruz et al. (2006) demostraron que el uso combinado de esporas de cepas de Beauveria bassiana podía tener un efecto positivo o negativo sobre la virulencia de Hypothenemus hampei Ferr. (broca del café), lo que apoyó el planteamiento de que la variabilidad de las cepas de este hongo para el control de la broca del café era el factor que pudiera permitir al hongo adaptarse a las condiciones ambientales cambiantes, y atacar exitosamente diferentes poblaciones de insectos.
Inglis et al. (1997) ya habían demostrado que la eficacia como controlador de Beauveria bassiana hacia plagas de acrídidos era superior a la de Metarhizium flavoviride bajo condiciones de temperaturas frescas, mientras que M. flavoviride se favorecía bajo condiciones simuladas de calor. Cuando ambos patógenos se aplicaron simultáneamente, la mortalidad era mayor para la combinación B. bassiana y M. flavoviride que para M. flavoviride solo en ambiente cálido simulado, e igual a B. bassiana sola en un ambiente fresco simulado. Estos resultados sugirieron que las mezclas de aislados fúngicos pudieran usarse para aumentar el rango de tolerancia a la temperatura, comparado con el que las especies fúngicas, individualmente, pudieran tolerar para provocar la enfermedad en acrídidos.
En Cuba el Centro Nacional de Sanidad Vegetal (CNSV) recomienda la combinación de la lucha biológica con la química para el control de plagas. Para los biológicos se aconsejaba ya en el 2000 el uso de biopreparados microbianos a base de Bacillus thuringiensis en mezclas con B. bassiana, M. anisopliae y Verticillium lecanii (syn. Lecanicillium sp.), basado en resultados en evaluaciones de campo [CNSV, 2000]. En experimentos de campo realizados durante el 2001 y el 2002, en el cultivo de la papa en La Habana se utilizaron mezclas de hongos entomopatógenos para el control de Thrips palmi Karny con resultados alentadores [Elizondoet al., 2008], por lo que un estudio más profundo de la aplicación de estos bioplaguicidas en mezclas pudiera contribuir a reducir la incidencia de plagas sin el requerimiento de otras tácticas de manejo incluidos agroquímicos.
En los últimos años se comercializan unos pocos formulados con mezcla de hongos, tanto con efecto entomopatógeno de mezcla de hasta tres hongos como B. bassiana, M. anisopliae y Verticillium lecanii (syn. Lecanicillium lecanii) [Nutri-Life Myco-Force, 2005] como formulaciones con mezclas duales de hongos entomopatógenos [Plantisana, 2003].
De acuerdo con estos antecedentes sobre la posibilidad de un mejor control de determinadas plagas, cuando se usa más de un hongo entomopatógeno en mezcla con otra cepa de la misma especie o de otra, surge la necesidad de estudiar el efecto de mezclas de cepas de hongos entomopatógenos autorizados en la producción en Cuba, desde el nivel básico de laboratorio hasta nuevos ensayos biológicos de campo. Para aportar datos al estudio básico de laboratorio se realizó este trabajo, donde se plantearon como objetivos evaluar el efecto in vitro sobre la germinación de mezclas conidiales duales de conidios de Beauveria bassiana, Metarhizium anisopliae y Lecanicillium lecanii, y evaluar el efecto in vitro de cultivos duales B. bassiana, M. anisopliae y L. lecanii sobre el crecimiento fúngico.
MATERIALES Y MÉTODOS
Para la determinación del efecto in vitro de mezclas de suspensiones conidiales de B. bassiana, M. anisopliae y L. lecanii sobre la germinación de los conidios, se prepararon mezclas conidiales a partes iguales de cultivos recién esporulados de B. bassiana cepa Bb-1, M. anisopliae cepa Ma-11 y L. lecanii (syn. V. lecanii) cepa Vl-5 a una concentración de 1 x 107 conidios • mL_1, determinada en cámara de conteo de Neubauer. Se prepararon cámaras húmedas que contenían portaobjetos con una fina capa de agar papa dextrosa (PDA), y se sembró 0,1 mL de una suspensión conidial que se ajustó a 1 x 106 conidios • mL_1. Las mezclas fueron incubadas por 20 h a 27°C. Se evaluó a las 24 h el porcentaje de conidios germinados de cada especie en cultivo dual bajo el microscopio óptico con contraste de fase a 800x. Se contó como germinado la espora que emitió el tubo germinativo independientemente de su longitud. Los conidios de cada especie del hongo ensayado en la mezcla (cocultivo) se sembraron individualmente, bajo iguales condiciones, los que se tomaron como controles negativos. El experimento fue replicado cinco veces y se calculó el valor medio para cada caso.
El efecto de cultivos duales de B. bassiana, M. anisopliae y L. lecanii sobre el crecimiento de la colonia se estudió por medio de la preparación de una suspensión conidial a una concentración de 106 conidios • mL_1 de B. bassiana cepa Bb-1, M. anisopliae cepa Ma-11 y L. lecanii (syn. V. lecanii) cepa Vl-5, de forma independiente, y se sembró 1 mL de cada suspensión de cada especie con espátula de Drigalski en placas Petri con PDA. Estas se incubaron a 27°C por cuatro días solamente para garantizar cultivos jóvenes sin esporular. Transcurrido este tiempo se tomaron discos de 4 mm de diámetro de los cultivos de la periferia de cada colonia de cada especie, los que se colocaron en los extremos opuestos de placas Petri de 9 cm de diámetro con PDA, a 7 cm de distancia un microorganismo del otro, de forma que se lograran cultivos duales de las tres especies fúngicas. Las placas fueron incubadas a 27°C en la oscuridad, y se midieron dos diámetros perpendiculares para la colonia de cada especie en cultivo cada 60 h a partir del día cinco hasta que el crecimiento cesara. Se tomaron cultivos únicos (puros) de cada especie como controles. Los tratamientos se replicaron cinco veces. Se halló el valor medio del radio de la colonia en milímetros. Se realizó un análisis de regresión lineal con la variable radio de la colonia (variable dependiente) contra el tiempo (variable independiente) para un valor de p < 0,01. El programa usado fue Statgraphics Plus versión 5.0. El tipo de interacción de los microorganismos se estableció según la escala propuesta por Magan y Lacey (1984) (Tabla 1).

En todos los experimentos los cultivos de partida de los hongos para los ensayos se obtuvieron en cuñas de agar dextrosa de Sabouraud, a un tiempo menor de ocho días de edad después de lograda la esporulación masiva del hongo, con parámetros de calidad adecuados (germinación conidial 90%, 108 conidios • mL_1 (NC 72-03, 1993; NC 72-04, 1993; NC 72-05, 1993), y las cepas correspondieron a las autorizadas en Cuba para su uso en la producción masiva de estos hongos.
RESULTADOS Y DISCUSIÓN
El comportamiento de la germinación en las mezclas de conidios de los hongos entomopatógenos ensayados arrojó que el porcentaje de conidios germinados era muy similar con respecto a los controles (Tabla 2). El desarrollo de los tubos germinativos fue el típico para cada especie estudiada.
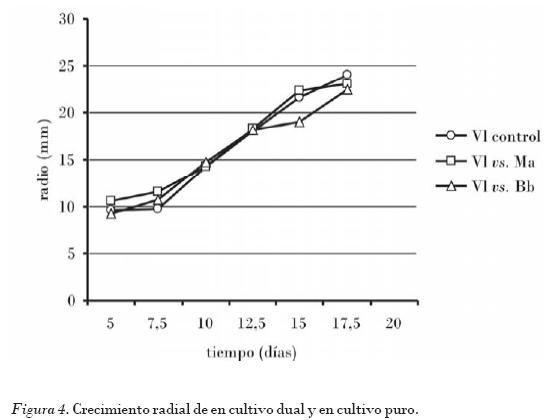
Los experimentos in vitro de cultivos duales de los hongos entomopatógenos B. bassiana, M. anisopliae y L. lecanii sobre el crecimiento mostraron resultados muy similares hasta el 14 con respecto al crecimiento en los controles. A partir de este momento se observó una tendencia a la disminución del crecimiento, lo que correspondió a una cercanía física de las márgenes de las colonias de ambas especies, donde la expresión a través del crecimiento de una posible interacción se hicieron mayores. Del 17 en adelante no hubo crecimiento adicional de ninguna de las colonias de las especies en cultivo en sentido de uno de los ejes perpendiculares al frente de crecimiento. Cuando las colonias se observaron al estereoscopio se vio una delimitación física entre las márgenes de cada cultivo, y no se pudieron detectar fenómenos de invasión del espacio colonizado por la especie opuesta (Figs. 1, 2, 3 y 4).

Ecuaciones de las rectas obtenidas para los experimentos con Metarhizium:
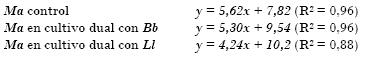
Ecuaciones de las rectas obtenidas para los experimentos con Beauveria:
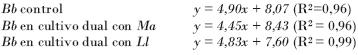
Ecuaciones de las rectas obtenidas para los experimentos con Lecanicillium:
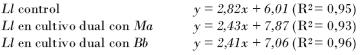
Nótese cómo los valores de las pendientes en las ecuaciones de las rectas de los cultivos controles son mayores que las pendientes de las rectas obtenidas en los cultivos duales, lo que indica que hubo una disminución de la velocidad de crecimiento en los cultivos duales (inhibición del crecimiento).
No es posible en este tipo de interacción hablar de una especie competidora, por lo que de acuerdo con la escala propuesta por Magan y Lacey (1984), la interacción presente es del tipo inhibición mutua del crecimiento.
En el caso de los cultivos duales la zona de inhibición entre las márgenes de cada colonia fue aproximadamente de 4-6 mm, por lo que se puede clasificar como inhibición mutua a distancia (Figs. 1, 2, 3 y 4).
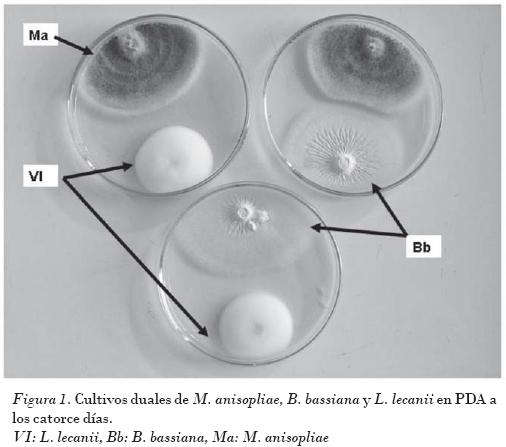
La temperatura y la actividad del agua puede afectar el tipo de interacción que se presente entre las especies fúngicas en cultivos duales, como lo demuestran los ensayos con hongos fitopatógenos como Alternaria alternata, Cladosporium cladosporioides y C. herbarum, y la evaluación de la capacidad competidora de V. lecanii [Magan y Lacey, 1984] o con hongos productores de micotoxinas como Fusarium sambucinum y Phoma glomerata [Caselles y Santamarina, 2007]. En el caso presentado estas variables se fijaron a valores óptimos en el cultivo.

La zona de no crecimiento entre las márgenes de las colonias en el cultivo dual se observó bajo el microscopio estereoscopio, y no se apreció proyección de las hifas de ninguna de las especies en el cultivo en la zona del halo que pudieran contactar las márgenes de la otra especie, lo que sugiere que el efecto sobre la inhibición del crecimiento es indirecto a través de la excreción de ciertos metabolitos por parte de las especies en cultivo.
El hecho de que la germinación no se afectara y el crecimiento durante los primeros días se mantuviera bastante estable ((Figs. 2, 3 y 4)), son datos relevantes que sugieren que las cepas de estos microorganismos pudieran estar en mezclas duales en formulados, si se tiene en cuenta que la germinación y la penetración a través de la cutícula del insecto conjuntamente son eventos claves tempranos para lograr la patogenicidad [Hughes et al., 2004]; sin embargo, se ha demostrado que una vez en el hemocele las coinfecciones de hongos entomopatógenos interaccionan de diversas formas. De hecho, el efecto real de mezclas de hongos entomopatógenos usados al unísono en el control de una misma especie plaga es muy controversial en la literatura consultada [Tigano-Milani, 1995; Inglis et al., 1997; Inglis et al., 1999; Hughes et al., 2004, Cruz et al., 2006].
Por otra parte, contar en los cultivos duales evaluados con una inhibición mutua a distancia que sugiera inhibición indirecta del crecimiento, pudiera tener un posible efecto indeseado in vivo una vez que las especies fúngicas participantes en la coinfección penetren al hemocele con la consiguiente producción de sustancias activas que puedan dar lugar a interacciones no deseadas para el éxito del proceso infeccioso, y que debe culminar con la muerte y colonización del insecto infectado.
Hay estudios que apuntan a esta hipótesis, donde se demostró que el uso combinado de esporas de cepas de Beauveria bassiana, comprobadas genéticamente que eran diferentes, podían tener un efecto antagónico o sinérgico contra la broca del café (H. hampei), comparado con el uso por separado de cada una de ellas. Las cepas sin diferencias detectables genéticamente no mostraron cambios en la virulencia al evaluarse mezcladas; sin embargo, las cepas individuales con una virulencia menor del 80% mostraron porcentajes de mortalidad del 93% cuando se usaron mezclas de ellas en los bioensayos contra broca. En contraste, cepas igualmente diferentes genéticamente, con virulencias individuales superiores al 85%, mostraron virulencias menores (57%) cuando se usaron en mezclas [Cruz et al., 2006].
Inglis et al. (1999) informaron de la influencia de la temperatura fluctuante sobre la infección competitiva y la colonización del saltamonte (Melanoplus sanguinipes F.) por B. bassiana y M. anisopilae var. acridae. Aunque la aplicación de los dos hongos resultó en coinfección, la mortalidad no se afectó significativamente. En algunos casos la población total de hongos de ambas especies en el hemocele de las ninfas coinoculadas fue menor que en las ninfas tratadas con uno de los hongos (una sola especie), lo que indicaba un grado de antagonismo interespecie. Estos autores informaron que entre el 28 y el 39% de los insectos inoculados con el o los hongos no podían ser aislados nuevamente de los cadáveres cuando el ensayo; sin embargo, seguía un comportamiento con menos del 10% de mortalidad en la muestra testigo. Las razones de esta conducta se desconocen. De esto resultó que el tomar la colonización por el hongo de los cadáveres de insectos como indicador de la causa de la mortalidad en este tipo de infecciones puede fallar.
Los mismos autores Inglis et al. (1999) detectaron que a una temperatura controlada constante de 25°C, Beauveria prevalecía sobre Metarhizium en el hemocele, y cuando el rango de temperatura en el bioensayo crecía, las poblaciones de Metarhizium aumentaban en relación con Beauveria. Ello sugería que la temperatura influenciaba su compatibilidad. Los autores concluyeron que la ventaja de combinar en este caso los dos hongos estaría reservada para condiciones climáticas caracterizada por días frescos (temperatura inferior a 25°C).
Sin embargo, existen formulaciones con mezcla de cepas de hongos biocontroladores, bien para mejorar la eficiencia en el control de una misma plaga o para el control de más de una [Nutri-Life Myco-Force, 2005; Plantisana, 2003]; a pesar de que estudios básicos que involucren las especies plagas que soporten la utilidad de estas formulaciones han sido controversiales. Por ello se considera que se necesitan estudios más profundos para esclarecer la forma de actuación de estos formulados y su eficiencia en el biocontrol de plagas, tanto in vitro como en condiciones de campo, así como su impacto en el agroecosistema.
CONCLUSIONES
• Las mezclas conidiales de Beauveria bassiana, Lecanicillium lecanii y Metarhizium anisopliae a las concentraciones de 106 conidios • mL_1 no afectaron la germinación de los conidios en cocultivo.
• En los cultivos duales de Beauveria bassiana, Lecanicillium lecanii y Metarhizium anisopliae se presentó una interacción del tipo inhibición mutua a distancia.
REFERENCIAS
Butt, T. M.; C. Jackson; N. Magan: «Introduction-Fungal Biocontrol Agents: Progress, Problems and Potential», Fungi as Biocontrol Agents, Chapter 1, CAB International, Inglaterra, 2001, pp.1-8.
Caselles, R.; M. P. Santamarina: «Interacciones competitivas entre Fusarium sambucinum Fuckel y Phoma glomerata (Corda) Wollenweber & Hochapfel en condiciones in vitro», Rev. Iberoam. Micol . 24:29-33, España, 2007.
CNSV: «Programa de defensa fitosanitaria para el cultivo de la papa», Minag, Ciudad de La Habana, 2000.
Cruz, L. P.; A. L. Gaitán; C. E. Góngora: «Exploiting the Genetic Diversity of Beauveria bassiana for Improving the Biological Control of the Coffee Berry Borer Through the Use of Strain Mixtures», Appl Microbiol. Biotechnol. 71(6):918-926, Alemania, 2006.
Elizondo, A. I.; C. A. Murguido; O. Elósegui; Y. Matienzo; E. Massó: «Nuevas tácticas para el manejo integrado de Thrips palmi Karny en el cultivo de la papa (MI-C11)», Resúmenes del II Taller Internacional de Manejo de Plagas, Fitosanidad 12(4):242, La Habana, 2008.
Hughes, W. O.; K. S. Petersen; L. V. Ugelvig; D. Pedersen; L. Thomsen; M. Poulsen; J. J. Boosma: «Density-Dependence and Within-Host Competition in a Semelparous Parasite of Leaf-Cutting Ants», BMC Evolutionary Biology 4:45, www.biomedcentral.com/1471-2148/4/45, 2004 (consulta: 23 de mayo del 2008).
Inglis, G. D.; D. L. Jonson; K. J. Cheng; M. S. Goettel: «Use of Pathogen Combinations to Overcome Constraints of Temperature on Entomopathogenic Hyphomycetes Against Grasshoppers», Biol. Contr. 8 (2):143-152, Elsevier, Holanda,1997.
Inglis, G. D.; G. M. Duke; L. M. Kawchuk; M. S. Goettel: «Influence of Oscillating Temperatures on the Competitive Infection and Colonization of the Migratory Grasshopper by Beauveria bassiana and Metarhizium anisopliae», Biol. Contr. 14 (2):111-120, Elsevier, Holanda. 1999.
Lacey, L.; D. R. Horton; T. R. Unruh; K. Pike; M. Márquez: «Control biológico de plagas de papas en Norteamérica», Taller en español sobre producción de papas, Proc. Wash. State Potato Conf. Trade Fair, Washington, 2001, pp. 103-117.
Magan, N.; J. Lacey: «Effect of Water Activity, Temperature and Substrate on Interactions Between Field and Storage Fungi», Trans. Br. Mycol. Soc. 82 (1):83-93, Inglaterra, 1984.
NC 72-03: «Biopreparado del entomopatógeno Verticillium lecanii. Especificaciones. Biotecnología Agrícola», norma cubana, 1993.
NC 72-04: «Biopreparado del entomopatógeno Metarhizium anisopliae. Especificaciones. Biotecnología Agrícola», norma cubana, 1993.
NC 72-05: «Biopreparado del entomopatógeno Beauveria bassiana. Especificaciones. Biotecnología Agrícola», norma cubana, 1993.
Nutri-Life Myco Force: «Lawrie & Co. Sustainable Agriculture Products», 2005, www.nrrbs.com.au/ntsmycoforce.htm (consulta: 20 de marzo del 2008).
Plantisana: «Productos Platisana», 2003, http://productos-plantisana. com/aboutus.aspx (consulta: 12 de enero del 2010).
Tigano-Milani, M. S.; R. J. Honeycutt; L. A. Lacey; R. Assis; M. McClelland; B. W. S. Sobral: «Genetic Variability of Paecilomyces fumosoroseus Isolates Revealed by Molecular Markers», J. Invertebr. Pathol. 65 (3):274-282, EE. UU., 1995.

